
茅台酒厂生态功能区优势植物的生理代谢及土壤理化性状
2015-07-01 08:01张习敏张宇斌
贵州农业科学 2015年6期
张习敏,申 刚,张宇斌,陈 玲,洪 鲲,乙 引*
(1.贵州师范大学 生命科学学院,贵州 贵阳550001;2.贵州省植物生理与发育调控重点实验室,贵州 贵阳550001)
植物需要从土壤溶液中吸收矿质元素以维持自身生长和生理代谢,植物吸收的矿质元素从数量上来说,有的矿质元素数量较大(如N、P、K、Ca、Mg),有的矿质元素的数量较少(如Fe、Cu等)[1-2]。影响植物吸收土壤矿质元素的因素很多,如植物生长状况、树木年龄、大气温度、光照条件和土壤因素等。从土壤因素来看,影响植物根系吸收矿质元素的因素有土壤温度、土壤通气状况、土壤溶液浓度、土壤pH、土壤含水量和土壤微生物等。其中,微生物在土壤养分和有机质转化中具有重要作用,是土壤有机质和土壤养分(如N、P、K、S等)转化和循环的动力之一。同时参与土壤有机质、腐殖质和林地枯枝落叶的分解、养分的转化和矿质营养的生物地球化学循环中的某些生化过程,在生态系统能量流动和养分转化中也起着重要的作用,是维持和恢复林地生产力的主要因素之一[3-8]。土壤微生物还影响土壤的理化性质,如有机质、pH 等。在森林土壤中,细菌、真菌和放线菌3大类微生物的数量通常作为评价森林土壤生物活性高低的重要标志,其分布广度、种群和数量直接影响林地土壤肥力和理化性质,进而影响林木的生长[9-13]。
贵州茅台酒厂坐落于贵州省仁怀市赤水河畔(E106°21',N27°51'),海拔约423 m,三面环山。年均温17.4℃,气温年差较大,1月均温6.9℃,7月均温27.9℃。夏季最高气温达39.9℃,炎热季节持续半年以上;冬季气候暖和,温差小,霜期短,年平均无霜期达326d,最低气温为2.7℃。干热少雨,年雨量仅有800~1 000 mm,日照丰富,年日照可达1 400h。土壤为紫色土,土层较厚,一般在50cm左右。该地区的植被覆盖率低,群落结构单一,主要优势物种有狗尾草(Setariaviridis)、箬竹(Indocalamustessellatus)、三叶鬼针草(Bidenspilosa)、复羽叶栾树(Koelreuteriabipinnata)、构树(Broussonetiapapyrifera)和青冈栎(Cyclobalanopsis glauca)。该地区主要优势物种吸收的大量元素是否存在差异,土壤微生物和土壤理化性状是否对这些物种吸收矿质元素有影响,探明这些问题可以帮助了解该地区植被的矿质营养代谢的多样性和生长状况。然而,与此相关的研究未见报道。因此,需要对该地区优势物种的矿质元素吸收及相关土壤因子进行调查分析。为进一步评价该生态功能区森林生态系统的现状和发展趋势,对茅台酒厂生态系统的维护与重建具有重要的指导意义。
1 材料与方法
1.1 样地选择
样地在茅台酒厂上游约2 000m 的取水口、茅台酒厂河对岸、茅台酒厂下游约1 000m,在每个样地中,随机设置2个20m×20m 的样方,分别记为MTS-01和MTS-02、MTZ-01和MTZ-02、MTX-01和MTX-02(图示)。记录各样方的经纬度、海拔、坡度、坡向、地貌类型、表层岩石、土壤类型、光照强度、温度、空气湿度、风速和郁闭度等生境指标(表1)。
1.2 植物及土壤样品采集
在茅台酒厂生态功能区上中下游的调查样地内,选择6种优势树种青冈栎、箬竹、三叶鬼针草、狗尾草、复羽叶栾树和构树,分别取其根系、茎和叶片,青冈栎、复羽叶栾树、箬竹和构树每个物种选5株进行取样并混合,三叶鬼针草和狗尾草则选择5 个1m×1m 区域生长的物种,按根、茎和叶分开采取,并将其混合。分别在每个样地中的优势物种根际周围取0~20cm、20~40cm 和40~60cm 土层土样。

图示 茅台酒厂生态功能区样地调查分布Fig.Sample plots distribution in ecological function area of Moutai distillery

表1 茅台酒厂生态功能区样地生境的基本情况Table 1 Basic situation of sample plot habitats in ecological function area of Moutai distillery
1.3 测定指标及方法
1)植物营养元素含量。植物样品置于80℃烘箱内烘干至恒重后,将根、茎和叶分别粉碎,取其粉末各1g,测定N、P、K、Ca、Mg的含量。总氮测定采用凯氏定氮法,总磷用钼锑抗比色法,K、Ca、Mg经烘干灰化后以1 mol/LHCl消解,用原子吸收分光光度计测定。新鲜土样带回实验室后,4℃冰箱保存,一部分样品用于测定土壤微生物,另一部分用于测定理化性状。
2)土壤理化指标。经风干、磨细、过60 目筛后,用于测定土壤有机碳(采用重铬酸钾外加热法);容重测定采用环刀切割法;比重采用排水法测定、pH 采用酸度计直接测定(V(水):M(土)=2.5∶1)、总氮测定采用凯氏定氮法;总磷采用HClO4-H2SO4消解后钼锑抗比色法;K、Ca、Mg经1mol/L HCl消解后,采用原子吸收分光光度计测定[14]。
3)土壤微生物数量。细菌采用牛肉膏蛋白胨培养基,真菌采用马铃薯琼脂糖培养基,放线菌采用高氏1号培养基,各处理重复3次。各类菌接种后置恒温培养箱中培养,细菌在28~30℃培养36~72h后采用稀释平板法计数,放线菌置28℃培养7d后计数,真菌25℃培养5d后计数。土壤微生物数量均换算成干土中的含菌数(个/g)[15-16]。
1.4 数据分析
采用SPSS18.0软件对数据进行分析处理,对土壤理化性质、微生物和矿质营养之间进行相关性分析(Pearson检验,P<0.05)。
2 结果与分析
2.1 主要优势物种的矿质营养元素含量
在6种优势植物中,5种主要矿质元素含量之间存在极大差异(表2)。K 和Ca含量整体高于P和Mg。N、P、K、Ca和Mg含量基本叶片含量最高、根部含量最低。N 含量最高的是复羽叶栾树(3.63%),最低的是狗尾草(1.18%);P含量最高的是三叶鬼针草(2.35g/kg),最低的为箬竹(0.79g/kg);K 含量最高的是三叶鬼针草(22.93g/kg),最低的是构树(3.95g/kg);Ca含量最高的是三叶鬼针草(31.80g/kg),最低的是构树(3.73g/kg);Mg含量最高的是三叶鬼针草(9.81g/kg),最低的是狗尾草(1.34g/kg)。
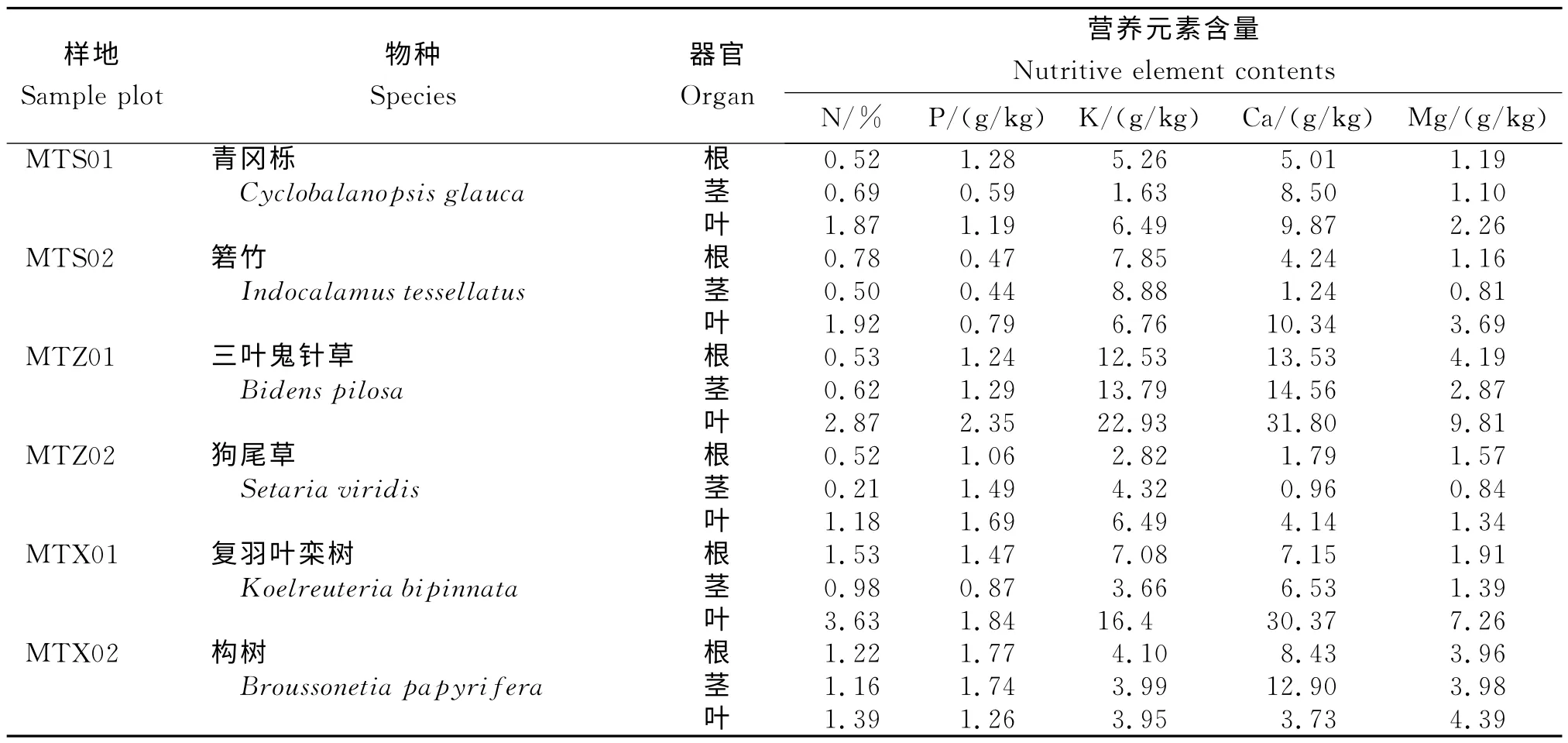
表2 茅台酒厂生态功能区优势物种器官营养元素含量Table 2 Nutritive element contents in dominant species in ecological function area of Moutai distillery
2.2 主要优势物种根际土壤的理化性状
茅台酒厂生态功能区上游土壤容重、土壤pH和土壤比重较低,中游和下游较高。表明,上游土壤偏酸性,土壤较松弛;中游和下游土壤偏碱性,土壤较坚固。同时,上游土壤有机质的含量均高于中游和下游(表3)。
2.3 主要优势物种根系土壤微生物数量
茅台酒厂生态功能区中游土壤的微生物总数最多。其中,中游细菌数量最多,约6×104个;放线菌下游数量最多,约为6×103个;3个区域的真菌数量差异不大(表4)。
2.4 相关性
几种优势植物的矿质元素(N,P,K,Ca,Mg)含量均与土壤容重、pH、比重和放线菌呈正相关。其中,P元素含量与土壤pH 显著相关(P<0.05)。土壤矿质元素含量与有机质和真菌数量呈负相关;N和Ca与细菌呈负相关,P、K 和Mg与细菌呈正相关(表5)。
3种微生物中,细菌和放线菌与容重、比重和pH 呈正相关,与有机质呈负相关;真菌与容重和比重呈负相关,与pH 和有机质呈正相关(表6)。

表3 茅台酒厂生态功能区主要优势物种根际土壤的理化性状Table 3 Physical and chemical properties of rhizosphere soil in ecological function area of Moutai distillery

表4 茅台酒厂生态功能区主要优势物种根际土壤的微生物数量Table 4 Microbial Population in ecological function area of Moutai distillery

表5 营养元素含量与土壤理化性状和微生物数量的相关性Table 5 Correlation of nutritive element content with soil physicochemical properties and microbial population

表6 土壤微生物数量与理化性状的相关性Table 6 Correlation of soil microbial population with physicochemical properties
3 结论与讨论
在茅台酒厂生态功能区内,主要优势物种有狗尾草(S.viridis)、箬竹(I.tessellatus)、鬼针草(B.pilosa)、复 羽 叶 栾 树 (K.bipinnata)、构 树(B.papyrifera)、青冈栎(C.glauca)。虽然均位于茅台酒厂生态功能区内,但是分布的具体位置不同,位于不同的土壤环境中。因而物种对土壤营养元素的吸收不同,具体表现在不同物种营养元素含量之间存在差异。矿质营养对植物的生长至关重要,每个物种需要一定比例的营养元素含量[17]。
土壤中矿质元素的可用性是影响植物对矿质元素吸收的重要因素,矿质元素的可用性受土壤母质、气候、风化程度和淋溶作用等影响[18]。同时,土壤理化性质也影响土壤中元素活度。席冬梅等研究发现,土壤pH 值影响土壤矿物质的风化强度、土壤有机质的转化、土壤中微生物的活动,同时,pH 还影响土壤溶液中化合物的溶解及沉淀,无机离子的吸附和交换。因此,植物对土壤中矿质元素的吸收严重受到土壤pH 的影响[19-20]。在茅台酒厂生态功能区内,上游、中游和下游的土壤理化性质存在差异,上游土壤呈酸性,pH 值低,上游的物种多样性优于中游和下游,同时,物种的多样性加剧了矿质元素的生物地球化学循环,尤其是土壤中P的吸收。由于矿质元素的生物地球化学循环加剧,因而可能影响土壤中微生物的数量,但研究中发现,中游土壤的微生物总量高于上游和下游。这可能是中游距离厂区较近,茅台酒厂厂区大气中微生物直接侵入土壤,导致土壤中的微生物数量增加,但是,具体导致土壤微生物差异的原因需要进一步研究。
[1]Agren G I.Stoichiometry and nutrition of plant growth in natural communities[J].Annual Review of Ecology Evolution and Systematics,2008,39:153-170.
[2]White P J,Brown P H.Plant nutrition for sustainable development and global health[J].Annals of Botany,2010,105:1073-1080.
[3]胡婵娟,傅伯杰,刘国华,等.黄土丘陵沟壑区典型人工林下土壤微生物功能多样性[J].生态学报,2009,29(2):727-733.
[4]Harris J A,Birch P.Soil microbial activity in opencast coal mine restoration[J].Soil Use and Management,1989,5:155-160.
[5]王锐萍,刘 强,彭少麟,等.尖峰岭不同树种枯落物分解过程中微生物动态[J].浙江林学院学报,2006,23(3):255-258.
[6]李延茂,胡江春,汪思龙,等.森林生态系统中土壤微生物的作用与应用[J].应用生态学报,2004,15(10):1943-1946.
[7]Schipper L A,Degens B P,Sparling G P.Changes in microbiai heterotrophic diversity along five plant successional sequences[J].Soil Biology and Biochemistry,2001,33:2093-2103.
[8]赵 萌,方 晰,田大伦.第2代杉木人工林地土壤微生物数量与土壤因子的关系[J].林业科学,2007,43(6):7-12.
[9]宋 漳,朱锦懋,杨玉盛.闽北常绿阔叶林土壤微生物学特性的研究[J].福建林学院学报,2000,20(4):317-320.
[10]徐秋芳,姜培坤,陆贻通.不同施肥对雷竹林土壤微生物功能多样性影响初报[J].浙江林学院学报,2008,25(5):548-552.
[11]马晓梅,尹林克,陈 理.塔里木河干流胡杨和柽柳根际土壤微生物及其垂直分布[J].干旱区研究,2008,5(2):183-189.
[12]Torsrik V L,Goksoyr J,Daae F L.High diversity in DNA of soil bacterial[J].Applied and environmental Microbiology,1999,56(3):782-787.
[13]Sonu S,Nandita G,Singh K P.Variations in soil microbial biomass and crop roots due to differ in resource quality inputs in a tropical dry land agroecosystem[J].Soil Biology and Biochemistry,2007,39:76-86.
[14]鲍士旦.土壤农化分析[M].北京:中国农业出版社,2000.
[15]许光辉,郑洪元.土壤微生物分析方法手册[M].北京:农业出版社,1986.
[16]中国科学院南京土壤研究所微生物室.土壤微生物研究法[M].北京:科学出版社,1985.
[17]贾文龙,邢国明.土壤矿质元素含量与古侧柏生存状态的相关性[J].山西农业科学,2010,38(9):45-47.
[18]连玉武.朱小龙,李利峰.三明小湖赤枝栲林地部分矿质元素及土壤理化性质研究[J].福建林业科技,2002,29(3):10-13.
[19]席冬梅,邓卫东,高宏光,等.云南省主要地质背景区土壤理化性质及矿物质元素丰度分析[J].土壤,2008,40(1):114-120.
[20]潘琼蓉.马尾松一代种子园土壤诊断研究[J].山地农业生物学报,2014(5):13-18.
猜你喜欢
山东冶金(2019年3期)2019-07-10
中成药(2018年11期)2018-11-24
民主与法制(2018年35期)2018-09-19
民主与法制(2018年26期)2018-07-17
现代园艺(2017年23期)2018-01-18
现代园艺(2017年21期)2018-01-03
中国水利(2015年5期)2015-02-28
植物营养与肥料学报(2012年1期)2012-10-26
植物营养与肥料学报(2011年3期)2011-10-24
中国工程咨询(2011年8期)2011-02-16
