
The inf l uence of selective cutting of mixed Korean pine(Pinus koraiensis Sieb.et Zucc.)and broad-leaf forest on rare species distribution patterns and spatial correlation in Northeast China
2015-06-09 18:06••
Journal of Forestry Research 2015年4期
••
ORIGINAL PAPER
The inf l uence of selective cutting of mixed Korean pine(Pinus koraiensis Sieb.et Zucc.)and broad-leaf forest on rare species distribution patterns and spatial correlation in Northeast China
Binbin Kan1•Qingcheng Wang1•Wenjuan Wu1
©Northeast Forestry University and Springer-Verlag Berlin Heidelberg 2015
This study aimed to demonstrate change in spatial correlation between Korean pine(Pinus koraiensis Sieb.et Zucc.)and three rare species,and change in spatial distribution of four species in response to a range of selective cutting intensities.We sampled three plots of mixed Korean pine and broad-leaf forest in Lushuihe Forestry Bureau of Jilin province,China.Plot 1,a control, was unlogged Korean pine broad-leaf forest.In plots 2 and 3,Korean pine was selectively cut at 15 and 30%intensity,respectively,in the 1970s.Other species were rarely cut.We used point-pattern analysis to research the spatial distributions of four tree species and quantify spatial correlations between Korean pine and the other three species, Amur linden(Tilia amurensis Rupr.),Manchurian ash (Fraxinus mandshurica Rupr.),and Mongolian oak (Quercus mongolica Fisch.)in all three plots.The results of the study show that selective cutting at 15%intensity did not signif i cantly change either the species spatial patterns or the spatial correlation between Korean pine and broadleaf species.Selective cutting at 30%intensity slightly affected the growth of Korean pine and valuable species in forest communities,and the effect was considered nondestructive and recoverable.
Korean pine broad-leaf forest⋅Cutting intensity⋅Rare species⋅Spatial pattern⋅Spatial correlation
Introduction
Population spatial patterns and spatial correlations are critical to population study(Borchsenius et al.2004;Daz et al.2000).Population spatial pattern is studied to quantitatively describe stable population structures and understand their genesis(Condit et al.2000;Druckenbrod et al. 2005).Such studies contribute to understanding species growth,reproduction,death,regeneration,and resource utilization,as well as ecological processes(such as seed dispersal,intra-or inter-species competition,and interference),the biological characteristics of population(such as life history,phototaxis,and tolerance of shade)and environmental factors(e.g.,plant adaptations to environments, environmental heterogeneity)(Nathan 2006).Population distribution patterns change over time(Chen and Bradshaw 1999;Schurr et al.2004;Wiegand and Moloney 2004)due to environmental adaptation and natural selection(Masaki et al.2004).Environment often inf l uences the progress of population growth.Human activity is an important factor, especially harvesting,which affects ecosystem structure in ways different from the direction and speed of natural succession.Under the inf l uence of disturbance,different species have different mechanisms to adapt,thereby affecting the competition of plants for limited resources and resulting in differentiation of spatial patterns of populations(He and Duncan 2000).
Large areas of original forest in China were cut between the 1950s and 1980s.After 30 years of protection,natural secondary forests formed with varying patterns resultingfrom natural succession.Questions for forest managers include:which pattern is more conducive to population development and what kind of cutting intensity yields a productive pattern.Study of distribution pattern aids understanding of the structure and of forest succession processes.This provides a guide for complementary planting and harvesting during forest regeneration.Many researchers in China have focused on the study of the spatial patterns or spatial correlations of forest species(Ge et al.2008;Tang et al.2003;Yu et al.2009).However,the inf l uence of cutting intensities on spatial patterns and spatial correlation analysis of rare species in northeast China has been little studied.In this study,we researched spatial distributions of four tree species and spatial correlation between Korean pine and three other species,Amur linden,Manchurian ash,and Mongolian oak.
Materials and methods
Study area
The study was conducted at Dongsheng tree farm of Lushuihe Forestry Bureau in Jilin Province,China,west of the Changbai Mountains.Average elevation of the study site was 600–800 m,rarely above 1000 m,and the aspect mainly faced northwest.The climate is temperate continental.The combination of low temperature,low evaporation,and moist atmosphere benef i ts the formation of thick deciduous broad-leaf and coniferous mixed forests. Mixed Korean pine and broad-leaf forest was the typical vegetation type,where coniferous tree vegetation included Korean pine and Larch(Larix gmelinii(Rupr.)Kuzen.). Broad-leaf vegetation included Manchurian ash(Fraxinus mandshurica),Amur linden(Tilia amurensis),Amur phellodendron(Phellodendron amurense Rupr),Mongolian oak(Quercus mongolica),White birch(Betula platyphylla Suk),Manchurian walnut(Juglans mandshurica Maxim),Poplar(Populus spp.),Elm(U.Pumila L.)and Mono maple(Acer mono Maxim.).
Field investigation and data collection
Three sampling plots on similar aspects,slope gradients, and forest stands were demarcated in 2011 based on a longterm f i eld investigation and historical data from the local forestry bureau in Lushuihe area(Table 1).Plot 1 was original Korean pine and broad-leaf forest.Korean pine and other species had not been cut and human disturbance was uncommon.In plots 2 and 3,Korean pine was selective cut in the 1970s at 15 and 30%intensity,respectively, while other species were rarely cut.Study plot 1 was used as a control.The size of each sampling plot was 100×100 m,divided into 25 squares with dimensions of 20×20 m using a compass.A basis point was set up every 20 m.All trees of diameter at breast height(DBH)>1 cm were tagged,and species,height,crown dimensions (east–west direction=crown length,north–south= crown length)and the tree location coordinates were recorded.
Statistical analysis
We used Ripley’s K function-based approach for spatial point pattern analysis(Ripley 1978).This method is commonly used to analyze spatial patterns at arbitrary scales because results are not inf l uenced by population density(Duncan and Stewart 1991;Zhan 1998;Rebertus and Moser 1989;Wang et al.1998).Ripley’s K function assigns individual trees a two-dimensional point and analyzes the spatial pattern and spatial correlation at multiple scales based on a two-point picture that includes all individuals.The method is widely used in the analysis of spatial patterns in plant populations.Besag(1977)optimized Ripley’s K function and proposed the evolutive function-L(t)(Besag 1977).In this study,L(t)functions were used for data analysis and processed using Programit software,in which the‘r’range of spatial scale was 0–50 m,and step length was 1 m with 98%conf i dence intervals bounded by upper and lower envelopes.These functions were calculated 100 times using the Monte-Carlo approach.In analyzing patterns of spatial distribution,if the function curve was above the upper envelope,under the lower envelope,or between these two envelopes,such pattern was considered an aggregated distribution,uniform distribution,or random distribution,respectively.For spatial correlation study,function L12(t)deformed from Ripley’s function k12(t)was adopted to calculate the spatial correlation of bivariate point patterns.If the function curve was above the upper envelope,under the lower envelope,or between these two envelopes,the relationship was considered positive,negative,or neutral correlation, respectively(Xin 2009).
Results
Structural analysis of diameter class
Korean pine was the dominant species in all three plots based on its highest percentile of large diameter trees (DBH>50 cm)and largest breast-height basal area.The total breast-height basal area of broad-leaf species was signif i cantly less than that of Korean pine(Table 2;Fig.1). Because of selective cutting,the number of Korean pine trees left in plot 1 was more than that of plot 2,and plot 2was more than plot 3.The diameters of most Mongolian oak,Amur linden,and Manchurian ash ranged from 10 to 40 cm.

Table 1 Characteristics of three sample plots
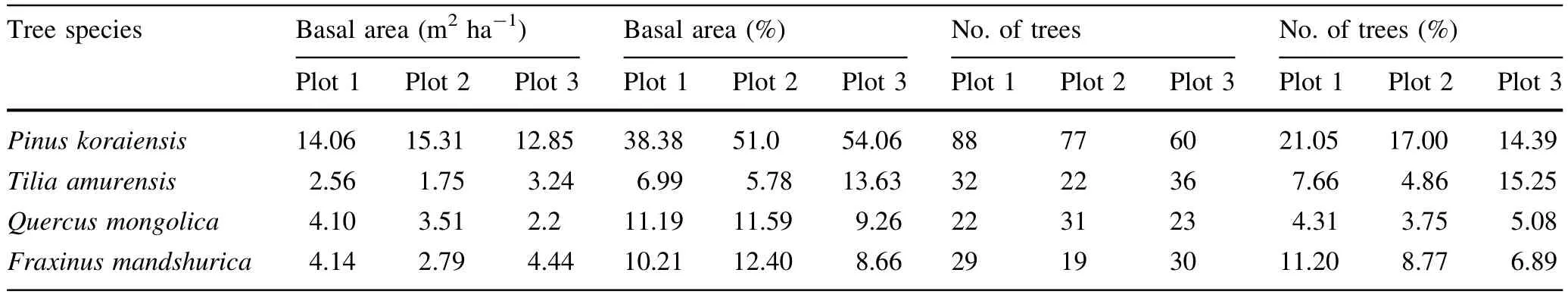
Table 2 Basal area of four tree species and number of trees in three plots
Spatial distribution of Korean pine and three associated species
The distribution of Korean pine in three plots is presented in Fig.2.In the three plots,the density of Korean pine followed the order from high to low,of plot 1,plot 2,and plot 3.Aggregated distribution of Korean pine was shown in plot 1.Selective cutting in plot 3 resulted in a lack of Korean pine in some areas,and aggregated distribution was infrequently observed.Korean pine was aggregated at 0–23 m scale and randomly distributed at 23–50 m scale in plot 1(Fig.3,a-1),Korean pine showed an overall random distribution with an aggregation trend in plot 2(Fig.3, a-2).In plot 3,the distribution of Korean pine was aggregated,random,and uniform on 0–7 m,7–25 m,and 25–50 m scales,respectively(Fig.3,a-3).Mongolian oak was randomly distributed in plots 1 and 2(Fig.3,b-1,b-2). Aggregated distribution of Mongolian oak was recorded in plot 3 at both 5–10 m and 15–37 m scales,but random distribution was recorded at other scales(Fig.3,b-3). Amur linden was randomly distributed at 20–33 m and 37–48 m scales,or aggregated on other scales in plot 1 (Fig.3,c-1),randomly distributed at 0–50 m scale in plot 2 (Fig.3,c-2),and aggregated at 0–50 m scale in plot 3 (Fig.3,c-3).Manchurian ash was randomly distributed in plots 1 and 2,but aggregated at 17–28 m scale in plot 2, (Fig.3,d-1,d-2)and aggregated at all scales in plot 3 (Fig.3,d-3).
Analysis of spatial correlation between Korean pine and rare trees
Korean pine and Mongolian oak showed overall neutral correlation in all three plots,but positive correlation at 10–20 m scale in plot 1(Fig.4,e-1)and at a 0–12 m scale in plot 3(Fig.4,e-3).Korean pine and Amur linden showed neutral correlation at 0–25 m scale or negative correlation at 25-50 m scale in plot 1(Fig.4,f-1).Neutral correlation was recorded at 0–10 m scale or a tendency to negative correlation at 10-40 m scale in plot 2(Fig.4,f-2), and positive correlation,neutral,and negative correlation at 3–18 m,18–30 m,and 30-50 m scales in plot 3(Fig.4, f-3).Korean pine and Manchurian ash were neutrally correlated in all three plots,but negatively correlated at 37–42 and 44-50 m scales in plot 1(Fig.4,g-1),negatively correlated at 5–11 m scale in plot 2(Fig.4,g-2),and negatively correlated at 11–16 m scale in plot 3(Fig.4,g-3).
Discussion
Effects of selective cutting on spatial pattern of trees
Numerous studies have demonstrated that most natural populations,including conifer species,follow a cluster pattern as a result of the combined actions of arboreal growth,survival,intra-or inter-species competition,and interaction between species and habitat,thus ref l ecting adaptive mechanism of populations(Taylor et al.1996; Manabe and Yamamoto 1997;Zheng 1997;Manabe et al. 2000).Tree harvest inf l uences resource availability andalters the resource utilization eff i ciency of plant populations,thereby affecting changes in spatial relationships of plants.In this study,Korean pine showed superiority in all three plots based on its percentile of basal area in all species,although two of the plots had been selectively harvested at different cutting intensities.More Korean pines of large diameter and fewer saplings were observed,and most broad-leaf species were located in renewal and succession layers.Korean pine was aggregated at small scale(0–23 m) or randomly distributed at large scale(>23 m)in plot 1, which could be explained by the number of saplings showing aggregated distribution around the parent trees that were randomly distributed.In previous studies of structural change in populations at different developmental stages,a similar phenomenon was observed(Li et al.2005; Xu et al.2005;Zhang et al.2009).This could be related to seed dispersal methods,renewal modes,and intra-or interspecies competition.Gravity is essential in the seed dispersal of Korean pine,in which seeds are scattered around the seed tree,resulting in aggregated distribution at small scales(Smith 1983).Otherwise,some scholars believe that the aggregate distribution of Korean pines is mainly due to action of squirrels(Lu 2003)Intra-specif i c competition for light,moisture,and nutrients causes large-scale mortality of Korean pine trees,resulting in a tendency toward random distribution of Korean pine in the population.Species tend to be aggregated at lower canopy layers,but randomly or evenly distributed in upper layers,the aggregation degree decreasing with canopy height(Zhang et al.2007). The severe selective cutting(30%intensity)caused a uniform distribution of Korean pines at broader scales as a result of human disturbance.Amur linden,Manchurian ash,and Mongolian oak all showed random distribution in plot 2,comparable to that in plot 1 and aggregated distribution in plot 3.Severe cutting produced large forest gaps that differed from the gaps formed by natural disturbance.This can inf l uence the structure and dynamic process of populations,thereby leading to the differentiation of distribution(Li and Wang 2009).Forest gaps increase ground temperatures and accelerate the decomposition of litter,this enables some light-tolerant species to accelerate growth,which increases population aggregation(Bai 2012 and Wan 2005).In plots 1 and 2,most broad-leaf species were recorded in succession and renewal layers.These species consume more resources,including light,moisture, and limited nutrients in the microenvironment.Competition results in natural forest thinning,which could explain why aggregated distribution is relatively uncommon and eventually manifests as uniform or random distribution.
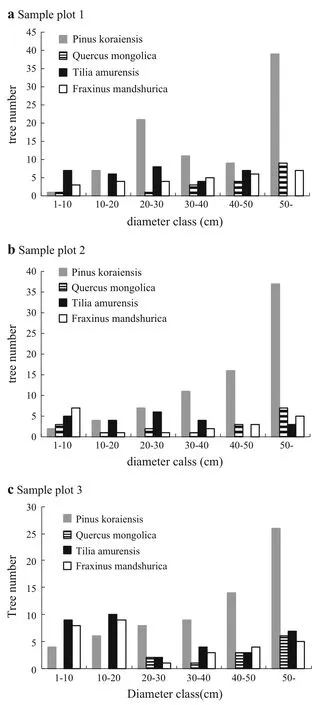
Fig.1 Diameter class of the four species,a–c is four species in plot 1–3
Selective cutting effects on spatial relationships of trees
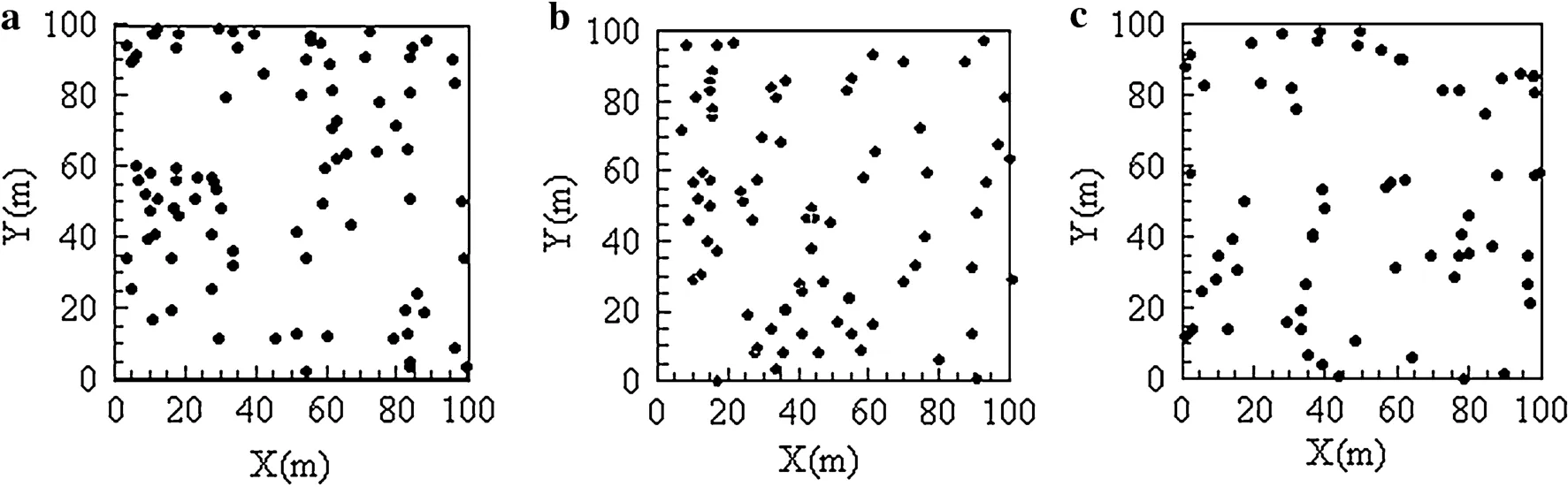
Fig.2 Spatial distribution of Pinus koraiensi s(dbh≥1 cm)in three plots(100×100 m),a–c is Pinus koraiensi s in plot 1–3

Fig.3 Spatial distribution of four species in each sample plot,X—scale/m,Y—L(t),black lines for L(t);the grey lines are the upper and lower envelopes;a1–a3 is the Pinus koraiensis spatial pattern in plots 1–3;b1–b3 is the Quercus mongolica spatial pattern in plots 1–3;c1–c3 is the Tilia amurensis spatial pattern in plot 1–3;and d1–d3 is the Fraxinus mandshurica spatial pattern in plots 1–3
Korean pine and Amur linden were negatively correlated at large spatial scale in all three plots(Fig.4,f1–f3).This suggested that Korean pine inhibited the growth of Amur linden at large spatial scale.This is consistent with results of Zhao et al.(2011).Korean pine was dominant in space and widely distributed in the overstory layer of the three plots,whereas Amur linden grew in the forest gaps,mostlylocated in the lower layer.Both Korean pine and Amur linden are light-tolerant species.(Song 2007;Xu 2001) Korean pine blocked sunlight required by Amur linden, resulting in a negative correlation or a tendency to be negatively correlated at large spatial scales.In contrast, Korean pine and Amur linden were positively correlated at small spatial scales under severe cutting because the selective cutting released the space occupied by Korean pine so that it enabled Amur linden to consume more sunlight and nutrients(Chen et al.2008).
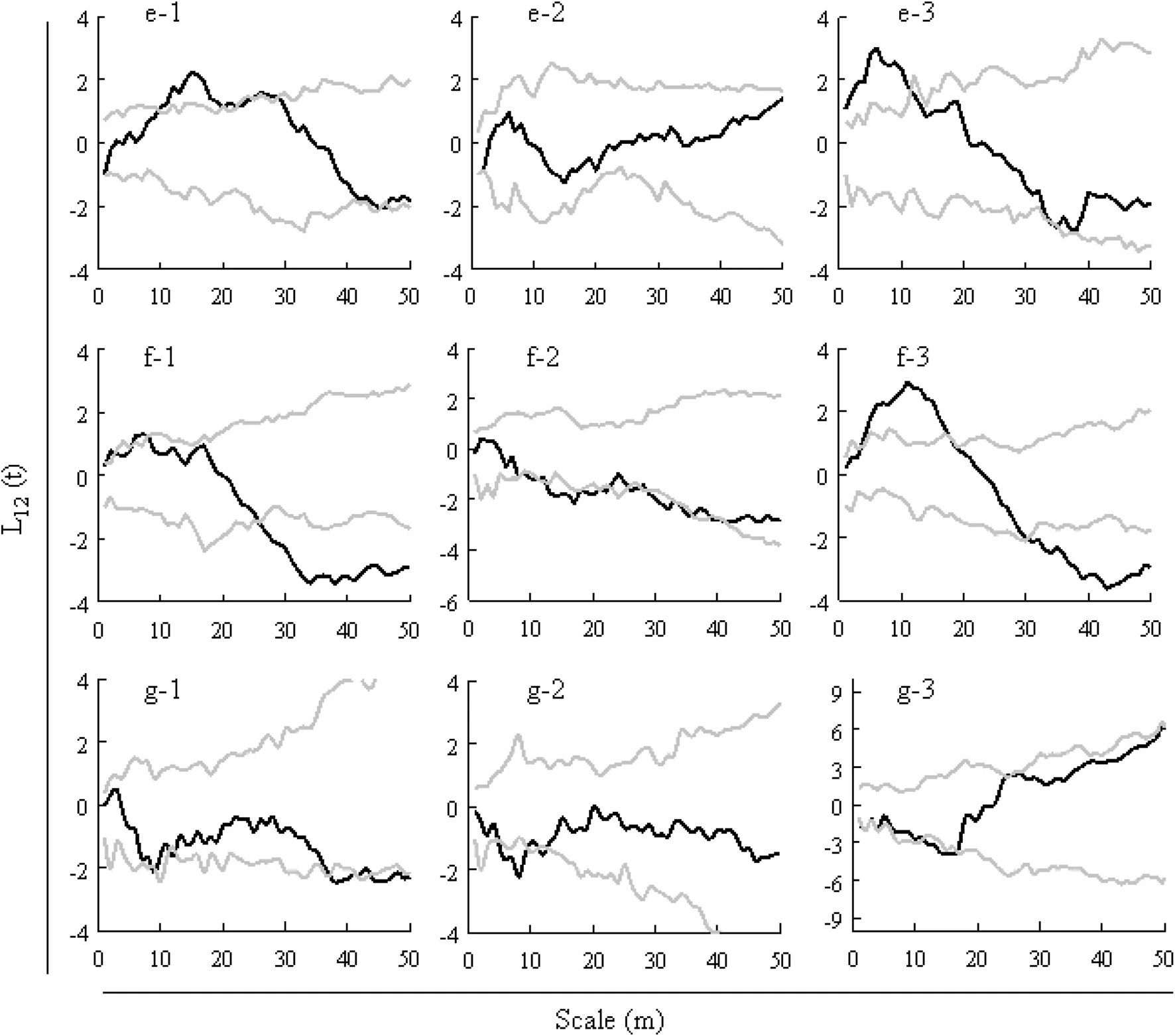
Fig.4 Spatial correlations of Pinus koraiensis and three species in each sample plot,X—scale/m,Y—L12(t),black lines for L12(t);the grey lines are the upper and lower envelopes;e1–e3 is the Pinus koraiensis and Quercus mongolica spatial correlation in plot 1–3;f1–f3 is the Pinus koraiensis and Tilia amurensis spatial correlation in plot 1–3;g1–g3 is the Pinus koraiensis and Fraxinus mandshurica spatial correlation in plot 1–3
Korean pine and Manchurian ash were neutrally correlated in the three plots,with a tendency toward positive correlation as cutting intensity increased.This f i nding indicated that these two species could coexist in one community with minimal interplay.Most Manchurian ash grew in the midstory layers,where the requirements of growth space and nutrients were slightly limited by Korean pine.However,these two species were located in different growth layers.Manchurian ash grew well in the gaps of Korean pine,Han et al.(2004)found that sunlight penetration is critical to the regeneration of Manchurian Ash (Han et al.2004).Brokaw(1987)reported that some species are restricted by the size of canopy gaps.However, spatial distribution of saplings of most species is unrelated to canopy gaps(Milton Lieberman and Rodolfo 1995; Hubbell et al.1999).So it is necessary to further study the relationship between the canopy gap size and spatial distribution of Korean pine broad-leaf forest with respect to the distribution pattern of lower species.
Korean pine and Mongolian oak were neutrally associated in three plots.But a positive correlation was recorded between them at 0–12 m scale in plot 3 where excessive partial cutting released suff i cient sunlight,moisture,and nutrients to the broad-leaf species in the midstory andunderstory layers.This result is different from that of Zhang et al.(2006),which showed a positive correlation between Korean pine and Mongolian oak at 48–50 m scale, Zhang et al.(2006)has doubts about this result,he thinks because of Korean pine and Mongolian oak from 48 to 50 m scales are randomly distributed,so although the two species showed positive spatial correlation,there may be no interaction between them.Such spatial correlation may not have been caused by the effects of interspecif i c relationships(Zhang et al.2006).
Conclusions
The control plot without cutting was observed to have stable structureatthe top levelofsuccession.Thecutting ofKorean pine atthe overstory layerbenef i ted the growth ofbroad-leaf species.Large Korean pines and few seedlings or saplings were recorded in the three sampling plots.Such a pattern is not conducive to regeneration of Korean pine populations. Therefore,appropriate selective cutting of Korean pine is a good alternative to facilitate itsregeneration and promote the growth ofothervaluable broad-leafspecies.Korean pine and Amur linden compete with each other at large scales.Thus, complementary planting of Amur linden is not suggested. Selective cutting of either species could be considered according to demand to reduce interspecif i c competition and accelerate species growth.
AcknowledgmentsThe study was funded by China National Science and Technology Support Program(Grant No.2012BAD21B02). We are grateful to Wang Shuli,Gao Qunzong,Wang Haiyang for fi eld investigation assistance,and the Lushuihe Forestry Bureau of Jilin Province of China for logistical support.We also would like to thank two anonymous reviewers for their invaluable suggestions.
Bai Y(2012)Effects of Different cutting methods on species diversity forLarix gmeliniForests.For Sci Technol 37(3):20–23
Besag J(1977)Contribution to the discussion of Dr Ripley’s paper. J R Stat Soc B39:193–195
Borchsenius F,Nielsen PK,Lawesson J(2004)Vegetation structure and diversity of an ancient temperate deciduous forest in sw denmark.Plant Ecol 175(1):121–135
Brokaw NVL(1987)Gap-phase regeneration of three pioneer tree species in a tropical forest.J Ecol 75:9–20
Chen J,Bradshaw GA(1999)Forest structure in space:a case study of an old growth spruce-f i r forest in changbaishan natural reserve. PR china.Forest Ecology and Management 120(1–3):219–233
Chen J,Mao ZJ,Ma LX(2008)Response of photosynthetic capacity and chlorophyll f l uorescence inQuercus mongolicaandTilia amurensisseedlings after light intensity transfer.Bull Bot Res 28(4):471–476
Condit R,Ashton PS,Baker P,Bunyavejchewin S,Gunatilleke S, Gunatilleke N,Hubbell SP,Foster RB,Itoh A,LaFrankie JV et al(2000)Spatial patterns in the distribution of tropical tree species.Science 288(5470):1414–1418
Daz S,Mercado C,Alvarez-Cardenas S(2000)Structure and population dynamics of pinus lagunae M.-F.Passini.For Ecol Manag 134(1):249–256
Druckenbrod DL,Shugart HH,Davies I(2005)Spatial pattern and process in forest stands within the virginia piedmont.J Veg Sci 16(1):37–48
Duncan RP,Stewart GH(1991)The temporal and spatial analysis of tree age distributions.Can J For Res 21(12):1703–1710
Ge HL,Zhou YZ,Tang MP,Ding LX(2008)A modif i ed ripley’s index.Acta Ecologica Sinica 28(4):1491–1497
Han YZ,Wang ZQ,Gu JC(2004)The effects of spatial eterogeneity of understorey light.Acta Phytoecologica Sinica 28(4):468–475
He F,Duncan RP(2000)Density-dependent effects on tree survival in an old-growth douglas f i r forest.J Ecol 88(4):676–688
Hubbell SP,Foster RB,O’Brien ST,Harms KE,Condit R,Wechsler B,Wright SJ,de Lao SL(1999)Light-gap disturbances, recruitment limitation,and tree diversity in a neotropical forest. Science 283(5401):554–557
Li F,Wang YY(2009)Population structure and distribution pattern of broussonetla papyrifera sprouting in different disturbed conditions.J Fujian Coll For 29(4):362–367
Li MH,He FH,Liu Y,Pan CD(2005)Spatial distribution pattern of tree individuals in the schrenk spruce forest northwest china. Acta Ecologica Sinica 25(5):1000–1006
Lu CH(2003)Review on the study of relationship between natural regeneration ofKorean pine and animals.Chin JEcol22(1):49–53
Manabe T,Yamamoto SI(1997)Spatial distribution of eurya japonica in an old-growth evergreen broad-leaf forest,sw japan.J Veg Sci 8(6):761–772
Manabe T,Nishimura N,Miura M,Yamamoto S(2000)Population structure and spatial patterns for trees in a temperate oldgrowth evergreen broad-leaf forest in japan.Plant Ecol 151(2): 181–197
Masaki T,OtaT Sugita H,Oohara H,Otani T,Nagaike T,Nakamura S(2004)Structure and dynamics of tree populations within unsuccessful conifer plantations near the shirakami mountains,a snowy region of japan.For Ecol Manag 194(1–3):389–401
Milton Lieberman DL,Rodolfo P(1995)Canopy closure and the distribution of tropical forest tree species at laselva,costa Rica. J Trop Ecol 11:161–178
Nathan R(2006)Long-distance dispersal of plants.Science 313(5788):786–788
Rebertus AJW,Moser EB(1989)Fire-induced changes in quercus laevis spatial pattern in f l orida sandhills.J Ecol 77(3):638–650
Ripley BD(1978)Spectral analysis and the analysis of pattern in plant communities.J Ecol 66(3):965–981
Schurr F,Bossdorf O,Milton S,Schumacher J(2004)Spatial pattern formation in semi-arid shrubland:a priori predicted versus observed pattern characteristics.Plant Ecol 173(2):271–282
Smith G(1983)Quantitative plant ecology.Blackwell Scientif i c Publications,Oxford,pp 55–56
Song XZ(2007)Research on Regeneration and microsites within logging gaps in Changbai Mountain.Chinese Aeademy of Forestry Press,Beijing,pp 4–25
Tang MP,Tang SZ,Lei XD,Zhang HR,Hong LX,Feng YM(2003) Edge correction of ripley’s k(d)function on population spatial pattern analysis.Acta Ecologica Sinica 23(8):1533–1538
Taylor A,Zisheng Q,Jie L(1996)Structure and dynamics of subalpine forests in the wang lang natural reserve,sichuan, china.Vegetatio 124(1):25–38
Wan H(2005)An impacts of logging operations on understory plants for the broadleaved/Korean pine mixed forest on Changbai Mountain.China.J For Res 16(1):27–30
Wang ZF,An SQ,Zhu XL,David G,Campell Yang XB(1998) Distribution pattern of tree populations in tropical forest and comparison of its study methods.Chin J Appl Ecol 9(6):575–580
Wiegand T,Moloney KA(2004)Rings,circles,and null-models for point pattern analysis in ecology.Oikos 104(2):209–229
Xin X(2009)Community structure and population spatial pattern of pinus tabulaeform is natural forests.J Northeast For Univ 37(3):18–20
Xu ZB,Dai LM,Chen JQ(2001)Natural regeneration condition inPinus koraiensisbroad-leaved mixed forest.Acta Ecologica Sinica 21(9):1413–1420
Xu XH,Yu MJ,Hu ZH,Li MH,Zhang FG(2005)The structure and dynamics of castanopsis eyrei population in gutian mountain natural reserve in zhejiang,east china.Acta Ecologica Sinica 25(3):645–653
Yu H,Yu XH,Ci LJ(2009)Variations of spatial pattern in f i remediated mongolian pine forest,hulun buir sand region,inner mongolia,china.Chin J Plant Ecol 33(1):71–80
Zhan JT(1998)Analysis of spatial point pattern for plant species. Acta Phytoecologica Sinica 22(4):344–349
Zhang CY,Zhao XH,Wang XY,Hou JH(2006)Spatial pattern and canopy structure of korean pine broadleaf forests in changbaishan mountains.J Beijing For Univ 12(28):45–50
Zhang JT,Hao ZQ,Li BH,Yao XL(2007)Spatial distribution patterns and associations ofPinus koraiensisandTilia amurensisin broad-leaf korean pinem ixed forest in changbaimountains. Chin J Appl Ecol 18(8):1681–1687
Zhang Y,Zhang CY,Zhao XH(2009)Spatial distribution pattern of main tree species in pinus tabulaeformis forest of heilihe in inner-mongolia.Acta Botanica Boreali-Occidentalia Sinica 9(1):0167–0173
Zhao ZH,Hu YB,Zhou HM,Zhang LJ(2011)Spatial distribution patterns and associations of dominate population in two types of broadleaf korean pine mixed forest.For Res 24(5):554–562
Zheng YR(1997)The applicability of various methods in analysis of picea mongolica population spatial distribution pattern.Acta Phytoecologica Sinica 21(5):480–484
28 June 2014/Accepted:12 January 2015/Published online:2 July 2015
Project funding:The study was funded by China National Science and Technology Support Program(Grant No.2012BAD21B02).
The online version is available at http://www.springerlink.com
Corresponding editor:Chai Ruihai
✉Qingcheng Wang
wqcnefu@163.com
1Forestry College of Northeast Forestry University,26 Hexing Rd,Harbin 150040,People’s Republic of China
 Journal of Forestry Research2015年4期
Journal of Forestry Research2015年4期
- Journal of Forestry Research的其它文章
- Drone remote sensing for forestry research and practices
- Life cycle environmental impact assessment of biochar-based bioenergy production and utilization in Northwestern Ontario, Canada
- Growth rates of Eucalyptus and other Australian native tree species derived from seven decades of growth monitoring
- Effect of f i rst thinning and pruning on the individual growth of Pinus patula tree species
- Modeling forest f i res in Mazandaran Province,Iran
- Effect of soil physical–chemical properties on the decay of standing Pinus koraiensis in Xiaoxing’an Mountains, northeast China