
2种干扰强度下清香木天然林群落特征及物种多样性
2015-08-20 11:25李晓芳王加国安明态袁丛军严令斌杨瑞
江苏农业科学 2015年7期
关键词:物种多样性
李晓芳 王加国 安明态 袁丛军 严令斌 杨瑞
摘要:采用常规群落学调查方法,设置12个典型群落样地,统计群落的Gleason物种丰富度指数、Simpson多样性指数、Shannon-Wiener多样性指数及Pielou均匀度指数值,研究2种干扰强度下清香木(Pistacia weinmannifolia)天然林群落结构及物种多样性。结果表明:除乔木层各多样性指数差异不明显外,都格弱干扰群落灌木层和草本层其余各多样性指数均为发耳强干扰群落的1倍或以上;2个群落均以中高位芽植物占优势,反映研究区域较为温热多湿的气候特征;发耳清香木群落物种频度等级分布规律为A>B>C=E,都格为A>B=C=D,2个群落频度组成均与 Raunkiaer 频度定律不吻合;人畜干扰是导致2个群落物种多样性差异及发耳灌木层、草本层不发达的主要原因。
关键词:清香木;干扰强度;群落特征;物种多样性
中图分类号: S718.5 文献标志码: A 文章编号:1002-1302(2015)07-0182-03
清香木(Pistacia weinmannifolia)隶属漆树科黄连木属常绿灌木至乔木,当地称之为“乌木”,野生于中国云南省、四川省、贵州省等地,是贵州省省级重点保护树种,主要分布在六盘水市、遵义市、黔南布依族苗族自治州及安顺市等地海拔500~1 350m的山地阔叶林及灌木林中。清香木树形美观,春夏翠绿,秋冬转红,叶片层次感好,果呈红色,根系发达,耐干旱,萌蘖能力强且寿命长,虫害率低,易造型,适合作为喀斯特地区尤其是干热河谷地带的造林先锋树种。清香木叶揉碎气味清香,皮、叶可提取芳香油,民间用作香料;枝叶及树皮可消炎解毒、收敛止泻;茎干可制作生活用品等,具有广阔的开发前景。物种多样性是人类赖以生存的生物资源,自然界中尚有很多具有潜在利用价值的生物资源未被开发利用[1]。研究植物群落物种多样性的目的是为了更好地保护利用多样性资源[2]。由于人们对野生资源进行乱砍滥伐、开荒及火灾等因素的干扰,清香木分布面积和数量均明显减少,物种多样性也随之减少。目前,我国有关清香木的研究多集中在种群生态[3-4]、苗木组培繁育与荒山造林[5-10]、耐旱性[11-14]及化学成分[15-19]等方面,对不同干扰强度下清香木群落及物种多样性特征方面的研究尚未见报道。笔者对不同干扰强度下清香木天然林群落特征及物种多样性进行研究,旨在为探讨清香木群落的自然规律及其种群拯救保育提供依据。
1 材料与方法
1.1 研究区概况
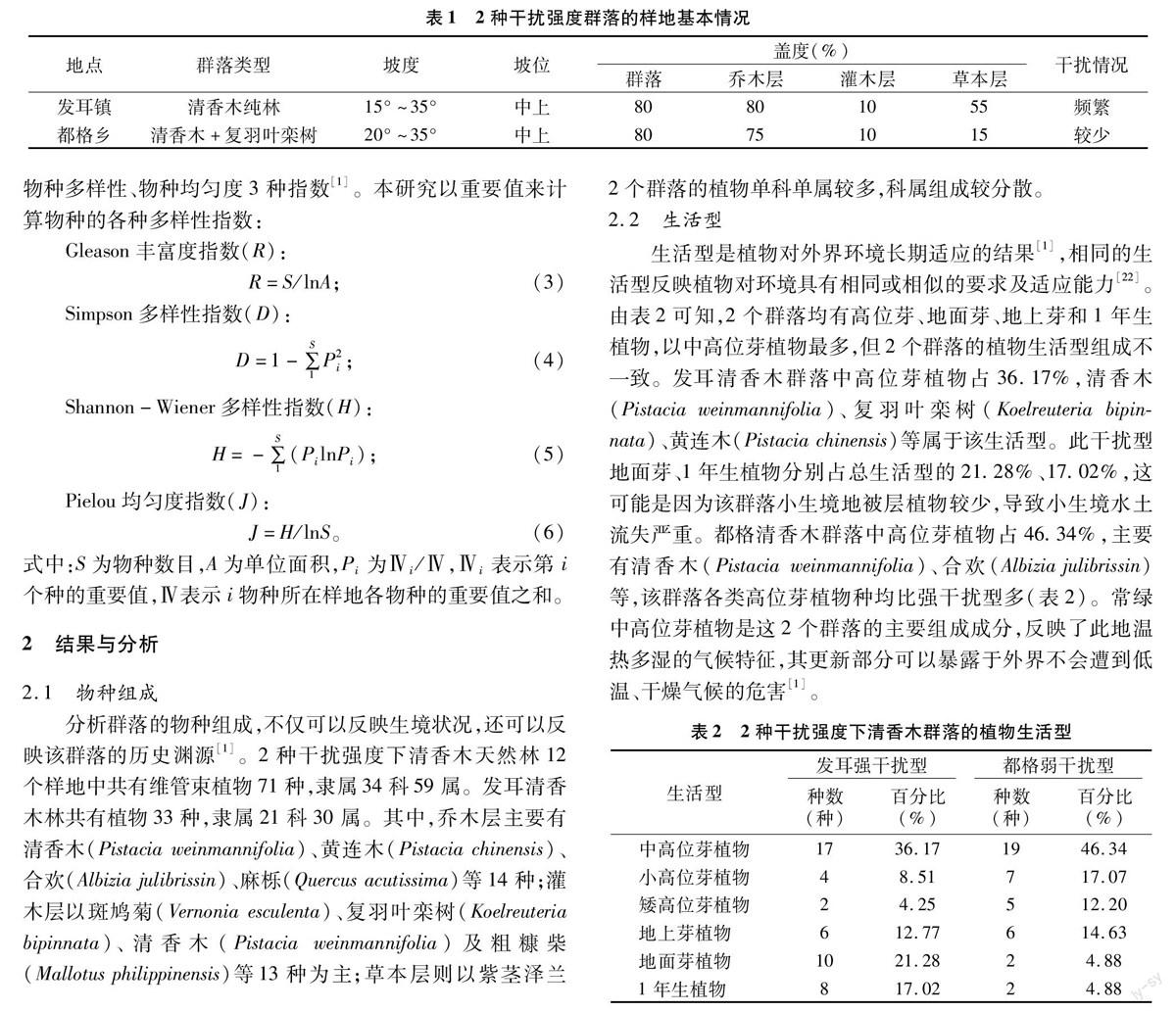
研究区位于贵州省水城县都格乡、发耳镇2个毗邻乡镇境内,是目前已知的清香木资源最丰富的2个天然乔木群落,也是目前仅存的2片天然分布的乔木群落,当地居民将其視为神树(风水林)。研究区属亚热带湿润季风气候,冬无严寒,夏无酷暑,年平均降水量1 100 mm,无霜期 250 d 左右。研究区位于山坡中部及中上部,海拔1 000~1 300 m,以紫色砂页岩、紫色土为主,主要伴生树种为复羽叶栾树(Koelreuteria bipinnata)、余甘子(Phyllanthus emblica)、栓皮栎(Quercus variabilis)、黄连木(Pistacia chinensis)、合欢(Albizia julibrissin)等常绿阔叶和次生性常绿落叶阔叶混交林,局部地段群落能形成清香木纯林。都格乡清香木林位于村寨后山,位置特殊,地段偏僻,人畜干扰相对较弱,属弱干扰类型。发耳镇清香木林临近村寨,尽管也是风水林,但林下人为、家禽家畜等活动较为频繁,属强干扰类型(表1)。
1.2 试验方法
2014年6月在研究区选择有代表性的地段进行典型样地法群落学调查,共设置12个20 m×30 m样地,总样地面积7 200 m2。采用相邻格子法进行观测,即在6个样地内布设72个10 m×10 m的乔木层样方;在每个样地的上、中、下分别等距布设5个2 m×2 m的灌木样方及5个1 m×1 m的草本样方进行灌木层、草本层调查,灌木层、草本层样方均为120个。记录海拔、坡向、坡度、郁闭度、基岩、土壤类型等指标,其中,乔木层记录各物种株数、胸径(地径)、树高、冠幅、起源、结实与否、健康状况等指标;灌木层各物种记录株数、地径、高度、盖度等指标;草本层各物种记录种名、株(丛)数、高度、盖度等指标。
1.3 数据处理
1.3.1 重要值计算 参照张金屯的方法[20]计算重要值,即:
乔木、灌木重要值(Ⅳ)=相对密度+相对频度+相对显著度;(1)
草本层重要值(Ⅳ)=相对频度+相对高度+相对盖度。(2)
1.3.2 物种多样性分析
由于重要值考虑了频度、盖度等多个参数,许多学者建议采用重要值来测度多样性指数[21]。α多样性用来测度均质群落内物种组成状况,包括物种丰富度、物种多样性、物种均匀度3种指数[1]。本研究以重要值来计算物种的各种多样性指数:
Gleason丰富度指数(R):
R=S/lnA;(3)
Simpson多样性指数(D):
D=1-∑S1P2i;(4)
Shannon-Wiener多样性指数(H):
H=-∑S1(PilnPi);(5)
Pielou均匀度指数(J):
J=H/lnS。(6)
式中:S为物种数目,A为单位面积,Pi为Ⅳi/Ⅳ,Ⅳi表示第i个种的重要值,Ⅳ表示i物种所在样地各物种的重要值之和。
2 结果与分析
2.1 物种组成
分析群落的物种组成,不仅可以反映生境状况,还可以反映该群落的历史渊源[1]。2种干扰强度下清香木天然林12个样地中共有维管束植物71种,隶属34科59属。发耳清香木林共有植物33种,隶属21科30属。其中,乔木层主要有清香木(Pistacia weinmannifolia)、黄连木(Pistacia chinensis)、合欢(Albizia julibrissin)、麻栎(Quercus acutissima)等14种;灌木层以斑鸠菊(Vernonia esculenta)、复羽叶栾树(Koelreuteria bipinnata)、清香木(Pistacia weinmannifolia)及粗糠柴(Mallotus philippinensis)等13种为主;草本层则以紫茎泽兰(Ageratina adenophora)、求米草(Oplismenus undulatifolius)、牛膝(Achyranthes bidentata)等10种为主。发耳群落物种多样性不高,这可能与其临近村寨,当地居民活动频繁(林间小道)及风俗习惯(如在大树下祭祀)等因素有关。都格清香木林共有维管束植物55种,录属28科47属。其中,乔木层主要有清香木(Pistacia weinmannifolia)、紫弹树(Celtis biondii)、复羽叶栾树(Koelreuteria bipinnata)等19种植物;灌木层物种多样性最高,有清香木(Pistacia weinmannifolia)、灰毛浆果楝(Cipadessa cinerascens)、斑鸠菊(Vernonia esculenta)等26种植物;草本层也相对较多,共有竹叶茅(Microstegium nudum)、紫茎泽兰(Ageratina adenophora)、千里光(Senecio scandens)、铁线蕨(Adiantum capillus-veneris)等24种植物。都格群落物种多样性较发耳高,这可能与其所处地段相对偏僻、人畜干扰较少等因素有关。从科属组成来看,发耳群落中只含有1种植物有12科,占总科数的57.14%;含5种或以上植物仅有1科,即菊科,仅占总科数的4.76%;含1种植物有24属,占总属数的80%。都格群落中含1种植物的有13科,占总科数的46.42%;含5种或以上植物的有6科,占总科数的21.43%;含1种植物的有30属,占总属数的6383%。可见,2个群落的植物单科单属较多,科属组成较分散。
2.2 生活型
生活型是植物对外界环境长期适应的结果[1],相同的生活型反映植物对环境具有相同或相似的要求及适应能力[22]。由表2可知,2个群落均有高位芽、地面芽、地上芽和1年生植物,以中高位芽植物最多,但2个群落的植物生活型组成不一致。发耳清香木群落中高位芽植物占36.17%,清香木(Pistacia weinmannifolia)、复羽叶栾树(Koelreuteria bipinnata)、黄连木(Pistacia chinensis)等属于该生活型。此干扰型地面芽、1年生植物分别占总生活型的21.28%、17.02%,这可能是因为该群落小生境地被层植物较少,导致小生境水土流失严重。都格清香木群落中高位芽植物占46.34%,主要有清香木(Pistacia weinmannifolia)、合欢(Albizia julibrissin)等,该群落各类高位芽植物种均比强干扰型多(表2)。常绿中高位芽植物是这2个群落的主要组成成分,反映了此地温热多湿的气候特征,其更新部分可以暴露于外界不會遭到低温、干燥气候的危害[1]。
2.3 频度分析
参照Raunkiaer的频度定律[1],将植物频度分为5个等级,即A级:1%~20%;B级:21%~40%;C级:41%~60%;D级:61%~80%;E级:81%~100%。频度越大,种群个体在群落中分布越均匀[22]。发耳清香木群落物种的频度等级有A、B、C、E 4个等级,以A级物种最多,占87.88%,共29种,超过一半的物种频度值仅为1%~5%,其中19种仅出现在1个样方中,占A级种的65.52%;其次是B级种2种,占606%;C级种1种,占3.03%;E级种1种(即清香木,其频度值为93.06%),占3.03%。说明在发耳清香木群落中,除清香木(频度值为93.06%)为优势种,分布较为均匀外,其余物种频度值多集中在1%~5%,分布极不均匀。该群落建群层(乔木层)物种较为单一,清香木占绝对优势,林下人类活动频繁,干扰强烈,清香木更新状况很差,群落中清香木的发展有衰退倾向。群落物种频度分布规律为A>B>C=E,群落没有D级物种,与Raunkiaer的频度定律不一致。都格清香木群落物种的频度也有A、B、C、D 4个等级,同样以A级物种最多,共52种,占94.55%,有25种仅出现在1个样方中,占A级种的45.45%;B、C、D 3级均仅有1个种,占1.82%,其中D级物种为清香木,频度值为77.78%。说明在都格清香木群落中,除清香木分布相对均匀外,另有82.69%的物种频度值仅为1%~5%,物种分布极不均匀。目前,该群落的建群层以清香木、复羽叶栾树为主要优势种,郁闭度较高,林下光照不足,清香木在林下更新层很少,幼年个体更新困难,该群落中清香木的发展同样有衰退倾向,有被其他物种替代的可能。该群落频度分布规律为A>B=C=D,群落没有E级物种,也与Raunkiaer的频度定律也不一致。2地的清香木群落频度分布值均与Raunkiaer的频度定律不一致,一方面可能与亚热带地区森林群落物种多样性、不均匀性有关;另一方面可能与2个群落皆为当地风水林,人们常常有目的地保护清香木,形成不同程度的气候顶极、干扰顶极有关。
2.4 物种多样性分析
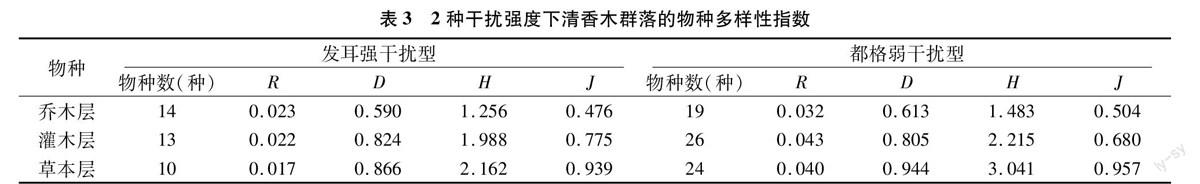
统计显示,发耳清香木群落各层物种数不多,草本层部分区域已被紫茎泽兰全部侵占,有的区域林下几乎没有其他植物生长,物种数很少,仅10种。根据中度干扰假说[23],中等程度的干扰水平对于维持物种多样性是有利的。发耳群落之所以物种多样性较低,可能是其临近村寨,林间有小道且林下人为、家禽家畜过度干扰等因素所致。中等程度的干扰水平,如林窗,特别是轻微的人畜干扰导致都格清香木天然林群落各层生长良好,种类较多,尤以灌木、草本层为典型,其中,都格乡灌木层物种数是发耳镇的1倍,都格乡草本层虽也有紫茎泽兰入侵,但数量较少,所以物种多样性较发耳镇高1倍多。这也可能是由都格清香木天然林位于村寨后山,人为干扰相对较少,除部分区域因火灾形成林窗,无大树外,其他位置林间郁闭度均较高等因素所致[24]。发耳镇清香木群落乔木层、灌木层、草本层物种数表现为乔木层>灌木层>草本层。都格清香木群落乔木层、灌木层、草本层物种数表现为灌木层>草本层>乔木层(表3)。
3 结论与讨论
本研究结果表明,强干扰型对发耳清香木群落物种多样性特别是灌木层、草本层影响较大,主要生活型的物种皆有分布,但物种丰富度不高,其他各指数与物种丰富度呈正相关。由于紫茎泽兰疯长,影响其更新层生长,因此更新层亦不发达。弱干扰对都格清香木群落物种多样性影响较小,群落垂直结构层次分明,林间郁闭度较高,林下光照不足,因此幼年个体更新情况不容乐观。但2个群落生活型组成均包含了Raunkiaer生活型系统的主要生活型,并且都以中高位芽植物占优势。2种干扰型的频度组成均与Raunkiaer频度定律不吻合,发耳清香木群落物种的频度组成为A、B、C、E 4个等级,其频度分布规律为A>B>C=E;都格清香木群落物种的频度等级为A、B、C、D 4个等级,其频度分布规律为A>B=C=D。一方面可能与亚热带地区森林群落物种多样性、不均匀性有关,另一方面可能与2个群落皆为当地风水林,人们常常有目的地保护清香木,形成不同程度的气候顶极与干扰顶极有关。由于处于亚热带气候区,水热条件较好,2种干扰强度下清香木林的植物种类组成较为丰富,但弱干扰条件下都格清香木群落各层的物种数都比强干扰条件下发耳高。研究表明,人为干扰强度对群落各层次物种多样性的影响不尽相同。人为干扰对乔木层而言,会使群落物种种类组成发生变化,不会使乔木层优势种的主体地位发生根本改变,但对灌木层、草本层的影响较大[25]。本研究结果表明,2种干扰类型的样地群落及其各层物种丰富度、多样性程度都不相同。都格清香木群落由于所受干扰较小,因此群落各层物种多样性相对较高,物种丰富度表现为灌木层>草本层>乔木层。发耳清香木群落由于人为活动频繁,乔木层物种数基本未受影响,但灌木层、草本层受影响较大,物种丰富度表现为乔木层>灌木层>草本层。自然地理位置差异和人为因素、家禽家畜的频繁活动等是造成2个群落物种组成、多样性特征差异的主要原因,人类干扰对群落建群层林木的林下更新影响很大。
参考文献:
[1]薛建辉. 森林生态学[M]. 北京:中国林业出版社,2006.
[2]汪殿蓓,暨淑仪,陈飞鹏. 植物群落物种多样性研究综述[J]. 生态学杂志,2001,20(4):55-60.
[3]王 妍,张 超,李 昆. 云南干热河谷地区清香木分布调查[J]. 林业资源管理,2013(3):156-160.
[4]王崇云,陈美卿,和兆荣,等. 记澜沧江河谷-硬叶小叶常绿阔叶林类型——锈鳞木樨榄、清香木群丛[J]. 植物分类与资源学报,2012,34(1):81-88.
[5]楊云花,朱春艳,蒋文智. 移植清香木衰弱原因及补救方法[J]. 农技服务,2012,29(9):1048.
[6]赵明荣. 野生清香木育苗技术[J]. 云南林业,2003,24(1):18.
[7]张国韩. 清香木在园林绿化中的应用[J]. 北京农业,2012(9):53-54.
[8]李亚萍,李家鹏,黄俊红. 清香木苗木调运及栽植技术[J]. 中国园艺文摘,2013(12):67-68.
[9]杨文才. 清香木荒山造林技术研究[J]. 绿色科技,2014(5):80-82.
[10]杨 静,石大兴,王米力,等. 清香木的组织培养与快速繁殖[J]. 植物生理学通讯,2007,43(3):523.
[11]贾利强,李吉跃,郎南军,等. 水分胁迫对黄连木、清香木幼苗的影响[J]. 北京林业大学学报,2003,25(3):55-59.
[12]赵湘江,王 妍,田 昆. 清香木叶片光合荧光特性对土壤水分胁迫的响应[J]. 干旱区资源与环境,2015,29(1):83-88.
[13]王 妍,张 超,李 昆. 干热河谷乡土树种清香木幼苗生长对不同水分梯度的响应[J]. 中南林业科技大学学报,2014,34(10):19-25.
[14]黄林敏,袁丛军,严令斌,等. 不同遮荫处理对清香木苗木生长与光合速率的影响[J]. 浙江农业科学,2014(2):217-219,224.
[15]乔永锋,彭永芳,方云山,等. 云南清香木绿叶和嫩红叶挥发性成分对比研究[J]. 安徽农业科学,2013,41(4):1583-1584,1587.
[16]刘布鸣,何开家,董晓敏,等. 野生清香木鲜叶挥发油的化学成分分析[J]. 国际医药卫生导报,2010,16(17):2086-2089.
[17]周葆华. 清香木叶与黄连木叶挥发油化学成分的对比[J]. 安庆师范学院学报:自然科学版,2008,14(2):60-62.
[18]周葆华. 清香木叶挥发油成分及其抑菌作用[J]. 应用化学,2008,25(3):305-308.
[19]郑 华,张 弘,李勇辉,等. 盆栽与露地清香木香气成分的差异[J]. 华北农学报,2009,24(增刊1):340-343.
[20]张金屯. 数量生态学[M]. 北京:科学出版社,2004.
[21]马克平,黄建辉,于顺利,等. 北京东灵山地区植物群落多样性的研究 Ⅱ. 丰富度、均匀度和物种多样性指数[J]. 生态学报,1995,15(3):268-277.
[22]杨 持. 生态学[M]. 北京:高等教育出版社,2012.
[23]马克平,刘灿然,于顺利,等. 北京东灵山地区植物群落多样性的研究 Ⅲ. 几种类型森林群落的种-多度关系研究[J]. 生态学报,1997,17(6):11-21.
[24]艾训儒,易咏梅,姚 兰,等. 旅游区人为干扰对森林群落物种多样性的影响[J]. 浙江林学院学报,2010,27(2):178-184.
猜你喜欢
山东工业技术(2017年5期)2017-03-16
山东农业科学(2017年2期)2017-03-15
绿色科技(2016年21期)2016-12-27
现代农业科技(2016年20期)2016-12-20
绿色科技(2015年6期)2015-08-05
