
Peroxiredoxin 2基因RNAi慢病毒载体构建及对SW480细胞增殖的影响*
2015-06-01 10:42冯继红傅仲学文坤明卢伟东陈旺盛郭金宝张寿儒
重庆医学 2015年1期
冯继红,傅仲学△,文坤明,卢伟东,王 昊,陈旺盛,郭金宝,张寿儒
(1.重庆医科大学附属第一医院胃肠外科 400016;2.遵义医学院附属医院胃肠外科,贵州遵义 563003;3.泸州医学院附属医院胃肠外科,四川泸州 646000)
论著·基础研究
Peroxiredoxin 2基因RNAi慢病毒载体构建及对SW480细胞增殖的影响*
冯继红1,傅仲学1△,文坤明2,卢伟东1,王 昊1,陈旺盛3,郭金宝1,张寿儒1
(1.重庆医科大学附属第一医院胃肠外科 400016;2.遵义医学院附属医院胃肠外科,贵州遵义 563003;3.泸州医学院附属医院胃肠外科,四川泸州 646000)
目的 构建Peroxiredoxin2(PRDX2)基因RNA干扰(RNAinterference,RNAi)慢病毒表达载体,探讨PRDX2基因干扰后对结直肠癌SW480细胞增殖的影响。方法 设计、合成靶向PRDX2的RNA干扰的序列,构建pGC-EGFP-shPRDX2慢病毒载体并进行鉴定,同时应用qRT-PCR和Westernblot方法观察转染的SW480结肠癌细胞PRDX2mRNA和蛋白表达的抑制效果,并通过MTT、平板克隆形成实验检测细胞增殖变化。结果 成功构建PRDX2基因慢病毒载体并经测序证实;pGC-EGFP-shPRDX2可有效抑制结直肠癌SW480细胞PRDX2的表达,感染慢病毒的SW480细胞中PRDX2mRNA和蛋白表达水平明显降低(P<0.05);SW480细胞经PRDX2RNA干扰后其生长和增殖能力显著降低(P<0.05)。结论 PRDX2基因RNAi慢病毒表达载体在SW480细胞中表达稳定可靠,PRDX2基因干扰后有效抑制了结直肠癌SW480细胞的增殖和生长,为进一步探讨PRDX2在结直肠癌发生、发展及转移中的作用奠定基础。
结直肠肿瘤;基因表达;绿色荧光蛋白质类;过氧化还原蛋白2;慢病毒载体
结直肠癌(colorectal carcinoma,CRC)是消化系统常见恶性肿瘤之一。全球范围内,结直肠癌分别位居男、女性恶性肿瘤的第3、4位,导致每年61万人死亡[1-2]。在中国,近年结直肠癌发病率逐年上升,且发病年龄明显提前,每年约有 40 万新发病例,现在我国消化系统恶性肿瘤中列第2位[3]。
随着肿瘤的快速生长,肿瘤组织内常常会出现局限性缺氧灶,并且这些缺氧灶随着肿瘤内血管的生长而变化。低氧导致了活性氧(reactive oxygen species,ROS)的产生,细胞保护系统可破坏ROS并修复其造成的损害。过氧化还原蛋白2(peroxircdoxin 2,PRDX2)是一种新近发现的相对分子质量在20~30×103的过氧化物酶家族(peroxircdoxins,PRDXs)成员之一,可通过其过氧化物酶等功能或以分子伴随物形式在肿瘤细胞对抗ROS、放化疗抵抗、细胞增殖、分化、凋亡及肿瘤血管形成等过程中发挥重要作用。前期实验结果证实PRDX2基因在结直肠癌中表达上调,且与结直肠癌的发生、发展有密切关系[4]。本实验通过构建PRDX2基因的慢病毒表达载体,制备高滴度慢病毒颗粒并感染结直肠癌SW480细胞株,检测PRDX2在mRNA 和蛋白质水平的表达情况,同时观察感染慢病毒的SW480细胞株其生长和增殖情况,为进一步探讨PRDX2基因对结直肠癌SW480细胞生物学行为的影响及其分子作用机制奠定基础。
1 材料与方法
1.1 材料
1.1.1 细胞与试剂 人结直肠癌SW480细胞株购自中国生命科学院上海细胞库,HpaI和XhoI酶切、增强剂Eni.S、polybrene购自上海吉凯基因化学技术有限公司,Lipofectamine 2000购于Invitrogen公司,QIAGEN Plasmid大抽Kit购自QIAGEN,T4 DNA ligase购自NEB公司,Taq DNA polymerase、dNTP Mix、DNA Ladder 购自Fermentas 公司,PrimeSTARTM HS DNA Polymerase购自Takara公司,胎牛血清(FBS)、牛血清清蛋白(BSA)购自上海微科生化试剂有限公司,DMEM、Leibovitz′s L-15培养基购自GIBCO公司,蛋白裂解液 RIPA 和蛋白酶抑制剂 PMSF 均购自于上海生物试剂厂,胰酶购自上海化学试剂公司,BCA蛋白含量试剂盒购于南京凯基生物公司,PRDX2一抗购自美国Abcam公司,GAPDH、GFP 抗体和HRP标记鼠二抗均购于Santa Cruz 公司。
1.1.2 RNAi重组慢病毒载体 据GeneBankPRDX2基因转录本(NM_005809.4)设计5个针对PRDX2基因的siRNA序列和阴性对照(negative control)siRNA,构建的目的慢病毒载体命名为pGC-EGFP-shPRDX2,由上海吉凯基因化学技术有限公司构建和包装。
1.2 方法
1.2.1 PCR扩增及产物鉴定 针对目的基因序列,据RNA 干扰序列设计原则,设计多个 RNA 干扰靶点序列,选择最佳的动力学参数靶点进入后续实验流程;同时设计RNAi 阴性对照Scramble 序列(5′-TTC TCC GAA CGT GTC ACG T-3′)。针对靶点设计5条shRNA,分别命名为:PRDX2-RNAi(8818-2)、PRDX2-RNAi(8819-1)、PRDX2-RNAi(8820-1)、PRDX2-RNAi(8821-1)、PRDX2-RNAi(8822-1),见表1。PCR 反应条件为:94 ℃ 30 s;94 ℃ 30 s、55 ℃ 50 s、72 ℃ 30 s、72 ℃ 6 min,30个循环;72 ℃延伸10 min。PCR 产物进行纯化回收后,经双酶切后的pGC-EGFP 载体连接产生shRNA慢病毒载体;对酶切片段进行回收、连接,然后转化大肠杆菌感受态细胞。挑取单菌落鉴定,引物是:上游5′- TTC TAC CCT CTG GAC TTC ACT-3′;下游 5′- GCA ATG CCC TCA TCT GTT- 3′,产物大小:251 bp。反应条件:94 ℃ 30 s;94 ℃ 30 s、60 ℃ 30 s、72 ℃ 30 s,30个循环;72 ℃延伸 10 min。设立阴性对照组(空载质粒为模板)和阳性对照组(插入GAPDH片段的载体组),对假阳性和假阴性结果进行排除。对PCR 阳性克隆片段测序,并在GenBank数据库中将测序出的结果进行Blast比对。
1.2.2 慢病毒的包装与病毒滴度测定 293 T细胞用含100 mL/L FBS的 DMEM 培养基,于37 ℃、50 mL/L CO2孵箱中培养。5个重组慢病毒质粒和空慢病毒载体分别与辅助包装载体质粒,用LipofectamineTM2000 共转染293T 细胞,具体方法参考试剂说明书进行操作。48 h后在荧光显微镜下观察到细胞绿色荧光的产生,收集上清液,20 000 r/min 超速离心3 h并浓缩病毒。最后包装产生的慢病毒有两种,即含有PRDX2基因序列的慢病毒pGC-EGFP-shPRDX2和空载对照慢病毒 pGC-EGFP-NC。逐孔稀释滴度测定法和PCR 测定其病毒滴度(TU/mL)。获得PRDX2 siRNA慢病毒溶液(5个干扰序列慢病毒液分别以设计的PRDX2干扰序列编码命名)及空载对照慢病毒pGC-EGFP-NC,病毒液分装后于-70 ℃保存。
1.2.3 病毒感染SW480细胞并检测其干扰效果 将SW480细胞计数并均匀平铺于6孔板,使其感染时细胞汇合率达60%左右,按照种植的SW480细胞数及MIO值(MIO=50)计算所加的病毒量,病毒原液用增强剂Eni.S稀释,使其浓度为标准浓度(1×108),依次加入终浓度为8 μg/mL Polybrene、Eni.S、L-15培养基及计算的标准浓度病毒液,总体积为1 mL。将含有PRDX2基因片段的慢病毒pGC-EGFP-shPRDX2与pGC-EGFP-NC分别感染SW480细胞。感染后的SW480细胞根据感染的病毒命名分组为pGC-EGFP-shPRDX组、pGC-EGFP-NC组,同时将未感染的SW480细胞作为空白对照组。各组细胞常规培养 24 h后更换为含有3 μg/mL嘌呤霉素(Puromycin)的L-15培养液进行抗性筛选,感染至3 d后行荧光显微镜下观察绿色荧光,在药物选择压力下培养4 d后收获细胞进行检测。
1.2.4 Western blot 法检测 将pGC-EGFP-shPRDX2和pGC-EGFP-NC成功转染至SW480细胞,经荧光显微镜观察,转染效率达95%以上。细胞分组如下:将GAPDH作为阳性对照组、空载对照慢病毒(pGC-EGFP-NC)以及由5个针对PRDX基因的序列的RNAi慢病毒载体质粒细胞组。收集各组细胞,PBS洗涤2次,离心弃上清液,加入100 μL预冷的RIPA细胞裂解液,置冰上30 min,4 ℃、12 000 g 离心15 min,取上清液并进行蛋白定量(BCA)法。取30 μg蛋白加入上样缓冲液,经100 ℃、10 min 变性后用100 g/L的十二烷基-聚丙烯酰胺凝胶电泳(SDS-PAGE)分离。蛋白半干转移至PVDF膜,用含50 g/L脱脂奶粉的TBST缓冲液室温封闭1 h,加入小鼠抗人PRDX2的一抗或小鼠抗人GADPH的一抗,4 ℃过夜,TBST溶液再次洗涤3次,加入HRP标记的山羊抗小鼠二抗37 ℃孵育1 h,TBST洗涤3次,化学发光(eleetroehemiluminescence,ECL)观察,将胶片拍照后用凝胶图像处理系统分析,实验重复3次。
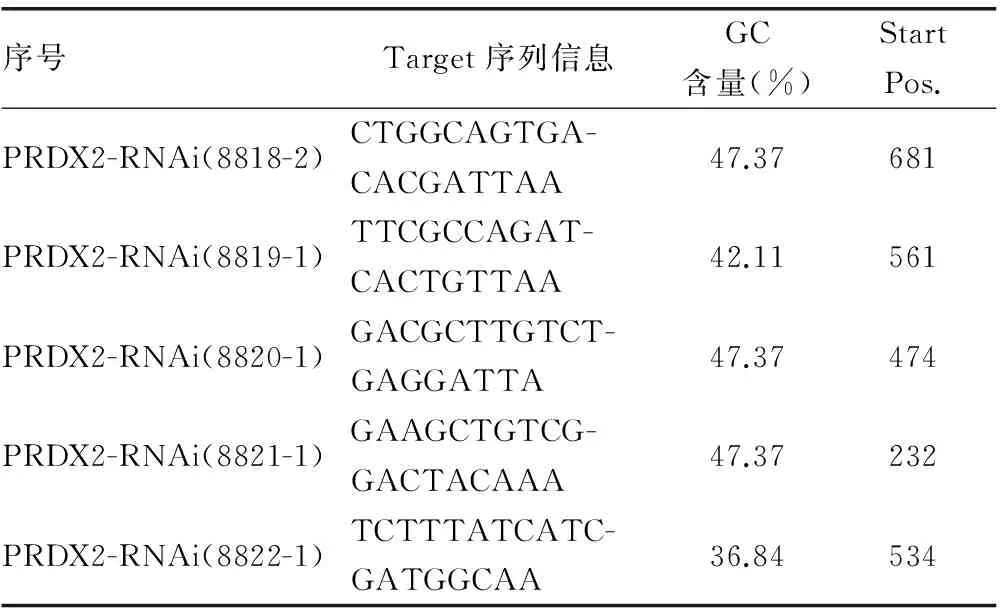
表1 化学合成的目的基因PRDX2的5条干扰序列
1.2.5 实时荧光定量PCR(qRT-PCR)检测 采用qRT-PCR技术检测慢病毒转染后的SW480细胞中PRDX2 mRNA水平表达。Trizol 抽提细胞RNA,采用Oligo(dT)引物和随机引物进行逆转录,以各组细胞cDNA作为模板,每个组设立3个复孔进行qRT-PCR检测。GAPDH上游引物为:5′-TGA CTT CAA CAG CGA CAC CCA-3′,下游引物为:5′-CAC CCT GTT GCT GTA GCC AAA -3′,引物序列长度225 bp;PRDX2基因上、下游引物为5′- CGG ACT ACA AAG GGA AGT ACG TG-3′、下游引物为5′- CCA GGT GGG TGA ACT GAG AGT C -3′,引物序列长度162 bp。PCR 的反应条件:95 ℃ 2 min;95 ℃ 20 s;60 ℃ 30 s;72 ℃ 30 s(40 个循环),反应完成后,得出Ct值,2-ΔΔCT法计算出相对表达量,分析各组之间PRDX2基因的mRNA相对表达的差异。
1.2.6 四唑盐(MTT)法测定细胞相对抑制率 取状态良好,处于对数生长期各组细胞,0.25%胰酶消化后制成单细胞悬液,调整浓度为1×104个/mL,均匀接种于96孔板内,每孔加200 μL L-15培养基;阴性对照组与空白对照组,每组设6个复孔,常规培养,每2天换1次液,连测7 d;每天固定时间向每孔加新鲜配制的5 g/L的MTT 10 μL,37 ℃孵育4 h;轻洗去上清液后每孔加DMSO 150 μL,水平震荡混匀,酶标仪于490 nm波长处测吸光度(A)值,取6孔平均值,绘制生长曲线。
1.2.7 平板克隆形成实验 取pGC-EGFP-NC组及pGC-EGFP-shPRDX2组的SW480细胞消化后平铺于6孔板中,常规培养,2~3 d更换1次新鲜培养基,待细胞生长出肉眼观察可见有明显克隆群落时取出结晶紫染色,拍照并计算细胞克隆生长群落。

2 结 果
2.1 慢病毒表达载体pGC-EGFP-shPRDX2的构建与酶切鉴定 以PRDX2基因全长克隆为模板,扩增出PRDX2基因编码框的序列,用PCR 产物进行琼脂糖凝胶电泳,PRDX2基因产物纯化后与载体pGC-EGFP行酶切、连接、转化,并得到数十个克隆,挑取5个克隆,摇菌后进行菌液鉴定,结果表明可扩增出约340 bp的目的片段(图1)。经测序鉴定与GenBank NM_005809.4序列一致,重组载体pGC-EGFP-shPRDX2构建成功。
1:ddH2O;2:pGC-EGFP;3:marker;4~8:pGC-1,2,3,4,5。
图1 阳性克隆 PCR 鉴定
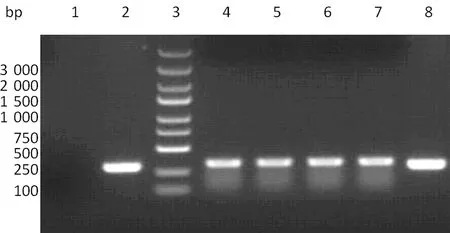
2.2 慢病毒的包装与病毒滴度的测定 慢病毒载体共转染293T细胞,于转染48 h后在荧光显微镜下观察其293T细胞发出的绿色荧光(图2)。经测定,慢病毒pGC-EGFP-shPRDX2浓缩后的病毒滴度大约为2×109TU/mL,pGC-EGFP-NC浓缩后的病毒滴度则为2×108TU/mL。
A:pGC-EGFP-shPRDX2;B:pGC-EGFP-NC。
图2 293 T 细胞病毒感染后荧光显微镜观察(×200)
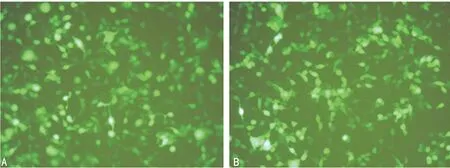
2.3 慢病毒转染SW480细胞后荧光表达检测 SW480细胞感染pGC-EGFP-shPRDX2 72 h后在荧光显微镜下观察其绿色荧光表达,从明视野和荧光视野二者比较中,可见其感染效率较高,可达95%以上,见图3。

A:明视野;B:荧光视野。
图3 转染SW480细胞72 h荧光表达检测(×200)
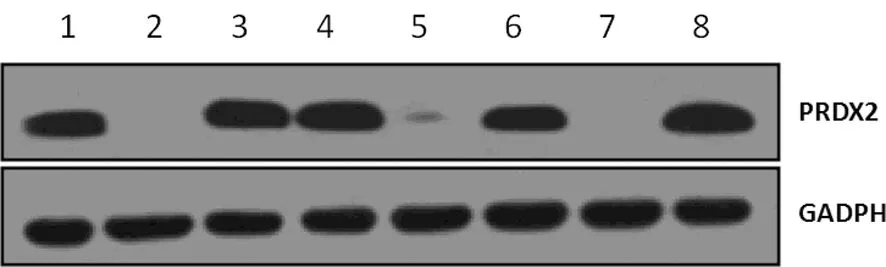
1:阳性对照组;2:空白细胞组;3:pGC-EGFP-NC组;4~8:含有针对PRDX2基因的不同干扰序列的RNAi慢病毒载体质粒细胞组(5、7表示有明显敲减效果)。
图4 Western blot 法检测感染后SW480细胞中PRDX2的蛋白水平

*:P<0.05,与pGC-EGFP-NC组比较。
图5PRDX2基因沉默在SW480细胞中的PRDX2 mRNA表达
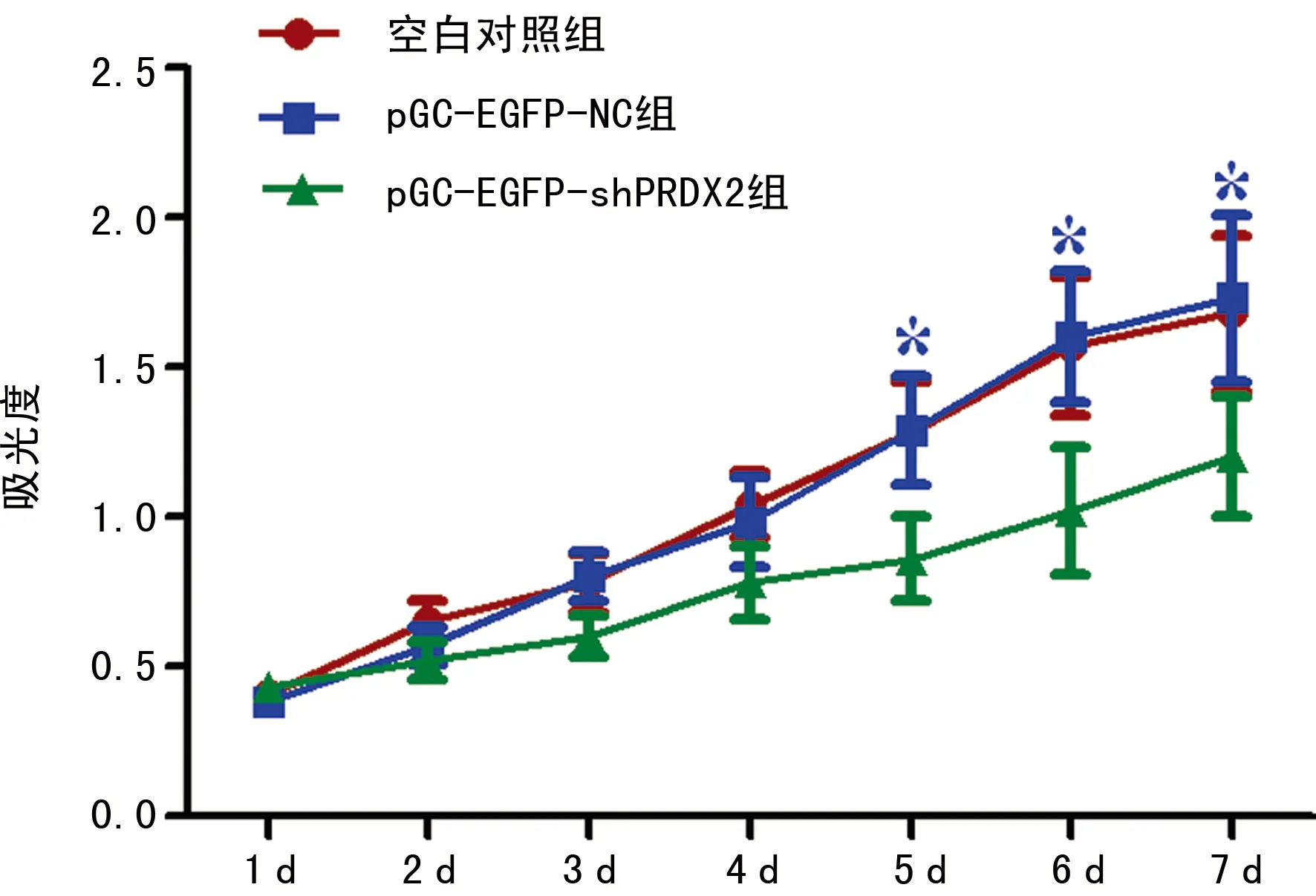
*:P<0.05,与空白对照组及pGC-EGFP-NC组比较。
图6 MTT检测PRDX2 RNAi对SW480细胞增殖抑制作用
2.4 Western blot 和qRT-PCR检测分析 检测转染不同序列 pGC-EGFP-shPRDX2慢病毒的SW480细胞中PRDX2蛋白表达水平,与pGC-EGFP-NC组相比,RNAi慢病毒载体质粒细胞组中5和7泳道表示对SW480细胞中PRDX2表达有明显敲减作用,其中7泳道PRDX2-RNAi(8822-1)的敲减效果最强,达到几乎完全抑制效果(图4)。qRT-PCR检测PRDX2基因的mRNA相对表达水平,与pGC-EGFP-NC组比较,pGC-EGFP-shPRDX2组的PRDX2基因表达明显降低,差异有统计学意义(P<0.05),可说明在mRNA水平基因敲减有效,见图5。
2.5 MTT法检测 MTT法检测各组SW480细胞6 dOD值,绘制生长曲线,结果显示,与pGC-EGFP-NC组及空白对照组相比,SW480 pGC-EGFP-shPRDX2组细胞增殖能力受到显著抑制,差异具有统计学意义(P<0.05),见图6。
2.6 平板克隆形成实验 细胞平板克隆形成实验结果(图7)显示,与pGC-EGFP-NC组及空白对照组比较,pGC-EGFP-shPRDX2组的平板克隆形成能力明显降低(P<0.05)。

*:P<0.05,与空白对照组及pGC-EGFP-NC组比较。
图7 pGC-EGFP-shPRDX2对SW480细胞平板克隆形成能力的影响
3 讨 论
生命有机体在不断进化和演变过程中,逐渐形成能够有效清除ROS产生的自由基等有害物质的细胞保护性防御系统,过氧化物酶类、超氧化物歧化酶、过氧化氢酶以及谷胱甘肽共同组成了细胞防御系统。PRDXs家族在细胞内发挥重要的抗氧化保护性功能,通过过氧化物酶活性,可减少或清除过氧化氢、过氧亚硝酸盐和一系列有机氢过氧化物(ROOH),该家族包括6个成员,广泛存在于真核及原核生物中。PRDX2属于PRDXs家族2-Cys PRDXs(PRDX 1~5)成员,是机体氧化代谢的重要酶之一,主要分布于细胞质[5]。PRDX2最主要的生物学功能是抗氧化,同时可通过调节细胞内H2O2浓度,参与H2O2介导的信号转导途径[6]。PRDX2以硫氧还蛋白为电子供体,清除活性氧族,与生物的生长发育、细胞衰老、免疫抑制密切相关,并在维持红细胞生存,促进NK细胞发挥自然杀伤功能的过程中扮演重要角色[7-8]。
近年来,随着研究的不断深入,PRDX2与肿瘤的关联机制逐渐被揭示,PRDX2不仅通过发挥抗氧化、分子伴侣、信号调节等功能维持细胞内的稳态,还通过多种信号转导途径参与肿瘤的发生、发展、转移、血管生成以及放化疗抵抗[9-13]。有研究报道,PRDX2可抑制DNA损伤诱导的肿瘤细胞死亡,更重要的是,这种保护性作用仅限于在肿瘤细胞内,且并非是通过过氧化物酶活性发挥作用的[14]。既往研究表明,在乳腺癌、肝癌、骨肉瘤及前列腺癌中PRDX2高表达,被认为是促癌因子,但在急性粒性白血病[15]、膀胱癌、黑色素瘤中表达降低,又被认为是抑癌因子。那么PRDX2在肿瘤细胞中究竟发挥什么角色?是发挥抗癌作用还是抑癌作用?这些争议仍有待探索。至今,PRDX2在结直肠癌中的作用尚未有明确报道,本研究前期实验研究也初步证实PRDX2在结直肠癌组织及细胞株中表达上调,且与结直肠癌的发生、发展、转移和预后有关。根据本实验结果,观察到转染pGC-EGFP-shPRDX2建立的稳定SW480细胞株其形态学发生了变化,不仅增殖和生长能力减弱,抗菌及抗凋亡能力也明显降低,但同时也发现细胞黏附能力和转移能力反而增强,这些生物学功能的变化虽仍需进一步验证,但亦可为下一步的研究提供一种思路,PRDX2在结直肠癌中是否可能发挥更为关键的作用,是否存在双向调控作用尚需证实,其具体分子调控机制也有待深入探讨。
本实验设计的针对PRDX2的特异性RNA干扰序列,将干扰序列定向克隆转接并感染目的细胞。为了验证PRDX2基因的siRNA重组体构建成功,本研究采用PCR鉴定和DNA测序鉴定构建的PRDX2基因siRNA慢病毒载体,证实了插入片段的序列正确,重组体构建成功。将5种构建的干扰重组体转染293T细胞,包装产生了高滴度的病毒颗粒,通过直接感染目的细胞,可有效避免感染目的细胞株出现的脱靶现象。实验得到的PRDX2 RNAi SW480细胞株,通过初步的功能学实验,证实了PRDX2基因敲减后可有效抑制结直肠癌SW480细胞的增殖和生长,这些初步研究结果为探讨PRDX2基因敲减后SW480细胞的成瘤特性、细胞周期变化及对抗肿瘤药物的敏感性等方面提供研究支撑,同时也为探讨PRDX2基因在结直肠癌中的具体调控作用及分子作用机制奠定研究基础。
[1]Siegel R,Naishadham D,Jemal A.Cancer Statistics[J].CA Cancer J Clin,2012,62(1):10-29.
[2]American Cancer Society.Colorectal cancer facts & figures[M].Atlanta:ACS,2011-2013.
[3]王锡山.结直肠癌综合治疗的规范化实施[J].中国实用外科杂志,2010,30(4):260-264.
[4]Wu XY,Fu ZX,Wang XH.Peroxiredoxins in colorectal neoplasms[J].Histol Histopathol,2010,25(10):1297-1303.
[5]Wood ZA,Schröder E,Robin Harris J,et al.Structure,mechanism and regulation of peroxiredoxins[J].Trends Biochem Sci,2003,28(1):32-40.
[6]Rhee SG,Kang SW,Jeong W,et al.Intracellular messenger function of hydrogen peroxide and its regulation by peroxiredoxins[J].Curr Opin Cell Biol,2005,17(2):183-189.
[7]Ishii T,Warabi E,Yanagawa T.Novel roles of peroxiredoxins in inflammation,cancer and innate immunity[J].J Clin Biochem Nutr,2012,50(2):91-105.
[8]Godoy JR,Funke M,Ackermann W,et al.Redox atlas of the mouse.Immunohistochemical detection of glutaredoxin-,peroxiredoxin-,and thioredoxin-family proteins in various tissues of the laboratory mouse[J].Biochim Biophys Acta,2011,1810(1):2-92.
[9]Han YH,Kwon JH,Yu DY,et al.Inhibitory effect of peroxiredoxin Ⅱ(Prx Ⅱ) on Ras-ERK-NFkappaB pathway in mouse embryonic fibroblast(MEF) senescence[J].Free Radic Res,2006,40(11):1182-1189.
[10]KangDH,LeeDJ,LeeKW,etal.PeroxiredoxinⅡisanessentialantioxidantenzymethatpreventstheoxidativeinactivationofVEGFreceptor-2invascularendothelial cells[J].Molecular Cell,2011,44(4):545-558.
[11]Choi MH,Lee IK,Kim GW,et al.Regulation of PDGF signalling and vascular remodelling by peroxiredoxin Ⅱ[J].Nature,2005,435(7040):347-353.
[12]Zhang B,Wang Y, Su Y.Peroxiredoxins,a novel target in cancer radiotherapy[J].Cancer Lett,2009,286(2):154-160.
[13]Shiota M,Yokomizo A,Kashiwagi E,et al.Peroxiredoxin 2 in the nucleus and cytoplasm distinctly regulates androgen receptor activity in prostate cancer cells[J].Free Radic Biol Med,2011,51(1):78-87.
[14]Lee KW,Lee DJ,Lee JY,et al.Peroxiredoxin Ⅱ restrains DNA damage-induced death in cancer cells by positively regulating JNK-dependent DNA repair[J].J Biol Chem,2011,286(10):8394-8404.
[15]Agrawal-Singh S,Isken F,Agelopoulos K,et al.Genome-wide analysis of histone H3 acetylation patterns in AML identifies PRDX2 as an epigenetically silenced tumor suppressor gene[J].Blood,2012,119(10):2346-2357.
Construction of lentiviral vector of peroxiredoxin 2 gene mediating RNAi and its effects on the proliferation of SW480 cell*
FengJihong1,FuZhongxue1△,WenKunming2,LuWeidong1,WangHao1,ChenWangsheng3,GuoJinbao1,ZhangShouru1
(1.DepartmentofGastroenterologicalSurgery,theFirstAffiliatedHospitalofChongqingMedicalUniversity,Chongqing400016,China;2.DepartmentofgastroenterologicalSurgery,theAffiliatedHospitalofZunyiMedicalCollege,Zunyi,Guizhou563003,China;3.DepartmentofGastroenterologicalSurgery,theAffiliatedHospitalofLuzhouMedicalCollege,Luzhou,Scihuan646000,China)
Objective To construct a lentiviral expression vector ofperoxiredoxin2(PRDX2) RNA interference (RNAi) and to investigate the effect of siRNA ofPRDX2 genes on the proliferation of human colonrectal cancer SW480 cell.Methods RNAi target sequences were designed and synthesized towards thePRDX2 gene sequences.The lentiviral vector pGC-EGFP-shPRDX2 was constructed and identified.The vector was transformed into SW480 cells,and the transfection efficiency was evaluated by fluorescence microscopy.The expression of PRDX2 was detected with Quantitative real-time PCR (qRT-PCR) and Western blot in the transfected cells.Cell growth and colony forming ability were detected with MTT and plate cloning technique.ResultsPRDX2 gene lentiviral vector was successfully established and was proved by gene sequencing.The expression of PRDX2 in mRNA and protein was significantly reduced(P<0.05).The PRDX2 mRNA and protein expression in SW480 transfected with lentiviral were significantly reduced (P<0.05),and the ability of growth and proliferation were significantly reduced(P<0.05).ConclusionPRDX2 gene lentiviral vector could be a stable and reliable tool.The proliferation and growth of SW480 cells transfected by pGC-EGFP-shPRDX2 could be effectively suppressed,which could facilitate further investigation of the roles ofPRDX2 gene in the development and progression of colorectal cancer.
colorectal neoplasms;gene expression;green fluorescent proteins;peroxiredoxin 2(PRDX2);lentivirus vector
10.3969/j.issn.1671-8348.2015.01.005
国家自然科学基金资助项目(81172295)。 作者简介:冯继红(1977-),副教授,博士,主要从事肿瘤干细胞和肿瘤免疫研究,现单位变更为遵义医学院附属医院肿瘤医院。△
,Tel:(023)67706399;E-mail:fzx990521@sina.com。
R
A
1671-8348(2015)01-0014-04
2014-09-12
2014-10-14)
猜你喜欢
环球时报(2022-09-20)2022-09-20
华人时刊(2022年9期)2022-09-06
今日农业(2020年24期)2020-12-15
华人时刊(2020年15期)2020-12-14
中国医药指南(2017年3期)2017-11-13
西南军医(2016年3期)2016-01-23
中国继续医学教育(2015年6期)2016-01-07
小资CHIC!ELEGANCE(2015年15期)2015-09-01
中国当代医药(2015年30期)2015-03-01
现代检验医学杂志(2015年4期)2015-02-06
