
温度对紫茎泽兰茎秆产沼气的影响
2015-08-20 17:24陈金发
江苏农业科学 2015年7期
陈金发

摘要:以2种不同预处理后的紫茎泽兰茎秆作为直接发酵原料,对厌氧发酵在不同温度条件下(25~50 ℃)产气情况以及对原料的利用情况进行比较,结果表明:(1)中温条件(25~35 ℃)是发酵的最佳温度,发酵产气量与产甲烷率随温度升高而上升;40~45 ℃不利于发酵微生物(包括非产甲烷微生物与产甲烷微生物)的发酵作用;50 ℃时,嗜热菌生长良好,产气量与产甲烷率得到恢复。(2)发酵甲烷直接利用对象主要是半纤维素,温度上升不利于发酵微生物对木质纤维的水解,所以粉状原料较杆状原料更适合发酵产甲烷。本试验条件下,粉状原料30 ℃时产气与产甲烷最佳,TS(总固体)与VS(有机干物质)产气率与产甲烷效率分别为152.8 L/kg TS、74.3 L/kg VS。
关键词:紫茎泽兰;甲烷;温度;原料利用;发酵
中图分类号: S216.4 文献标志码: A 文章编号:1002-1302(2015)07-0400-04
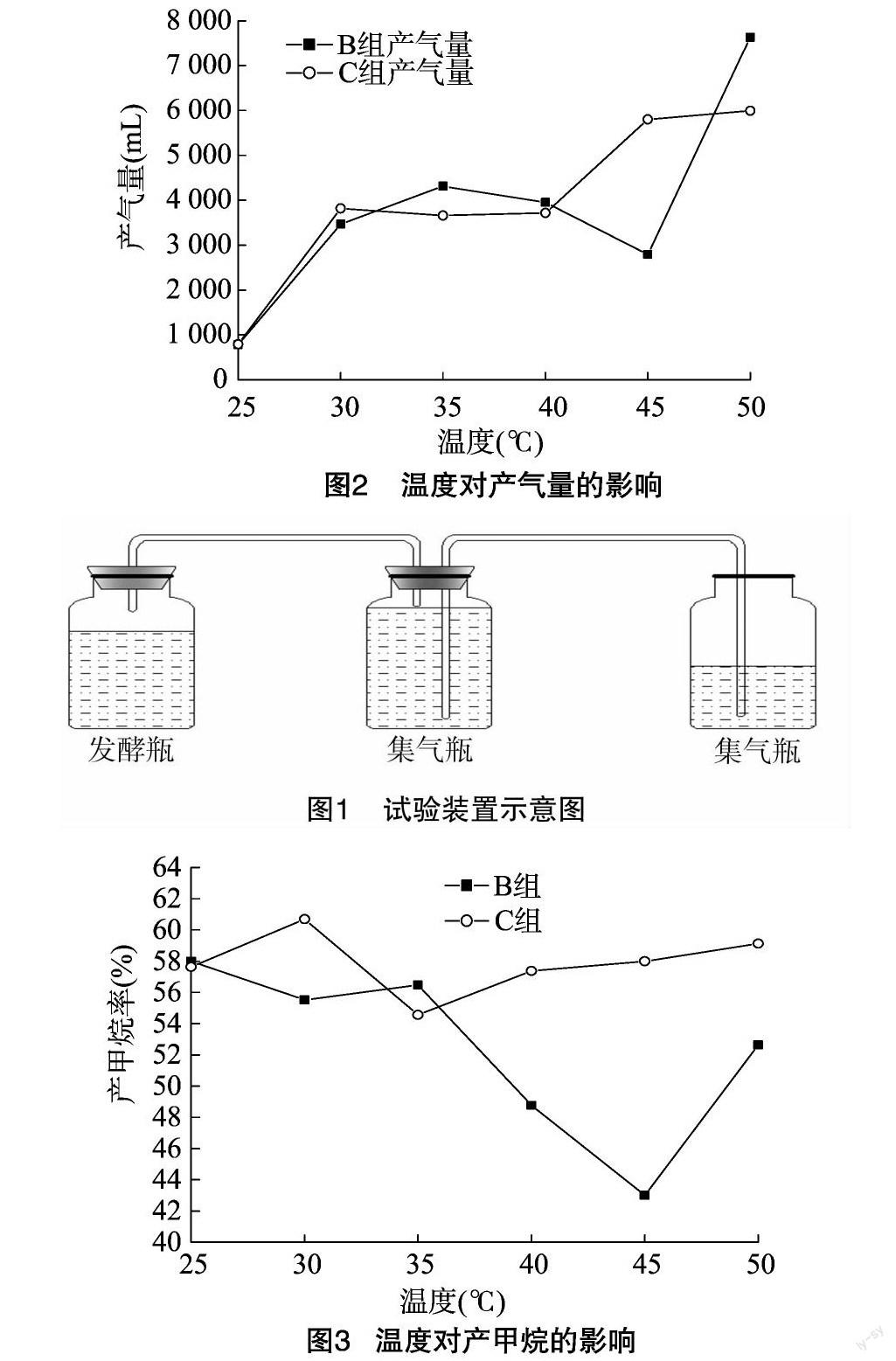
通过对日产气情况的分析可以看出,在预处理过程中,发酵微生物尤其是非产甲烷微生物有效富集,使得发酵前期具有较好的产气能力,且随着温度的上升,初期的产气能力呈增长趋势,产甲烷率却呈相反的趋势。非产甲烷微生物与产甲烷微生物的作用温度与作用时间存在明显差异,非产甲烷微生物对高温适应能力强,所以产甲烷微生物的富集较非产甲烷微生物而言要晚10 d左右,这10 d时间内处于发酵水解阶段,主要是非产甲烷微生物对纤维素成分进行水解,为产甲烷微生物提供可直接利用的原料[24],当非产甲烷微生物对原料进行有效水解后产甲烷微生物生长稳定并已经较好地适应发酵环境,产甲烷率随着发酵时间增加而逐渐增大且在后期达到70%以上。
B组原料在45 ℃下日产气与其他温度下相近,但产甲烷效率较低,然而C组却有较高的甲烷产率。可见,在这一温度下物料状态对于原料能否被有效利用影响较大,木质素的存在是纤维素类原料沼气发酵中限速因素,厌氧条件下木质素很难生物降解[25]。这可能是由于B组原料未被粉碎,未被粉碎的原料木质结构较粉状更为严密,导致微生物不能进行有效分解利用。
2.2 不同温度下发酵对TS、VS的利用情况
2.2.1 温度对TS产气影响 TS(总固体)利用情况是发酵微生物对原料利用产气效率的重要评价指标。由图4可见,中温条件下B、C组TS利用率随温度升高而上升,40~45 ℃条件下TS利用率急剧下降,B组材料在40 ℃条件下最低,只有2.5%,50 ℃恢复到13%左右。可见,40~45 ℃不利于发酵微生物群对原料的利用。
C组TS产气率总体明显高于B组,粉状原料较杆状原料更易被直接利用。C组在30 ℃的TS产气率最高,达到峰值152.8 L/kg。
2.2.2 温度对VS产甲烷影响 VS(有机干物质)产甲烷情况是产甲烷微生物对有机物质(即含有机碳部分)转化利用的评价标准之一。由图5可以看出,C组VS的利用受到温度的影响,中温条件下VS利用率与温度成正比。B组在45 ℃下利用率最低,表明该温度对杆状原料的影响比对粉状影响大。
C组VS产甲烷率明显高于B组,表明粉状原料较杆状原料更易被直接利用。C组在30 ℃下,VS产气率达到 74.3 L/kg 的峰值。
2.3 原料不同预处理对利用效率的影响
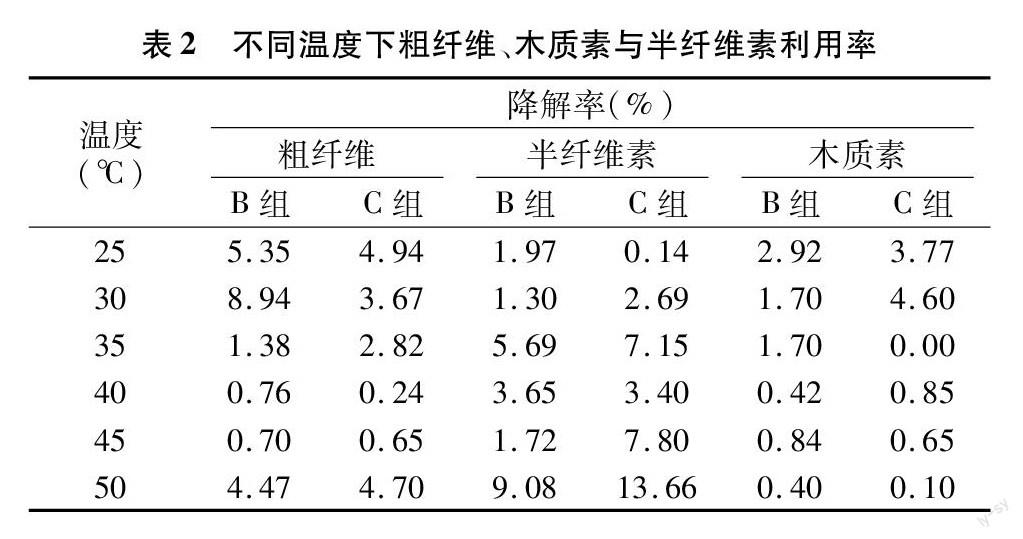
粗纤维是植物细胞壁的主要组成成分,包括纤维素、半纤维素、木质素及角质等。通过半纤维素与木质素的降解数据(表2)可以看出,B、C 2组原料中半纤维素平均利用率分别为4.7%、7.0%,C组原料的半纤维素的利用率约为B组的1.5倍,原料利用率更高。2组原料的木质素的平均利用率都为1.6%,可以看出发酵过程中发酵微生物对木质素没有进行有效的利用,相比半纤维素,天然木质纤维素结构更加复杂。进行生物转化的难点在于如何将木质素与纤维素降解成为可发酵糖[25]。生产甲烷的原料利用则主要集中在半纤维素上,未经粉碎的杆状原料相比于粉状原料,木质纤维结构更加紧密,从而降低了微生物对其的利用效果。杆状原料的木质素比粉状原料更难以被水解,进一步影响了微生物利用半纤维等成分进行沼气发酵。
3 结论
试验表明,中温条件(25~35 ℃)是发酵的最佳温度,发酵产气量与产甲烷率随温度升高而上升。该湿度条件下,有利于嗜中温菌发酵微生物群的富集作用,且对原料(TS、VS)的利用也随温度升高而提高。40~45 ℃是发酵的不利温度,是嗜中温菌与嗜热菌的过渡温度,不利于发酵微生物(包括非产甲烷微生物与产甲烷微生物)的发酵作用,发酵产气量、产甲烷率以及对原料的利用都受到一定限制,当达到50 ℃时,嗜热菌生长良好,成为发酵作用主要微生物群,产气量与产甲烷率得到恢复。
粉状原料較杆状原料更适于发酵产甲烷,且受温度影响程度明显低于杆状原料。本试验条件下,粉状30 ℃时产气情况最佳,原料TS产气率达到152.8 L/kg,VS产甲烷率达到74.3 L/kg。
参考文献:
[1]段 惠,强 盛,吴海荣. 紫茎泽兰(Eupatorium adenophorum Spreng)[J]. 杂草科学,2003(2): 36-38.
[2]华劲松. 凉山州紫茎泽兰的分布及危害情况调查[J]. 现代农业科技,2012(22):139-140.
[3]李 丽,张无敌,尹 芳. 紫茎泽兰的各种利用研究[J]. 农业与技术,2007,27(4):51-54.
[4]王银朝,赵宝玉,樊泽锋,等. 紫茎泽兰及其危害研究进展[J]. 动物医学进展,2005,26(5):45-48.
[5]杨紫美. 紫茎泽兰的危害及防除[J]. 草业与畜牧,2011(1):32,34.
[6]梁小玉,张新全. 紫茎泽兰发生特点、防治及其利用[J]. 四川草原,2004(2):13-15.
[7]赵运涛,李新华,张征文. 中国主要外来入侵物种的危害及防治措施[J]. 林业调查规划,2005,30(6):113-117.
[8]张无敌. 恶性有毒杂草紫茎泽兰的利用[J]. 云南林业科技,1996,74(1):78-81.
[9]朱先洲. 入侵植物紫茎泽兰资源化利用研究进展[J]. 湖北农业科学,2012,51(14):2905-2907,2917.
[10]雷永松,陈 亮,赵 青. 外来恶性杂草紫茎泽兰综合利用研究进展[J]. 湖北林业科技,2007(3):36-39.
[11]侯太平,刘世贵. 有毒植物紫茎泽兰研究进展[J]. 国外畜牧学:草原与牧草,1999(4):6-8.
[12]杨发根,段家锦,朱桂玲. 紫茎泽兰脱毒作猪饲料原料的研究[J]. 粮食与饲料工业,1998(5):21-22.
[13]江蕴华,余晓华. 用紫茎泽兰生产沼气的研究[J]. 太阳能学报,1986,7(3):288-294.
[14]Sang W G,Zhu L,Axmacher J C. Invasion pattern of Eupatorium adenophorum Spreng in southern China[J]. Biological Invasions,2010,12(6): 1721-1730.
[15]闫乾胜,杨 婕,李华民,等. 入侵物种紫茎泽兰化学成分及生物活性研究进展[J]. 北京师范大学学报:自然科学版,2006,42(1):70-73.
[16]白 洁,曹坳程,郭美霞,等. 紫茎泽兰9-羰基-10,11-去氢泽兰酮分布积累动态[J]. 植物学报,2011,46(4):470-475.
[17]陳金发,廖茂芪,周家兴,等. 紫茎泽兰茎秆厌氧发酵产甲烷[J]. 环境工程,2014,32(4):153-157.
[18]裴占江,王大慰,张 楠,等. 温度对产甲烷菌群发酵性能的影响[J]. 黑龙江农业科学,2009(5):128-129,140.
[19]王林风,程远超. 硝酸乙醇法测定纤维素含量[J]. 化学研究,2011,22(4):52-55,71.
[20]李 华,孔新刚,王 俊. 秸秆饲料中纤维素、半纤维素和木质素的定量分析研究[J]. 新疆农业大学学报,2007,30(3):65-68.
[21]张丽英. 饲料分析及饲料质量检测技术[M]. 北京:中国农业大学出版社,2003:68-70.
[22]于晓章,彭晓英,周朴华. 温度对厌氧嗜热菌群产甲烷能力的影响[J]. 湖南农业大学学报:自然科学版,2005,31(4):422-426.
[23]虞方伯,罗锡平,管莉菠,等. 沼气发酵微生物研究进展[J]. 安徽农业科学,2008,36(35):15658-15660.
[24]蒙 杰,王敦球. 沼气发酵微生物菌群的研究现状[J]. 广西农学报,2007,22(4):46-49.
[25]胡启春. 微生物降解木质纤维素研究的某些进展[J]. 中国沼气,1987,5(1):4-7.
猜你喜欢
疯狂英语·新读写(2021年8期)2021-11-05
军民两用技术与产品(2021年10期)2021-03-16
水上消防(2020年1期)2020-07-24
疯狂英语·新读写(2018年3期)2018-11-29
小学生优秀作文(高年级)(2018年4期)2018-09-11
少儿科学周刊·儿童版(2016年1期)2016-03-14
自动化与仪表(2014年10期)2014-02-26
