
小秦岭国家级自然保护区乔木生物量随地形因子变化格局
2015-06-27 03:47:47贾宏汝韩军旺叶永忠黄群策袁志良河南财政税务高等专科学校河南郑州6河南农业大学生命科学学院河南郑州000河南农业大学林学院河南郑州000河南小秦岭国家级自然保护区管理局河南灵宝700郑州大学离子束生物工程省重点实验室河南郑州00
河南农业大学学报 2015年6期
贾宏汝, 张 旭, 陈 云, 韩军旺, 叶永忠, 黄群策, 袁志良(.河南财政税务高等专科学校,河南 郑州6;.河南农业大学生命科学学院, 河南 郑州 000;.河南农业大学林学院, 河南 郑州 000;.河南小秦岭国家级自然保护区管理局, 河南 灵宝 700;.郑州大学离子束生物工程省重点实验室, 河南 郑州 00)
小秦岭国家级自然保护区乔木生物量随地形因子变化格局
贾宏汝1, 张 旭2, 陈 云3, 韩军旺4, 叶永忠2, 黄群策5, 袁志良2
(1.河南财政税务高等专科学校,河南 郑州451464;2.河南农业大学生命科学学院, 河南 郑州 450002;3.河南农业大学林学院, 河南 郑州 450002;4.河南小秦岭国家级自然保护区管理局, 河南 灵宝 472500;5.郑州大学离子束生物工程省重点实验室, 河南 郑州 450052)
在河南省小秦岭地区设置56个样方进行调查,根据树木异速生长方程,探讨了小秦岭森林乔木地上总生物量随地形因子的变化格局,并研究了该区物种多度最大的华山松生物量随海拔梯度的变化格局。结果表明:(1)地上生物量与海拔因子在统计学上具有显著相关性;(2)乔木地上生物量随海拔上升呈现出先升高后下降的趋势;(3)物种多度最大的华山松的地上生物量随海拔上升呈现逐渐下降的趋势,这与总体乔木地上生物量的变化格局有一定差别,可能是由于华山松生长在较高海拔不占优势;(4)华山松各器官生物量均随海拔先上升后波动性下降,与华山松生物量随海拔变化一致;(5)华山松地上生物量与地下生物量变化趋势一致,其比值在3.1~3.2,随海拔升高呈现出逐渐增高的趋势。
小秦岭;生物量;异速生长方程;海拔
森林生物量的大小和变化是森林植被生物学、生态学及其他自然因素与区域人类活动综合作用的表现,是衡量森林生态系统结构和功能变化的重要指标之一[1]。因此,准确地估算区域森林的生物量对研究区域陆地生态系统的生产力、营养元素格局和生物质能积累具有十分重要的意义,并对进一步研究森林群落的结构、功能和评价森林的可持续经营提供科学依据。在近几年碳汇储备研究的大背景之下,生物量的研究已经成为热点。对森林生物量的研究,不仅有助于了解森林生态系统的物质循环和能量流动,而且能为森林的规划和物种的保护提供重要的理论基础,同时也能更好地评估人类经营活动和全球气候变化对森林的影响。大量学者对水分梯度、林龄、月份等因素影响生物量分配格局做了不少研究[2-5],此外针对地形因子(海拔、坡位、坡向)对生物量的影响也做出了很多研究[6-9]。林敦梅等[10]以古田山国家级自然保护区为样地对中国亚热带常绿阔叶林地上生物量进行的研究发现,地形变化对生物量的分布具有一定程度的影响。这些基于生物量对森林群落的研究,无疑对森林生态的研究与保护产生了积极的影响。
河南省小秦岭国家级自然保护区的森林研究多以资源调查、生物多样性及物种空间格局的分布为主[11-12],而以生物量的变化格局为基础对森林群落的探讨较为少见。基于此背景,本研究参考秦岭火地塘林区生物量研究[13-14],选择合适的地形并设置一定数量的样方,对河南小秦岭乔木地上生物量随地形因子变化格局进行研究。对研究区乔木地上总生物量变化格局的研究,可以反映出整体群落的一般特性,但整体的一般性在一定程度上掩盖了局部的特殊性,因此研究小群落的变化格局也非常有必要。华山松主要分布于海拔1 600~1 750 m的垂直范围,和锐齿槲栎组成针阔叶混交林,本研究选取了样地中多度最大的物种华山松,对其生物量随海拔变化进行研究,以便与样地乔木地上总生物量的变化格局进行对比,说明样地群落整体的一般性及局部的特殊性。该对比研究对保护区的规划建设及物种的保护,具有深远的影响。
以往对森林生物量的估计大多采用直接收获法,此法费时费力且不具有连续性,而且对森林的破坏性较大,因此国内外众多学者对生物量计算做了大量的探索研究[15-17]。现今对森林生物量的研究都是通过样地调查法,调查典型样地的树木物种、树高及胸径,采用相关曲线法、构建模拟法、皆伐实测法、标准木法计算生物量。使用最为广泛的是异速生长方程,其中以树高(H)和胸径(D)来估算生物量的异速生长关系最为常见。需要注意的是,大部分物种生物量是以树高(H)和胸径(D)2个变量来估算的,但也有部分物种以胸径(D)作为单变量来估算生物量比较合适。考虑到对森林树木的保护,本研究采用异速生长方程进行生物量估算,所用方程均来自该区或类似生境地区。另外对于地上生物量和地下生物量的关系,国内学者对此做出了大量研究[18-19],因此本研究针对样地内物种多度最大的华山松的地上生物量和地下生物量的关系也做了分析。
1 研究区概况和研究方法
1.1 研究区及样地概况
河南小秦岭国家级自然保护区位于豫陕2省交界的灵宝市西部、小秦岭北麓;东接崤山丘陵,西连秦岭主脉,南倚莽莽群山,北濒涛涛黄河;地理坐标为北纬34°23′~34°31′,东经110°23′~110°44′;本区地处暖温带向亚热带的过渡区,为暖温带大陆性季风型半干旱气候,四季分明。最高海拔2 413.8 m,年平均气温依不同的海拔,变化在5.5~14.2 ℃之间,极端最低气温为-17.0 ℃,极端最高气温为42.7 ℃;年均降水量在506~719.2 mm,土壤类型南坡是棕壤、黄棕壤,北坡是褐土[11-12]。在研究区内选择具有代表性的山体,依据海拔梯度的变化,在海拔1 020~2 413 m的范围内,随海拔每上升50 m在南北坡各选择合适的群落设置1个20 m×20m的乔木样方,共设置56个20 m×20 m的乔木样方。所选样地植物群落中的乔木主要由华山松(Pinusarmandii)、领春木(Eupteleapleiosperma)、东陵山柳(Salixphylicifolia)、漆树(Toxicodendronvernicifluum)、葛萝槭(Acergrosseri)、油松(Pinustabulueformis)、糙皮桦(Betulautilis)、蒙古栎(Quercusmongolica)、秦岭冷杉(Abieschensiensis)、槲栎(Quercusaliena)等物种组成。
1.2 数据调查与数据处理方法
用GPS定位测量56个样方海拔,用测高仪测量树高,用钢围尺测量树木的胸径,并记录每个样方海拔、坡度、坡向和各样方树木编号、物种名、树高、胸径。查阅相关文献选择本研究区或相似生境地区的各物种的异速生长方程,通过树高和胸径与各器官的异速生长方程来算得相应器官的生物量,然后累加各器官的生物量得到地上生物量。对于其他物种,采用广义异速生长方程计算。本研究各物种所用到的异速生长方程如表1。本研究采用R语言进行统计计算分析和绘图分析。

表1 小秦岭乔木地上生物量的异速生长方程Table 1 Allometric equations of above-ground biomass of Xiaoqinling
注:表中H表示树高,D表示胸径,WS表示树干生物量,WB表示树枝生物量,WL表示树叶生物量,WR表示树根生物量,AGB表示地上总生物量。
Note:H represents the tree height,D=diameter at breast height,WS=trunk biomass,WB=branch biomass,WL=leaf biomassand,WR=the root biomass,AGB=total biomass on the ground.
2 结果与分析
2.1 三地形因子与生物量的相关性测验
用R语言分别对3地形因子(海拔、坡度、坡向)与生物量的相关性进行统计测验,分析结果见表2。
通过地形因子与生物量相关性检测结果表明,海拔梯度与生物量的变化相关性极为显著(P<0.01),而坡度和坡向对生物量的变化在统计学上不具有明显的相关性。因此,研究海拔与生物量的变化关系,对森林的可持续经营和调整林分结构具有重要意义。
2.2 地上生物量随海拔梯度的变化格局
基于海拔与生物量的相关性,通过R语言作图,拟合出海拔与地上生物量的线性回归图形,如图1。从图1中可以看出,基于散点图的曲线拟合较好,反映出在海拔1 000~2 000 m,随海拔上升,地上生物量呈现增长趋势;而在海拔2 000 m以上,随海拔上升,地上生物量呈现波动型下降趋势;在海拔1 600~2 000 m生物量较高并出现峰值。

表2 地形因子与生物量的相关性测验Table 2 The correlation test of topographical factors and biomass
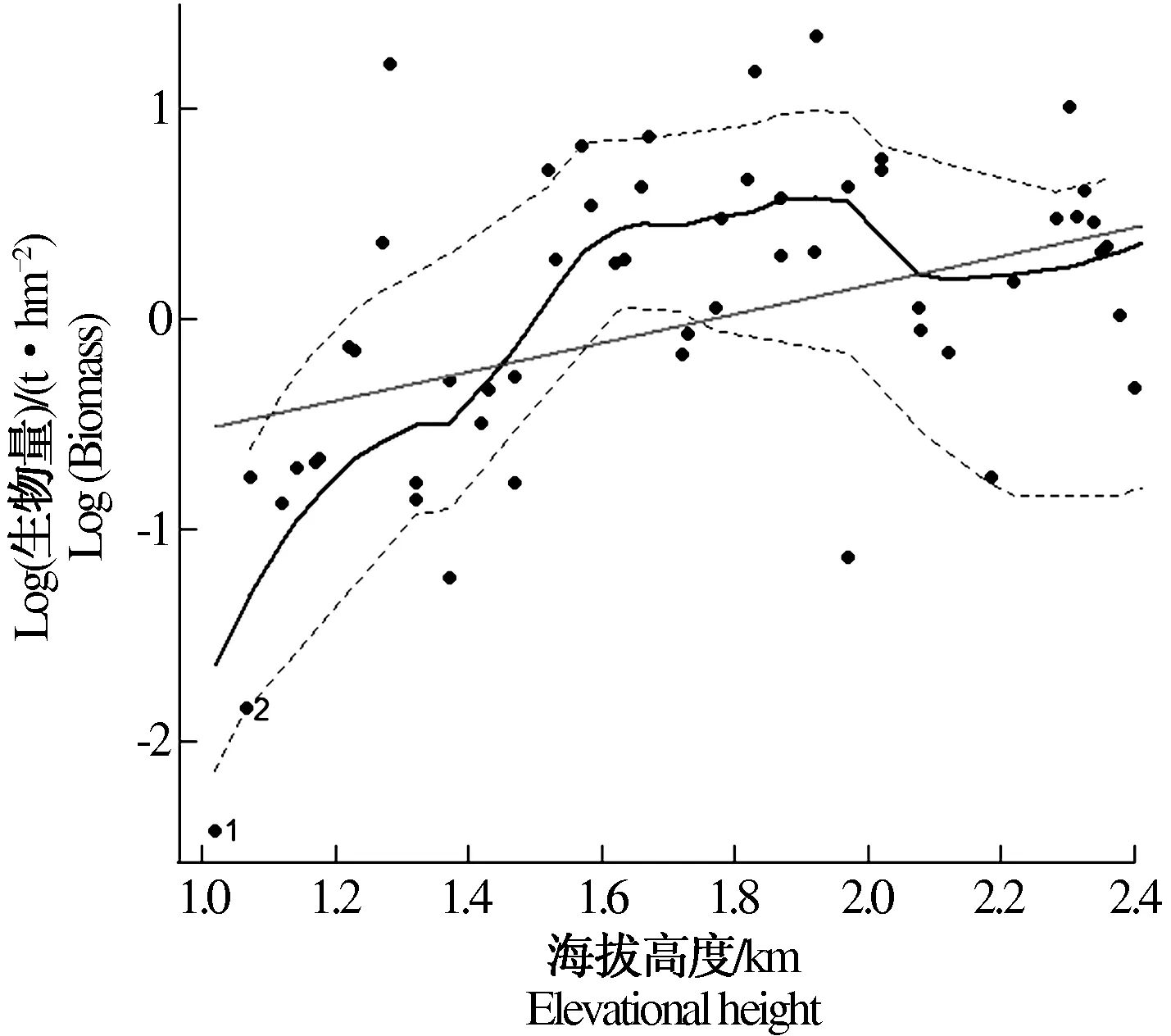
图1 地上生物量随海拔梯度的变化格局Fig.1 Changes in the pattern of above-ground biomass with elevation gradient
随海拔梯度的升高,立地因子(土壤、温度、湿度、光照、生物影响)都会有较大的变化,进而会影响树木的固碳及吸收营养物的能力,最终导致树木的生存生长受到影响。图1中乔木地上生物量随海拔梯度的变化呈现出先上升后下降的整体趋势,这与蔺恩杰等[6]对天目山自然保护区典型森林植被乔木层生物量的研究结果有一定差距,这是由于小秦岭国家自然保护区样地中等海拔的土壤水肥适宜,受人为干扰的影响较小,且相对紧张的资源环境有利于物种间竞争生长。
2.3 华山松生物量随海拔梯度的变化格局
2.3.1 华山松各器官生物量随海拔梯度的变化格局 华山松作为样地内物种多度最大的乔木树种,利用其各器官生物量的异速生长方程(表1),估算出沿海拔1 020~2 413 m分布的每棵华山松的各器官生物量,并运用R语言进行绘图,分析华山松各器官生物量随海拔梯度的变化格局,结果如图2。从图2可以看出,华山松各器官生物量随海拔上升均呈现出先略微上升后或保持稳定,海拔1 400 m以上又逐渐下降的趋势,说明随海拔的上升所带来的其他环境因子的变化,对华山松所有器官均产生了不利影响,造成了较高海拔物种生物量整体变少的趋势。
干、枝、叶和根生物量变化趋势均呈现先略微上升(或保持稳定后)又逐渐下降的趋势,证明植物器官之间的生长表现出既相互依赖又相互制约的关系。

图2 华山松各器官生物量随海拔梯度的变化格局Fig.2 Patterns of armandi biomass change with elevation gradient
2.3.2 华山松地下与地上生物量随海拔梯度的变化格局 利用华山松各器官生物量的异速生长方程,计算华山松地上生物量和地下生物量,并运用R语言绘图分析,通过线性回归图形分别分析地上生物量、地下生物量与海拔梯度的变化关系,结果如图3和图4。
由图3可知在低海拔的样方区域,华山松的生长状况较为良好,生物量较高,随海拔的升高,生物量逐渐减少。对比图3与图1中地上生物量随海拔分布格局,结果表明,研究区单个物种华山松地上生物量随海拔梯度变化呈现出先略微上升后逐渐下降的趋势,与样地中所有乔木地上总生物量随海拔梯度的变化格局不完全一致,这种差异可能是由于华山松的生物量在较低海拔1 000~1 400 m占据优势,而在较高海拔1 400~2 400 m优势已不明显,但生物量最后都下降是因为都同样受海拔变化所引起的相关环境因子改变的影响,导致最后的生物量逐渐下降。进一步研究发现,华山松地下生物量及各个器官生物量均随海拔上升,显现出逐渐下降的趋势,与地上生物量的变化较为一致。由此可以看出,随海拔上升,华山松无论是整体上还是局部上均受到了不利影响。
图4中华山松地上生物量与地下生物量的比值保持在3.1~3.2,随海拔的上升呈现出逐渐上升的趋势,这表明随海拔上升,立地条件变差,根系的生长优先于地上部分的生长,即华山松地上生物量随海拔下降较地下生物量下降趋势相对缓慢。同时华山松地上生物量与地下生物量变化趋势的一致性,体现了植株生长地上部分于地下部分相互促进、相互依赖的特性。

图3 华山松地上生物量与地下生物量随海拔梯度的变化格局Fig.3 Changes in above-ground biomass and below-ground biomass with elevation gradient

图4 华山松地上生物量与地下生物量的比值随海拔梯度的变化Fig.4 Armandi above-ground biomass and below-ground biomass ratio change with altitude
3 结论与讨论
本研究以河南省小秦岭国家级自然保护区56个20 m×20 m的样地监测数据为基础,采用异速生长方程进行各相关生物量的计算,研究了小秦岭自然保护区乔木地上总生物量与地形因子(海拔、坡度、坡向)的相关性,阐述了小秦岭样地乔木地上生物量与样地中数量最大的单个物种华山松生物量随海拔的变化格局,发现二者随海拔的变化格局虽有一定的差异性但呈现总体一致性。进而研究了华山松各器官的生物量随海拔梯度变化格局,主要得出以下结论:
(1)样地乔木地上总生物量与海拔、坡度、坡向的相关性数据分析表明,该研究区中海拔与生物量的相关性极为显著,坡度和坡位与生物量的相关性不显著。
(2)小秦岭样地乔木地上总生物量,随海拔的上升,呈现出先上升后波动性下降的趋势,且海拔1 600~2 000 m范围内具有相对较高的生物量,这表明该研究区的立地条件适宜乔木的生长,且该区受人为影响较少。
(3)华山松各器官生物量随海拔上升均呈现下降趋势,说明随着海拔的上升其他环境因子发生了变化,这对华山松所有器官均产生了不利影响,也说明了华山松各器官间生长的一致性。
(4)单一物种华山松地上生物量随海拔梯度变化呈现出先略微上升(或保持不变)后逐渐下降的趋势,与样地中所有乔木地上总生物量随海拔梯度的变化格局不完全一致,这与随海拔上升华山松龄级和分布密度(对环境的适应)的不同相关,而且由于同样受海拔变化所引起的相关环境因子改变的影响,导致最后的生物量逐渐下降。因此进行森林群落的研究时,既要考虑到整体的一般性,又要考虑到局部的特殊性,整体中包含了局部的特殊性,同时局部中又会体现整体的一般性。因此在进行该区森林规划和物种的研究及保护时,应充分兼顾这种相对关系,从合适的角度出发,制定切实可行的措施。例如该区的濒危物种秦岭冷杉主要生长于高海拔生境,虽然中等海拔具有较大的生物量贡献,但是对这种濒危种的保护也是绝对不容忽视的。
(5)对样地单个物种华山松生物量与海拔的变化格局的研究表明,华山松地上生物量、地下生物量及各个器官的生物量均随海拔上升呈现下降趋势,但地上生物量与地下生物量的比值随海拔在3.1~3.2内呈现出逐渐升高的趋势,这说明随海拔上升,华山松地上生长和地下生长保持相互依赖的关系。这与马安娜等[19]对中国草地植被地上和地下生物量的关系分析和杨昊天等[18]对荒漠草地4种灌木生物量分配特征中地上生物量与地下生物量的关系研究结果无太大差别,这为以后估算地下生物量提供了依据。
对小秦岭自然保护区的生物量随地形因子的研究,结合以往对该区森林群落空间格局及生物多样性的研究,可以更好地为保护区的规划及物种保护提供有力的理论依据。
[1] 冯宗炜, 王效科, 吴 刚. 中国森林生态系统的生物量和生产力[M]. 北京:科学出版社, 1999.
[2] 陈 骥, 曹军骥, 魏永林, 等. 青海湖鸟岛水分梯度下草地生物量分配格局初步研究[J]. 干旱地区农业研究, 2014, 32(3): 202-208.
[3] 杜 虎, 曾馥平, 王克林, 等. 中国南方3种主要人工林生物量和生产力的动态变化[J]. 生态学报, 2014, 34(10): 2712-2724.
[4] 张国斌, 刘世荣, 张东远, 等. 岷江上游亚高山林区老龄林地上生物量动态变化[J]. 生态学报, 2008, 28(7):3176-3184.
[5] 张佳蕊, 张海燕, 陆健健. 长江口淡水潮滩芦苇地上与地下部分月生物量变化比较研究[J]. 湿地科学, 2013, 11(1): 7-12.
[6] 蔺恩杰, 江 洪, 赵明水, 等. 天目山自然保护区典型森林植被乔木层生物量研究[J]. 浙江林业科技, 2013, 33(1): 21-24.
[7] 黄钦忠. 不同坡位8年生厚朴人工林生物量分配格局[J]. 江西林业科技, 2010, 6 : 16-18, 21.
[8] 孙巧玉, 刘 勇, 李国雷, 等. 坡位对油松人工林地上生物量分配格局的影响[J]. 中南林业科技大学学报, 2012,32(9): 102-105,129.
[9] 王雪峰. 海拔高度和坡向对长鞭红景天生物量的影响 [D]. 拉萨:西藏大学农牧学院, 2010.
[10]LIN D M, LAIi J S, MULLER-LANDAU H C, et al. Topographic variation in aboveground biomass in a subtropical evergreen broad-leaved forest in China[J]. PloS one, 2012, 7(10): e48244.
[11]陈 云, 王海亮, 韩军旺,等. 小秦岭森林群落数量分类排序及多样性垂直格局.生态学报[J]. 2014,34(8): 2068-2075.
[12]王海亮. 河南小秦岭自然保护区天然混交林物种多样性研究[J]. 河南科学, 2013(1):53-57.
[13]陈存根,彭 鸿. 秦岭火地塘林区主要森林类型的现存量和生产力[J]. 西北林学院学报, 1996,11(增): 92-102.
[14]李 银,侯 琳,陈军军,等. 秦岭火地塘林区典型灌木生物量估算模型[J]. 东北林业大学学报, 2014, 42(2): 116-119.
[15]黄劲松, 邸雪颖.帽儿山地区6种灌木地上生物量估算模型[J]. 东北林业大学学报, 2011, 39(5): 54-57.
[16]王天博, 陆 静. 国外生物量模型概述[J]. 中国农学通报, 2012, 28(16): 6-11.
[17]SAH J P, ROSS M S, KOPTUR S, et al. Estimating aboveground biomass of broadleaved woody plants in the understory of Florida keys pine forests[J]. Forest Ecology and Management, 2004, 203: 319-329.
[18]杨昊天, 李新荣, 刘立超, 等. 荒漠草地4种灌木生物量分配特征[J]. 中国沙漠, 2013, 33(5): 1340-1348.
[19]马安娜, 于贵瑞, 何念鹏, 等. 中国草地植被地上和地下生物量的关系分析[J]. 第四纪研究, 2014, 34(4): 769-776.
(责任编辑:朱秀英)
The arbor biomass change with terrain factor in Xiaoqinling National Nature Reserve
JIA Hongru1, ZHANG Xu2, CHEN Yun3, HAN Junwang4, YE Yongzhong2, HUANG Qunce5, YUAN Zhiliang2
( 1.Henan Finance and Taxation College, Zhengzhou 451464, China; 2.College of Life Sciences, Henan Agricultural University, Zhengzhou 450002, China; 3.College of Forestry, Henan Agricultural University, Zhengzhou 450002, China; 4.Administration Bureau of Xiaoqinling National Nature Reserve of Henan, Lingbao 472500, China; 5.Provincial Key Laboratory of Ion Beam Bio-engineering, Zhengzhou University, Zhengzhou 450052, China)
According to the investigation of 56 quadrats in Henan Xiaoqinling, this study explored the change pattern of above-ground biomass of the Xiaoqinling forest trees and studied the change pattern of Pinus armandii biomass through allometric equations. The results showed : (1) the ground biomass was statistically correlative with the altitude factor; (2)with altitude rising the tree aboveground biomass showed a downward trend after rising; (3) armandi pine is the largest species abundance whose aboveground biomass showed a trend of gradual decline with altitude ascent, which is different from the change pattern of tree biomass on the ground, and this may be caused by the disadvantage in high elevation for armandi pine; (4) with altitude rising the biomass of each organ of armandi pine falls, which is in line with armandi pine biomass changes ; (5) armandi pine aboveground biomass and underground biomass change trend is consistent, and their ratio presents a rising trend between 3.1 and 3.2 along with the elevation.
Xiaoqinling; biomass; allometric equations; altitude
2014-11-10
河南省重点科技攻关项目(132102110133);河南省教育厅科学技术研究重点项目(14A180013)
贾宏汝(1979-),女,河南驻马店人,博士研究生,主要从事生物多样性研究。
袁志良(1976-),男,河南信阳人,副教授,博士。
1000-2340(2015)06-0777-06
Q948
A
猜你喜欢
科海故事博览(2022年28期)2022-11-25 03:09:48
天天爱科学(2022年4期)2022-05-23 12:41:46
当代陕西(2022年5期)2022-04-19 12:10:28
现代园艺(2021年23期)2021-12-01 07:47:44
林业勘查设计(2020年1期)2021-01-18 02:40:48
新农业(2020年18期)2021-01-07 02:17:08
中国森林病虫(2020年1期)2020-01-18 07:21:52
新闻传播(2018年15期)2018-09-18 03:20:12
小哥白尼(野生动物)(2018年4期)2018-08-01 03:41:10
诗选刊(2015年6期)2015-10-26 09:47:16
