
青冈1年生播种苗生长节律
2015-06-09 14:20:32窦全琴
江苏林业科技 2015年1期
汤 槿,汤 榕,侯 超,陈 超,窦全琴
(1.江苏汤氏园林有限公司,江苏 句容 212400;2.江苏省林业科学研究院,江苏 南京 211153)
青冈1年生播种苗生长节律
汤 槿1,汤 榕1,侯 超1,陈 超1,窦全琴2*
(1.江苏汤氏园林有限公司,江苏 句容 212400;2.江苏省林业科学研究院,江苏 南京 211153)
依据青冈1年生苗高、地径的观测数据,采用Logistic方程对其生长过程进行拟合。结果表明:(1)青冈播种苗出苗时的高、茎生长很快,出苗后的高、径生长符合“S”型曲线,呈现“慢-快-慢”节律。(2)根据生长量变化节律,苗高、地径生长过程可分为出苗期、生长前期、速生期和生长后期;苗高速生期为55~169 d,速生点出现在第111 d;地径速生期为44~209 d,速生点时间为第125 d;苗高、地径的累积生长量均在速生期最大,分别占理论上限值的57.59%和56.17%。(3)苗高、地径生长的速生期持续时间有差异。苗高的速生期持续时间少于地径,而速生点的出现则早于地径,111 d之前苗木主要进行高生长,111 d之后则以地径生长为主。
青冈;播种苗;Logistic方程;生长节律
青冈(Cyclobalanopsis glauca)是壳斗科(Fagaceae)青冈属常绿高大乔木,在长江以南的北亚热带、中亚热带以及南亚热带的常绿阔叶林和落叶常绿阔叶混交林中均有自然分布。青冈是江苏珍贵的硬阔叶用材树种,但是长期以来对青冈的研究开发工作很少,现有的种质资源均零星分布在马尾松、杉木针叶林以及其他次生林、残次林中,正面临枯竭[1],因此,亟需开展种苗繁育技术研究,满足江苏造林绿化对青冈良种壮苗的需求。目前,针对青冈苗木培育的研究,主要集中在种子萌发的生理过程及形态结构[2-3]、不同施肥方式对苗木生长的影响等方面[4]。关于青冈苗木生长节律方面的研究,仅见易咏梅比较了光皮桦和青冈1年生苗的生长规律[5]。对于如何准确表述植物生长进程的问题,许多学者已经进行了大量研究和探讨[6-9],研究苗期生长规律是培育壮苗的基础。本文从1年生青冈苗木动态的苗高、地径生长量入手,采用Logistic方程对苗高、地径生长动态进行模拟,根据其中参数分析生长动态规律,期望为青冈壮苗培育提供科学依据。
1 材料和方法
1.1 试验地概况
试验地设在江苏省句容茅山镇丘陵岗坡地,属于北亚热带季风气候,年平均气温15.4℃,极端最高气温40.7℃,极端最低气温-14.0℃,年降雨量1 031 mm,年均相对湿度77%,无霜期227 d。土壤为下蜀系黄土母质发育的黄褐土,pH5.0~6.0,土质粘重,土壤肥力一般。
1.2 试验材料
试验用种子为2010年10月中旬采自句容茅山磨盘林场,随即进行沙藏处理。翌年3月5日播种在塑料拱棚的沙床中,3月30日开始出苗,4月12日将长有1对真叶的芽苗移入苗圃地,株行距15 cm×30 cm,6月中旬施用尿素1次,整个生长季不进行遮荫。
1.3 数据调查
生长过程采用定株、定期调查法。于5月2日待苗木生长正常开始进行第1次测量,以后每15 d左右定期测定标准株的苗高、地径,直至11月30日结束,此时苗高生长停止,苗梢已经封顶,调查期237 d,调查14次,调查数据使用Excel2007,DPS v14.10软件进行统计及分析。
1.4 模型选择与参数估计
根据青冈出苗后的苗高、地径与时间的散点图可知其生长曲线近似“S”形。本文选用Logistic方程模拟青冈苗期高、地径的年生长动态。
其方程可以表示为Y=k/(1+ea-bt) (1)
式中k,a,b 3个参数在DPS v14.10统计分析平台上运用麦考特算法进行同步估计[7-8]。此法能动态估算k,a,b的值,避免了3点法、4点法等方法估算a,b时对k值精确的依赖,且参数的残差平方和最小,方程拟合精度更高[9]。对(1)式进行二阶、三阶求导,即可了解生长曲线在时间轴上的瞬时变化情况[10-11]。
首先,对式(1)求二阶导数,且令其为0

可得生长曲线瞬时最大斜率对应的时间,即

t0为生长量日增长速度最大时间,称之为速生点(D)。再对(1)式求三阶导数,也令其为0,即可
得d3y/dx3=b3k ea-bt(e2lna-2bt-4ea-bt+1)(1+ea-bt)4=0,即e2lna-2bt-4ea-bt+1=0,可得生长曲线上瞬时斜率连续变化最快的2个时间点,当t<(a-1.317)/b为生长初期,在此阶段苗高、地径生长缓慢。
当(a-1.317)/b<t<(a+1.317)/b为速生期,在此阶段苗高、地径生长迅速。
当t>(a+1.317)/b为生长后期,在此阶段苗高、地径生长缓慢[7]。
2 结果与分析
2.1 苗高与地径生长状况
青冈子叶留土萌发,胚根向下生长,胚芽呈钩状伸出播种面,幼茎由红褐色转为暗绿色,茎生长至3.0 cm以上时1对初生叶展开。青冈出苗后首次测定的苗高达3.34 cm,占全年高生长的20.39%,此后幼苗生长非常缓慢,6月中旬前苗高净生长量为1.28 cm,占全年高生长的7.81%,6月中旬后苗高生长逐渐加快,7月2日至7月24日(22 d)苗高出现第1次生长高峰,高生长量为全年的18.10%。7月25日至8月11日苗高的生长量次之,8月12日后苗高生长进入第2次高峰,高生长量为全年的14.46%,7月2日至9月26日(86 d)苗高净生长量为10.11 cm,占全年高生长的61.72%,9月27日后苗高净生长量为1.65 cm,占全年高生长的10.07%。
青冈地径在出苗期生长较快,出苗后的生长较缓慢,6月底7月初以前(71 d)的地径生长量仅为0.062 cm,占全年地径生长量的14.35%,7月后生长加快,7月3日至10月26日(117 d)地径净生长量为0.246 cm,占全年生长的56.94%,10月27日后地径净生长量为0.052 cm,占全年生长的5.72%(见表1)。
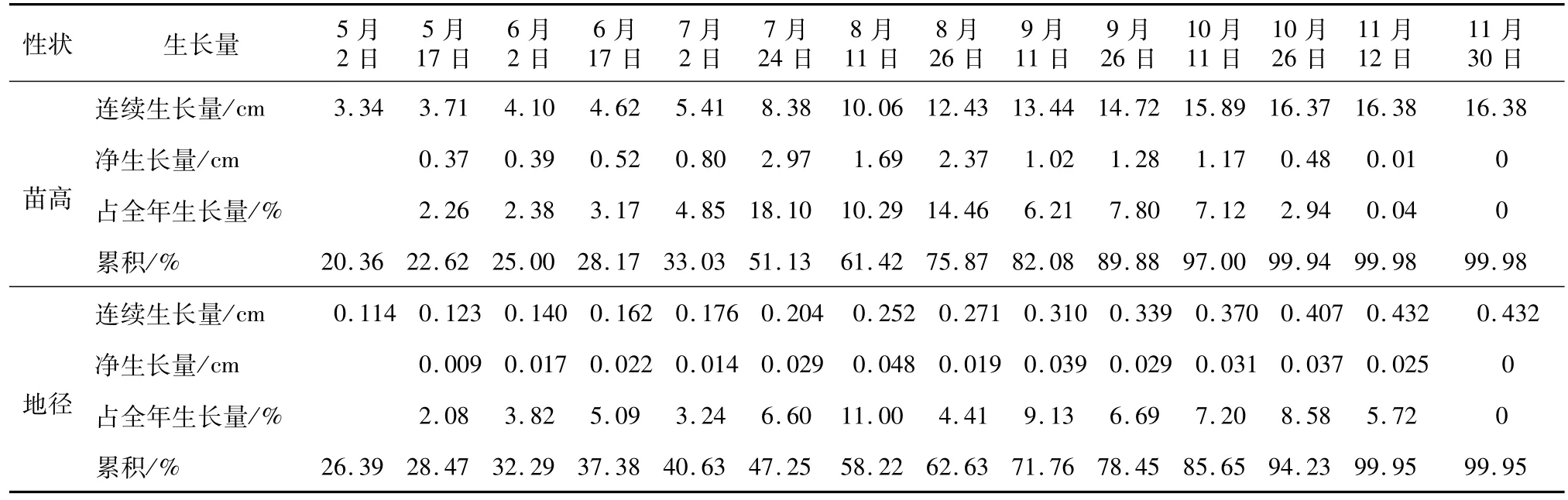
表1 青冈1年生播种苗苗高、地径生长量
2.2 苗高、地径生长动态分析
应用Logistic方程和青冈不同时期苗高、地径生长量,得到苗高生长数学模型:YH=18.109 535/(1+e(2.554698-0.022860t)(R2=0.986);地径生长数学模型:YD=0.516 318/1+e(1.97203-0.015711t)(R2= 0.992)。上述2个回归方程的决定系数均98%以上,因此可以采用此方程进行生长参数估计,青冈1年生苗出苗后的苗高生长过程符合“慢-快-慢”的S型曲线的节律,54 d前苗高生长缓,55~169 d为速生期,速生点出现在第111 d(见图1);与苗高相比,地径的速生期为44~209 d,持续时间长达166 d,速生点出现的时间晚于苗高(125 d),生长后期时间较短为28 d(见图2)。
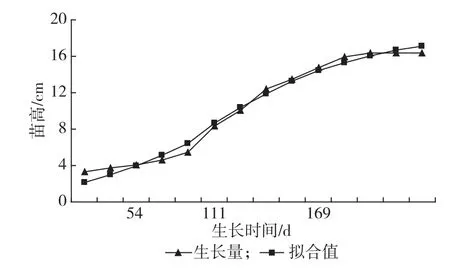
图1 青冈苗高生长进程
2.3 苗高、地径生长时期划分
青冈苗高速生期始于6月1日结束于9月24日,累积生长量达10.43 cm,占理论上限值的57.59%,速生点出现在7月27日,生长前期和生长后期的生长量较小,分别占理论上限值的21.87%和15.02%;地径的生长节律基本同于苗高,地径的速生期持续的时间较长(166 d),累积生长量达0.29 cm,占理论上限值的56.17%,生长前期的累积生长量占理论上限值略高于苗高,而生长后期持续时间很短(28 d),累积生长量仅0.06 cm,占理论上限值的11.62%(见表2)。
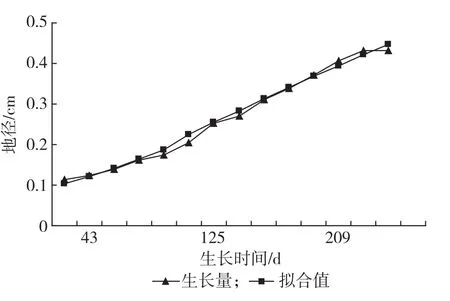
图2 青冈地径生长进程
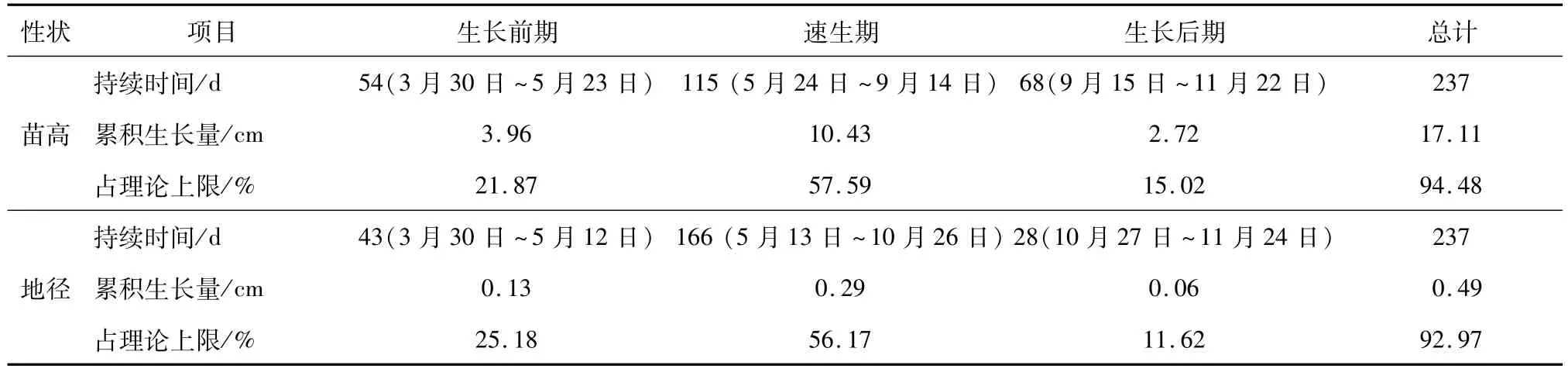
表2 青冈1年生播种苗苗高、地径生长时期
3 结论与讨论
(1)青冈1年生播种苗形态构成期的高、茎生长很快,这与青冈萌发生理过程有关,其种子富含淀粉60%以上,种子沙藏吸涨阶段也是种子内物理和化学变化的过程。曾丹娟等研究结果表明,淀粉含量在青冈种子萌发过程中从第2 d开始一直呈减少的趋势,说明淀粉在整个萌发过程中是青冈种子主要的能量来源,把贮存淀粉水解为简单有机物,并运输到胚根、胚轴、子叶,供给幼苗至形态构成等一系列生理活动,需要一段时间[2]。幼苗破土时,胚轴生长没有土壤限制,胚轴伸出1对真叶萌发时,幼茎即3.0 cm以上,出现第1个生长的“异养”高峰。但出苗后的50 d内,种子贮存的营养物质随形态构成逐渐耗尽,地上部分叶面积小,合成的光合产物较少,高、茎生长进入缓慢的间歇期;50 d后当真叶增长到充足的数量,加之根系吸收能力的增强,光合能力和生理活力也随之增强,幼苗进入快速生长阶段,又出现第2个生长“自养”高峰,苗高速生期在55~169 d,170 d后生长变缓;地径速生期在44~209 d,210 d后生长基本停滞,因此青冈出苗后其生长呈现“慢-快-慢”节律,生长进程符合Logistic曲线模型方程。
(2)由青冈生长曲线的变化节律可知,可将1年生青冈苗高、地径生长过程划分为出苗期、生长前期、速生期和生长后期,与易咏梅对青冈苗期生长阶段的划分基本一致[5],但苗木速生期出现的时间与持续时间略有差异,本试验青冈苗木速生期出现的时间稍晚,速生持续时间较易咏梅所得结果长30 d左右[5],可能与试验立地条件和苗期施肥管理等因素有关。应用Logistic方程可计算各生长阶段相应的生长量累积,速生期的苗高、地径生长量占理论上限值的比例分别为57.59%和56.17%。速生期内的生长量累积在总生长量中所占贡献最大,因此可作为制定培育青冈良种壮苗的田间水肥管理措施的理论依据。
(3)青冈自然分布于本省常绿阔叶林或常绿阔叶与落叶阔叶混交林中,是北亚热带宁镇丘陵区具有代表性的森林群落之一,幼苗喜湿耐阴,苗期生长较慢。本试验青冈1年生播种苗是在全光照圃地上培育,因此,尚需探讨采取不同遮荫措施和林下侧方庇荫条件对青冈幼苗生长发育和生理特性的影响,以期为青冈种苗培育,现有林分抚育及植被恢复提供理论依据和技术支撑。
[1] 黄宝龙.江苏森林[M].南京:江苏科学技术出版社,1998.
[2] 曾丹娟,罗艾莹,白坤栋,等.青冈种子萌发的生理生态学研究[J].种子,2013,32(10):8-11.
[3] 张德楠,罗艾莹,徐广平,等.温度和土壤含水量对青冈种子萌发的影响[J].广西植物,2013,33(3):306-312.
[4] 陈秋夏,王金旺,郑 坚,等.不同施氮水平对青冈容器苗的形态和生理特性影响[J].中国农学通报,2011,27(28):28-35.
[5] 易咏梅.光皮桦与青冈一年生播种苗生长特性比较[J].林业科技通讯,1999,(9),24-26.
[6] 王莽莽,李典谟.用麦夸方法最优拟合逻辑斯谛曲线[J].生态学报,1986,6(2):142-147.
[7] 李秋元,孟德顺.Logistic曲线的性质及其在植物生长分析中的应用[J].西北林学院学报,1993,8(3):81-86.
[8] 郑立飞,赵惠燕,刘光祖.Richards模型的推广研究[J].西北农林科技大学学报:自然科学版,2004,32(8):107-110.
[9] 程毛林.Richards模型参数估计及其模型应用[J].数学的实践与认识,2010,40(12):139-143.
[10]唐启义,冯明光.实用统计分析及其DPS数据处理系统[M].北京:科学出版社,2002.
[11]董筱昀,黄利斌.金焰彩栾嫁接苗年生长规律研究[J].江苏林业科技,2014,41(2):20-23
S792.16
A
10.3969/j.issn.1001-7380.2015.01.009
1001-7380(2015)01-0040-04
2014-09-03;
2014-12-02
江苏省林业三新工程项目“珍贵树种青冈的种苗快速繁育与推广”(LYSX[2012]51)
汤 槿(1973-),女,江苏句容人,大学本科毕业,研究方向:林木栽培与管理技术。
*通信作者:窦全琴(1965-),女,河南沈丘人,研究员,大学本科毕业,主要从事林木遗传育种工作。E-mail:douqq2008@163.com。
猜你喜欢
莫愁(2023年9期)2023-03-17 09:37:12
莫愁·家教与成才(2023年3期)2023-03-15 00:55:23
中国民间疗法(2021年13期)2021-08-30 08:56:36
农技服务(2020年1期)2020-12-17 08:15:01
山东农业科学(2019年11期)2019-12-24 01:11:27
现代农业科技(2019年1期)2019-07-11 01:36:25
现代农业科技(2017年24期)2018-01-22 21:35:06
西藏科技(2016年9期)2016-09-26 12:21:38
江苏农业科学(2016年6期)2016-07-25 00:16:35
林业与生态(2016年2期)2016-02-27 14:23:57
