
光照强度对狐臭柴叶片形态与结构的影响
2015-07-01 08:02刘诗雅张明生高晓峰刘贵贤韦红边
贵州农业科学 2015年6期
刘诗雅,张明生,高晓峰,刘贵贤,韦红边,吕 享
(贵州大学 生命科学学院,贵州 贵阳550025)
狐臭柴(PremnapuberulaPamp.)系马鞭草科豆腐柴属多年生落叶灌木,又名神仙豆腐柴,分布于我国贵州、四川、广西、湖北、湖南和陕西等省区海拔700~1 800 m 山坡丛林中[1]。狐臭柴是一种药食兼用植物,其叶能清湿热解毒、治水肿毒疮,根具有调经壮阳、治月经不调、风湿关节炎和阳痿等功效[2-3];且叶片富含果胶、蛋白质、氨基酸和纤维素等营养成分[4-5],民间用叶揉碎制作清凉可口、色香味俱全的“神仙豆腐”,深受人们的喜爱,具有较大的经济利用价值。狐臭柴因其自然繁殖能力较差[4],加之生态环境被破坏,该物种野生资源日益减少,分布零散,采集困难[6],故开展其人工种植十分必要。狐臭柴叶片是主要的食用部位,作为光合作用的主要器官,是产生同化物的重要场所,直接影响植株生物量的形成[7]。光是植物生长的主要能源,也是影响植株叶片形态、解剖结构、生理代谢及生长发育的主要因子之一[8-13]。而目前尚无有关光照对狐臭柴叶片形态结构产生影响的研究报道。对此,笔者通过设置不同光照强度,探索其对狐臭柴叶片形态和解剖结构的影响,以期为狐臭柴人工种植技术的构建与开发利用提供理论依据。
1 材料与方法
1.1 材料
1.1.1 试验植物 狐臭柴,来源于贵州省务川仡佬族苗族自治县,由贵州大学鉴定。
1.1.2 遮阳网 不同目数的黑色遮阳网,市售。
1.2 试验设计
以不同目数的黑色遮阳网调节光照强度,分别设重度遮光、中度遮光和不遮光3种光照强度处理。处理1,重度遮光,透光率5%~10%;处理2,中度遮光,透光率40%~50%;处理3,不遮光为对照(CK)。使用85%的壤土混合15%的沙土作为栽培基质,以排水良好的塑料桶(桶高32cm,上口径35cm,下口径32cm)栽植狐臭柴一年生扦插苗,装土深度为27cm,每桶栽植1 株,每个处理栽植36桶。
1.3 叶片形态指标测定
于7月初采摘狐臭柴植株一级枝条健康的倒三叶,用FAA 进行固定,石蜡切片法制片,切片厚度10μm。用OLYMPUS-BX40显微镜在20倍和40倍物镜下观察并摄像,使用Photoshop 7.0以20倍和40倍物镜下的显微测微标尺为标准,对叶片厚度、叶脉直径、栅栏组织厚度、海绵组织厚度、上下表皮厚度及上下表皮细胞密度等进行统计(表皮装片为撕取上、下表皮制作的临时装片),叶片形态指标按照植物学常规描述。
1.4 数据处理
以Excel和DPS统计软件进行数据分析。
2 结果与分析
2.1 光照强度对狐臭柴叶片形态的影响
从表1和图1看出,随着光照强度的增加,叶片表面积(叶片大小)逐渐减小;叶形由心形、近心形过渡到卵形;叶缘由锯齿、部分锯齿到全缘;叶尖由长尾尖、尾尖到急尖;叶色由深绿色、绿色到浅绿色;叶质由纸质逐渐变为坚纸质。说明,光照强度是狐臭柴生长的重要环境因子,荫蔽条件下叶面积增大(包括锯齿、心形、长尾尖)、叶色变深(叶绿素含量增加)、叶质变软(角质层减少)等均是狐臭柴植株对阴生环境的良好适应。
2.2 不同光照强度下狐臭柴叶片的解剖结构
狐臭柴叶片上、下表皮各有一层细胞,细胞大小不等,形状不规则,排列紧密,上表皮气孔极少,下表皮气孔较多,上、下表皮均有表皮毛和腺体(图2~3)。遮光处理下叶片的上表皮细胞形态与对照差异较大(图2-A~C),而下表皮细胞形态在不同处理间无明显差异(图2-D~F)。
不同光照强度下狐臭柴叶片组织结构有不同程度的变化(表2,图3~4)。上表皮细胞密度、叶脉直径随光照强度的增加呈极显著正相关,而下表皮细胞密度受光照强度变化的影响不明显;叶片总厚度、上表皮厚度、栅栏组织厚度和海绵组织厚度在2种遮光条件下变化不大,但与对照均有极显著差异,而叶片下表皮厚度受光照强度的影响很小。狐臭柴是典型的阴生植物,强光条件下,上表皮变厚且细胞密度增大、气孔极度减少,有利于降低水分蒸腾;栅栏组织和海绵组织增厚,有助于充分回收光呼吸释放的胞间CO2;叶脉变粗,可保障光合产物的瞬时转运,有效地降低光呼吸消耗。

表1 不同光照强度下狐臭柴叶片的形态指标变化Table 1 Variation of leaf morphology indices under different sunlight intensity

图1 不同光照强度下狐臭柴叶片的形态变化Fig.1 Leaf shape changes under different sunlight intensity

表2 不同光照强度下狐臭柴叶片的组织结构变化Table 2 Changes of leaf tissue structure under different sunlight intensity

图2 不同光照强度下狐臭柴叶片的表皮细胞形态Fig.2 Leaf epidermal cell morphology under different sunlight intensity

图3 不同光照强度下狐臭柴叶片的解剖结构(横切)Fig.3 Leaf anatomical structure under different sunlight intensity(transection)
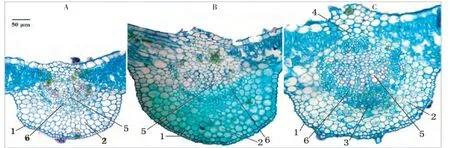
图4 不同光照强度下狐臭柴叶脉的解剖结构(横切)Fig.4 Leaf vein anatomical structure under different sunlight intensity(transection)
3 小结与讨论
1)研究结果表明,随着光照强度的增加,狐臭柴叶片表面积(叶片大小)逐渐减小;叶形由心形、近心形过渡到卵形;叶缘由锯齿、部分锯齿到全缘;叶尖由长尾尖、尾尖到急尖;叶色由深绿色、绿色到浅绿色;叶质由纸质逐渐变为坚纸质。表明,狐臭柴属于阴生植物的特征。
2)地球上的每一种植物,在长期进化过程中形成了适应于特定自然环境、有利于自身生长发育的形态结构,而且这些特定的形态结构会随着环境的变化而发生一定的改变,这就是植物的生态适应性。本研究所用的狐臭柴属于阴生植物,光照强度是影响其生长发育的重要生态因子,强光下,其叶面积减小,叶片上表皮角质增加、细胞密度增大、栅栏组织和海绵组织增厚,这些变化充分反映了狐臭柴对光逆境的适应性,也符合大多数植物在高光强下叶片解剖结构的变化[14-18]。然而,植物的逆境适应能力有限,例如,适度的强光照可促进叶片表面角质层的形成,进而提高叶片反光度以减轻光对叶肉细胞的灼伤,同时通过薄膜干涉和改变光反射方式影响光量子吸收[19];若光照过强,则使叶绿体变小、类囊体结构破坏,光合作用强烈受阻而导致植株死亡[20-23]。
[1]中国科学院植物研究所.中国高等植物图鉴[M].北京:科学出版社,1985:87-88.
[2]中国科学院中国植物志编辑委员会.中国植物志:第65卷[M].北京:科学出版社,1982:86-87.
[3]徐 利,史梦娜,张明生.狐臭柴扦插繁殖技术初步研究[J].山地农业生物学报,2011,30(1):82-86.
[4]史梦娜,张明生,徐 利,等.溶液培养对狐臭柴植株生长及叶片神仙豆腐的影响[J].中南林业科技大学学报,2012,32(03):70-74.
[5]Zhang M S,Shi M N,Xu B R,et al.A key component determination on forming Fairy Tofu from the leaf of Premna Puberula(Verbenaceae)[J].Journal of Applied Pharmaceutical Science,2012,2(9):31-35.
[6]许邦锐,张明生,李祥栋,等.光照强度对狐臭柴植株生长及叶片生理生化指标的影响[J].中南林业科技大学学报,2013,33(12):82-85,95
[7]赵立华,胡中会,李成云,等.不同遮光程度对玉米叶片结构的影响[J].中国 农 学通报,2012,28(6):43-46.
[8]Philip M P,Fraser J G M.Effects of shade on growth biomass allocation and leaf morphology in European yew (Taxus baccata L.)[J].European Journal of Forest Research,2013,132:211-218.
[9]Zhang S B,Yin L X.Plasticity in photosynthesis and functional leaf traits of Meconopsis horridula var.racemosa in response to growth irradiance[J].Botani-cal Studies,2012,53:335-343.
[10]Lusk C,Reich P B,Montgomery R A,et al.Why are evergreen leaves so contrary about shade[J].Trends in Ecology and Evolution,2008,23:299-303.
[11]许邦锐,张明生,李祥栋,等.光照强度对狐臭柴植株生长及叶片生理生化指标的影响[J].中南林业科技大学学报,2013,33(12):82-85,95.
[12]Chen J W,Zhang Q,Li X S,et al.Steady and dynamic photosynthetic responses of seedlings from contrasting successional groups under low-light growth conditions[J].Physiologia Plantarum,2011,141:84-95.
[13]李芳兰,包维楷.植物叶片形态解剖结构对环境变化的影响与适应[J].植物学通报,2005,22(增刊):118-127.
[14]江 莎,胡 阳,郑书馨,等.不同光强与光质对“达赛莱克特”草莓叶片形态结构的影响[J].电子显微学报,2009,28(5):453-461.
[15]史刚荣,蔡庆生.白三叶叶片解剖可塑性及其对光强的响应[J].草地学报,2006,14(4):301-305.
[16]Ashton P M S,Berlyn G P.Leaf adaptation of some Sharea species to sun and shade[J].New Phytologist,1992,121:587-596.
[17]Ashton P M S,Berlyn G P.A comparison of leaf physiology and anatomy of Quercus(Section Erythrobalanus-Fagaceae)species indifferent light environments[J].American Journal of Botany,1994,81(5):589-597.
[18]Crystalarens N.Response of leaf anatomy to light environments in the tree fem Cyahtea caracasana(Cyatheaceae)and its application to some ancient seed fem[J].Palaios,1997,12:84-94.
[19]Wolfgang B,Olle B,Susan S T.Light-induced spectral absorbance changes in relation to photosynthesis and the epoxidation state of xanthophyll cycle components in cotton leaves[J].Plant Physiology,1989,91:542-551.
[20]Tiffany A M,Mark H E,Casey W B,et al.Differences in chloroplast ultrastructure of Phaeocystis antarctica in low and high light[J].Marine Biology,2006,149:1281-1290.
[21]Wheeler W S,Fagerberg W R.Exposure to low levels photosynthetically active radiation induces rapid increases in palisade cell chloroplast volume and thylakoid surface area in sunflower(Helianthus annuus L.)[J].Protoplasma,2000,212:38-45.
[22]王菊凤,李鹄呜,廖飞勇,等.光照对盾叶薯蓣荧光光谱和叶绿体结构的影响[J].吉首大学学报,2006,27(1):80-85.
[23]陈章和,朱素琴,李韶山,等.UV-B 辐射对东南亚热带森林木本植物幼苗生长的影响[J].云南植物研究,2000(22):467-472.
猜你喜欢
青春期健康(2022年24期)2022-12-26
建材发展导向(2021年14期)2021-08-23
祝您健康·文摘版(2019年4期)2019-06-11
祝您健康·文摘版(2018年9期)2018-05-14
上海农业学报(2017年4期)2017-04-10
中国中西医结合皮肤性病学杂志(2016年4期)2016-07-18
中外文摘(2016年2期)2016-03-22
中国房地产业(2016年9期)2016-03-01
中国果业信息(2015年12期)2015-01-24
中学生理科应试(2014年2期)2014-04-23
