
不同苗龄剑麻(Agave sisalana)生理指标的相关性分析及抗旱性评价
2015-05-30 03:27:18王春雪纪中华潘志贤岳学文李纪潮闫帮国陈涛
热带作物学报 2015年7期
王春雪 纪中华 潘志贤 岳学文 李纪潮 闫帮国 陈涛
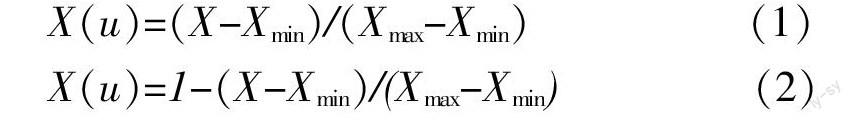

摘 要 在元谋金沙江干热河谷区进行盆栽控水试验,模拟干旱胁迫对剑麻H.11648不同苗龄(18个月、12个月、6个月)植株的生理及形态指标的影响。结果表明: (1)大苗在干旱胁迫下,随着干旱胁迫的加剧,叶片含水量随之降低,叶绿素也随之遭到破坏,叶片含水量越低叶绿素破坏越严重。对于叶片MDA含量的变化,Chl a/b及叶片含水量都是这一变化的敏感生理指标。(2)中苗在干旱胁迫下,SOD与POD两种酶协同作用来共同抵制干旱对剑麻的侵害,使其受伤害程度降到最低。(3)小苗在干旱胁迫下,酶系统中POD与SOD两种酶呈协同关系,随着丙二醛含量的增加,剑麻小苗的叶绿素、含水量、生长状况等各方面都受到显著的影响,这对小苗的破坏是全方位的。(4)根据模糊隶属函数法计算出各苗龄阶段剑麻H.11648的抗旱隶属度进行抗旱性比较,得出中苗(苗龄12个月)的综合抗旱能力最强。在对剑麻H.11648的引种过程中,12个月苗龄的剑麻在抗旱生理方面最适合栽种,它的抗旱生理特征能保证其在干热河谷最大程度的存活。
关键词 剑麻H.11648;不同苗龄;生理指标;抗旱评价
中图分类号 S563.8 文献标识码 A
Abstract Pot water control experiments were performed in Jinsha River Dry-hot Valleys in Yuanmou. The experiments simulated drought stress on different age of Agave sisalana H.11648 seedlings (18 months, 12 months, 6 months), and studied on the physiological and morphological indices. The results showed that: (1)under drought stress, with the drought stress aggravated, the leaf water content of large seedling decreased, and the chlorophyll was destroyed, chlorophyll a/b and leaf water content were all sensitive physiological indicators to blade malondialdehyde content. (2)Under drought stress, SOD and POD enzyme of medium seedling were synergy to jointly resist drought, to make its level of damage to a minimum. (3)Under drought stress, the two kinds of enzymes POD and SOD of little seedlings were in collaborative relationships. Along with increase of the content of malondialdehyde, chlorophyll content, water content, and the growth of small seedling was significantly affected, the damage to small seedling was comprehensive. (4)According to the fuzzy membership function, the membership degree of drought resistance of each age of seedling A. sisalana H.11648 was calculated, which was concluded that the medium seedling(12 months)had the strongest comprehensive drought resistance capacity. In the process of the introduction of A. sisalana H.11648, the 12 months seedling was the most suitable plant for its drought-resistant physiological. Its drought resistance and physiological features could ensure it lived to a great extent in the dry-hot valleys.
Key words Agave sisalana H.11648;Different age of Agave sisalana seedlings;Physiological indees;Drought tolerance evaluation
doi 10.3969/j.issn.1000-2561.2015.07.012
剑麻(Agave sisalana)是元谋干热河谷引进的优质产纤维植物,具有产量高、抗旱等特点,而干热河谷的水热矛盾却是限制其生长的关键问题。剑麻对干旱胁迫的适应性是其能否在干热河谷环境下生存的重要条件。不同生长期的植物对干旱的抗性是不相同的,植物个体能否忍耐干旱胁迫,主要取决于其生理、形态以及生长特性的表现[1]。植物在极端环境下对水分胁迫的生理反应和适应机制是植物水分关系研究中的重要课题之一[2]。前人对不同植物的抗旱性都进行了大量的研究分析,总结出了形态、生理、分子等不同层次下的抗旱指标,为植物抗旱性研究提供了理论支持。植物对土壤水分胁迫的响应包含着极其复杂的生理生化变化,并形成了受遗传性制约的适应机制[3-6]。因此,本研究针对同一品种不同苗龄的剑麻H.11648在元谋干热河谷干旱胁迫下抗旱生理指标进行研究,通过模拟自然干旱的控制实验来揭示不同苗期剑麻的抗旱能力,探索其适应机制,同时进行了各抗旱指标的相关性分析及抗旱性评价,为选择引种不同苗龄的剑麻提供了理论依据。
1 材料与方法
1.1 材料
1.1.1 试验材料 供试材料为剑麻品种H.11648不同苗龄的幼苗,分为大、中、小苗,苗龄分别为18、12和6个月,3种苗龄的剑麻均引种于广西。供试土壤为砖红壤,其中全氮(以N计)为0.069%,全磷(以P计)为0.033%,全钾(以K计)为0.557%,土壤容重为1.68 g/cm3。
1.1.2 试验地概况 盆栽实验于云南省农业科学院热区生态农业研究所试验大棚内进行,试验地位于金沙江元谋干热河谷区,该区地处101°35′~102°06′E, 25°23′~26°06′N, 海拔1 000~1 400 m之间,年均温21.5 ℃, 最热月(7月)均温为27.1 ℃, 最冷月(1月)均温为14.9 ℃,年均降水量<700 mm, 其中6~10月降雨量占年降水量的90%以上, 年均蒸发量>3 000 mm。
1.2 方法
1.2.1 试验组处理 试验于2012年7月到2012年11月进行,历时4个月。试验分6个小组,其中每个苗龄剑麻分别设立干旱胁迫组和正常给水组,每个试验小组设置十个剑麻盆栽,供试花盆高23 cm,口径为34 cm,装干土15 kg,采用自然状态下的砖红壤,无施肥。干旱胁迫组剑麻每30 d浇一次水,浇水量为1 L,土壤含水量保持在4%~6%;正常给水组剑麻每周浇一次水,水量为1 L,土壤含水量保持在30%~35%。各组剑麻的生理指标和形态指标每月测定一次,连续测定4个月,胁迫组和对照组均在每次浇水前一天早上测定。生理指标每组随机选取3个植株进行测定,形态指标每组随机选取5个植株进行测定,测量叶片均选取最新伸展且没有病害的成熟叶片。
1.2.2 形态及生理指标的测定 测定的形态指标有:株高、叶长、叶宽、叶厚、叶数,生理指标有:叶绿素(Chl)含量、叶片丙二醛(MDA)含量、叶片脯氨酸(Pro)含量、叶片过氧化物酶(POD)活性、叶片超氧化物歧化酶(SOD)活力、叶片含水量,测定方法参照陈建勋文献中的方法[7]。
1.3 数据处理与分析
数据采用Excel和Spss软件进行分析,采用Excel作图。用Pearson相关系数分析抗旱指标的相关关系,其中,极显著性检验水平α=0.01进行一元线性回归。
各苗龄剑麻综合抗旱能力,采用模糊隶属函数法[8-9]计算抗旱隶属度进行比较。该方法根据模糊数学原理,利用隶属函数进行综合评价。先求出各生理指标在各剑麻苗龄中的隶属函数数值,然后对各苗龄隶属函数值进行累加,求其平均值,得出综合评价指标值。相关计算公式为:
X(u)=(X-Xmin)/(Xmax-Xmin) (1)
X(u)=1-(X-Xmin)/(Xmax-Xmin) (2)
式(1)、(2)中,X(u)为某苗龄剑麻某指标的抗旱隶属函数,X为该苗龄该指标的测定值,Xmax和Xmin为各苗龄中指标最大和最小测定值。指标与抗旱性成正相关则使用公式(1),若指标与抗旱性成负相关,则使用公式(2)。将每个苗龄剑麻各指标的抗旱隶属数值累加起来,求其平均数,隶属函数均值越大,就证明该苗龄剑麻的抗旱性越强。
2 结果与分析
2.1 不同苗期剑麻生理与形态指标的相关性分析
剑麻由多种生理生化过程综合作用来适应干旱胁迫,对不同的生理及形态指标进行相关性分析,并将其中呈显著相关(p<0.01)的指标进行拟合分析,结果如下。
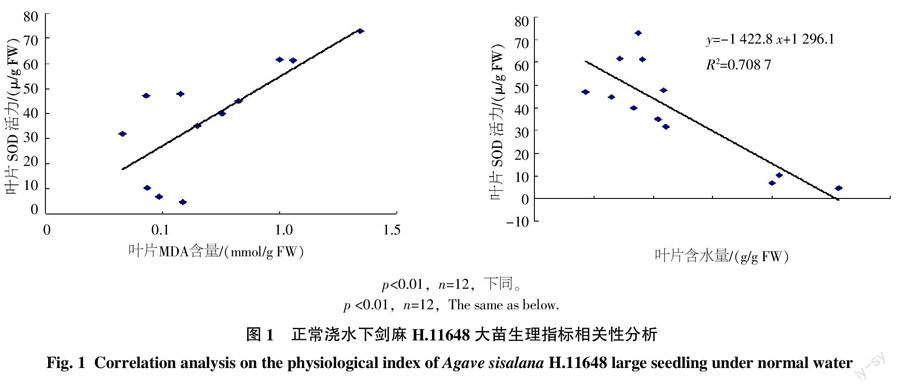
2.1.1 剑麻H.11648大苗各指标的相关性分析 如图1所示,剑麻H.11648大苗在正常浇水的情况下,叶片SOD活力与MDA含量呈显著正相关,叶片SOD活力与含水量呈显著负相关。
剑麻H.11648大苗在正常浇水情况下,SOD活力与MDA含量呈显著正相关,这说明H.11648大苗在水分胁迫前,MDA毒害的同时就启动了SOD的保护机制,这有利于植物在遇到干旱胁迫时能很快地进行自身的保护。其SOD酶活力与叶片含水量呈显著负相关,这说明大苗在正常浇水情况下,SOD酶保护系统与叶片含水量也是息息相关的,叶片含水量在正常水分环境下降低,SOD活力则相应升高。叶片SOD活力随着MDA含量以及含水量的变化趋势都说明了剑麻H.11648在正常水分环境下保护酶系统就已经启动了。
如图2所示,剑麻H.11648大苗在干旱胁迫条件下,叶片含水量与Chl a/b呈显著正相关,叶片Chl a/b与MDA含量呈显著负相关,叶片含水量与MDA含量呈显著负相关。
剑麻H.11648大苗在干旱胁迫的情况下Chl a/b与叶片含水量呈显著正相关,说明其随着干旱胁迫的加剧,叶片含水量随之降低,Chl也随之遭到破坏,叶片含水量越低Chl破坏越严重。
干旱胁迫下能够维持较高的组织含水量是植物抗旱性的一个重要特征。剑麻H.11648大苗在干旱胁迫下叶片MDA含量与Chl a/b呈显著负相关,这说明在干旱胁迫下对于叶片MDA含量的变化,Chl a/b及叶片含水量都是这一变化的敏感生理指标,这说明干旱胁迫下大苗产生的丙二醛可以同时破坏叶片的光合作用和水分调节。
2.1.2 剑麻H.11648中苗各指标的相关性分析 如图3所示,剑麻H.11648中苗在正常浇水情况下,叶厚与叶片POD活性呈显著正相关,叶片含水量与SOD活力呈显著负相关。
叶厚是剑麻的重要形态指标之一,它可以指示剑麻的缺水程度。在正常浇水情况下,中苗叶厚与叶片POD活性呈显著正相关,这说明在正常浇水情况下,随着叶厚的增加,叶片POD活性也相应地升高,叶片越成熟POD活性就越高。SOD活力与叶片含水量呈显著负相关,说明在无干旱胁迫情况下,叶片含水量越低,叶片SOD活力越高,这也说明了在干旱胁迫没有发生之前SOD保护机制已经启动。POD和SOD的活性与叶片水分参数显著相关也说明中苗的酶系统在环境还没有干旱胁迫之前就随着植物水分变化而随时保护叶片,而叶片越成熟,保护酶越完善。
如图4所示,剑麻H.11648中苗在干旱胁迫下,叶片Chl a/b与POD活性呈显著负相关,叶片Chl b含量与SOD活力呈显著正相关,叶片POD活性与叶片SOD活力呈显著正相关。
对于活性氧假说,由于Chl a对活性氧的反应较Chl b敏感,活性氧直接引发Chl的破坏及部分特异性地破坏Chl a[10-11],致使Chl含量下降及Chl a/b比值降低[12]。图4表明在干旱胁迫下Chl的破坏程度越高,中苗的POD、SOD的活性越高。POD与SOD的活性呈显著正相关,说明在干旱胁迫下两种酶协同作用来共同抵御干旱对剑麻的侵害,使其受伤害程度降到最低。
2.1.3 剑麻H.11648小苗各指标的相关性分析 如图5所示,剑麻H.11648小苗在正常浇水情况下,叶片含水量与叶数呈显著正相关。
叶数和叶片含水量都是植物的重要形态指标,剑麻H.11648小苗在正常浇水情况下生理指标间没有显著的相关关系,这说明小苗在水分适宜的情况下,抗旱机制还没有形成,但是叶数和叶片含水量呈显著正相关,这说明叶片含水量越高剑麻的叶数越多,植株的生长越快。
如图6所示,剑麻H.11648小苗在干旱胁迫下,叶片Pro含量与叶宽呈显著负相关,Chl a/b与脯氨酸含量呈显著负相关,叶片SOD活力与POD活性呈显著正相关。
叶宽是剑麻表征含水量重要的形态指标,在干旱胁迫下Pro含量与叶宽呈显著负相关,这说明渗透调节中重要的调节物质脯氨酸随着叶宽的减小而升高。Chl a/b与Pro含量呈显著负相关,说明干旱胁迫对剑麻小苗叶绿素破坏的同时,渗透调节就开始起作用。SOD与POD活性呈显著正相关,说明干旱胁迫下小苗的酶系统呈协同关系,POD与SOD共同抵御干旱对小苗的侵害。
叶片MDA含量与Chl a/b、叶片含水量、叶数呈显著负相关,说明随着MDA含量的增加,剑麻小苗的Chl、含水量、生长状况等各方面都受到显著的影响,这对小苗的破坏是全方位的。
2.2 各苗龄剑麻H.11648抗旱能力动态变化综合评价
剑麻抗旱指标的隶属函数值及其排序(表1)表明,苗龄为18个月的剑麻H.11648大苗的抗旱能力整体表现为9月份>11月份>8月份>10月份,苗龄为12个月的剑麻H.11648中苗的抗旱能力表现为11月>10月>9月>8月,苗龄为6个月的剑麻H.11648小苗的抗旱能力表现为9月>10月>11月>8月。
3 讨论与结论
POD、SOD是植物体内保护酶系统的重要组成成分,对于清除逆境下产生的有害自由基具有重要作用[13]。已有的研究表明: 干旱胁迫下植物抗旱能力和受伤害程度与保护酶的活性变化密切相关[13]。对于抗氧化酶系统,大苗在干旱胁迫发生之前,保护酶系统就已经启动,这一阶段可以称之为被动抗胁迫阶段。与大苗相比,中苗的保护酶系统在干旱胁迫之前也已经启动,并且叶片越成熟,保护酶系统越完善。在干旱胁迫真正到来时,中苗的保护酶系统与植物的受破坏程度密切相关。小苗在正常水分情况下,保护酶系统还没有启动,在干旱胁迫下,保护酶系统POD和SOD协同抵御对苗体的侵害,这说明小苗的被动抗胁迫性还没有形成,其直接进入主动适应阶段[14]。张娜等[15]研究表明,抗旱性强的燕麦品种在水分胁迫下,保护性酶被激活的速度加快,保护性酶活性上升的幅度较大,这与本研究的大苗和中苗的结果一致。
MDA是膜质过氧化的主要产物之一,它可与细胞膜上的蛋白质、酶等结合、交联使之失活,从而破坏生物膜的结构与功能,它是检测膜脂过氧化程度的一个重要指标[16]。MDA含量高低在一定程度上反映植物细胞膜脂过氧化水平和膜结构受伤害程度及植株自我修复的能力[17]。对于MDA的含量,大苗在干旱胁迫之前,叶片中少量的MDA已经启动了SOD的保护机制,在干旱胁迫下,MDA又使Chl降低,破坏叶片的光合能力。小苗在干旱胁迫下,随着叶片MDA含量的增加,Chl、含水量、生长状况等方面的危害相应增加。吴芹等[18]研究发现山杏、沙棘、油松3个树种SOD 在抵御轻、中度干旱时发挥着重要作用,干旱胁迫加剧时 POD 活性作用更大,这与本研究的大苗SOD先启动相一致。
周江等[19]研究表明,一般情况下,抗旱性强的植物具有较高的相对含水量,从而有效保护叶绿体结构以维持有效的光合作用,因此,叶绿素含量和相对含水量与植物抗旱性关联度最大。本研究中大苗在干旱胁迫下,叶片含水量降低,Chl被破坏,光合受到影响。中苗在干旱胁迫时Chl被破坏,酶系统会启动保护机制。小苗在干旱胁迫下,Chl随MDA增加而破坏。
根据张卫星[20]的抗旱性5级划分:隶属度≥0.7 为强抗,定为Ⅰ级;≥0.6 为抗,定为Ⅱ级;≥0.4 为中抗, 定为Ⅲ级;≥0.3为弱抗, 定为Ⅳ级;<0.3 为不抗, 定为Ⅴ级。因此,3种苗龄期的剑麻H.11648的隶属度均在(0.4,0.6)范围内,属于中抗。由隶属函数值可以看出,剑麻H.11648中苗表现出规律性的抗旱能力,其在干旱胁迫的4个月内抗旱能力不断增强,且在3种苗龄中的隶属函数值最高,这说明中苗较之于大苗和小苗具有更高的抗旱能力,适合于干热河谷的引种需求。根据模糊隶属函数法计算出各苗龄阶段剑麻H.11648的抗旱隶属度进行抗旱性比较,得出中苗(苗龄12个月)的综合抗旱能力最强。这可能是由于中苗较小苗已经形成了成熟的抗旱机制,且较之于大苗又在各种抗旱机能的最高效期。
在对剑麻H.11648生理及形态抗旱指标进行综合分析的前提下,发现小苗由于还没有形成特有的抗旱机制,导致其在干旱胁迫条件下酶系统、叶绿素、含水量、生长状况等各个方面都受到不同程度的破坏。而大苗在进行的抗旱综合评价中也不如中苗得分高,所以可以认为在对剑麻H.11648的引种过程中,12个月苗龄的剑麻在抗旱生理方面最适合栽种,它的抗旱生理特征能保证其在干热河谷最大程度的存活。
参考文献
[1] 龚吉蕊, 黄永梅, 葛之葳,等. 4种杂交杨对土壤水分变化的生态学响应[J]. 植物生态学报, 2009, 33(2): 387-396.
[2] 刘刚才, 纪中华, 方海东, 等. 干热河谷退化生态系统典型恢复模式的生态响应与评价[M]. 北京: 科学出版社, 2011.
[3] Egert M, Tevini M. Influence of drought on some physiological parameters symptomatic for oxidative stress in leaves of chives(Allium schoenoprasum)[J]. Environmental and experimental botany, 2002, 48: 43-49.
[4] Fornca M G C, Thi A T P, Pimentel C, et a1. Differences in growth and water relations among Phaseol us vulgaris cultivars in response to induced drought stress[J]. Environmental and experimental botany, 2000, 43:227-237.
[5] Jiang Y, Huang B. Drought and heat stress injury to two cool season turfgrasses in relation to antioxidant metabolism and lipid peroxidation[J]. Crop Science, 2001, 41: 436-442.
[6] Media B V. Regulation of gene expression during water deficit stress[J]. Plant Grow Regul, 1999, 29: 23-33.
[7] 陈建勋,王晓峰. 植物生理学实验指导[M]. 广州: 华南理工大学出版社, 2002.
[8] 黎燕琼, 刘兴良, 郑绍伟, 等. 岷江上游干旱河谷四种灌木的抗旱生理动态变化[J]. 生态学报, 2007, 27(3): 870-878
[9] 王 移, 卫 伟, 杨兴中, 等. 黄土丘陵沟壑区典型植物耐旱生理及抗旱性评价[J]. 生态与农村环境学报, 2011, 24(4): 56-61.
[10] Ingunn M V, Roar M, Eli R.Diurnal temperature ahernations(DIF/drop)affect chlorophyll content and chlorophyll a/chlorophyll b ratio in Melissa officinalis L and Ocimum basilicum L but not in Viola x wittrockiana Gams[J]. Science Horticult,2003, 97: l53-l62.
[11] 许长成, 邹 琦, 程炳嵩. 干旱条件下大豆叶片H2O2代谢变化及其同抗旱性的关系[J]. 植物生理学报, 1993, l9: 216-220.
[12] Ruth H and Harald P. Folding in vitro of light-harvesting chlorophyll a/b protein is coupled with pigment binding[J]. Journal of Molecular Biology,2002, 318: 547-556.
[13]王玉魁, 阎艳霞, 余新晓, 等. 干旱胁迫下灰白滨藜保护酶活性及抗旱生理特性研究[J]. 干旱区资源与环境, 2010, 24(10): 122-126.
[14] 安玉艳, 梁宗锁. 植物应对干旱胁迫的阶段性策略[J]. 应用生态学报, 2012, 23(10): 2 907-2 915.
[15] 张 娜, 赵宝平, 郭若龙,等. 水分胁迫对不同抗旱性燕麦品种生理特性的影响[J]. 麦类作物学报, 2012, 32(1): 150-156. [16] 林植芳, 李双顺, 林桂珠,等. 丙二醛对菠菜叶片中光合羧化酶和细胞保护酶活性的影响[J]. 植物学报, 1989, 31(11): 860-866.
[17] 王 剑, 周志春, 饶龙兵, 等. 不同磷效率马褂木种源对磷胁迫的生理反应[J]. 林业科学研究, 2006, 19(4): 527-531.
[18] 吴 芹, 张光灿, 裴 斌, 等. 3个树种对不同程度土壤干旱的生理生化响应[J]. 生态学报, 2013, 33(12): 3 648-3 656.
[19] 周 江, 裴宗平, 胡佳佳,等. 干旱胁迫下3种岩石边坡生态修复植物的抗旱性[J]. 干旱区研究, 2012, 29(3): 440-444.
[20] 张卫星, 赵 致, 柏光晓, 等. 不同基因型玉米自交系的抗旱性研究与评价[J]. 玉米科学, 2007, 15(5): 6-11.
猜你喜欢
幼儿智力世界(2024年10期)2024-12-31 00:00:00
林业勘察设计(2021年3期)2022-01-04 03:29:06
中国果业信息(2021年9期)2021-12-01 10:17:17
纺织科技进展(2021年5期)2021-07-22 08:41:42
农业科技通讯(2021年4期)2021-05-23 14:02:34
小读者之友(2020年3期)2020-04-10 11:08:15
现代园艺(2018年3期)2018-02-10 05:18:12
星星·散文诗(2017年22期)2017-12-30 05:43:59
西南农业学报(2017年5期)2017-06-23 08:14:29
山东林业科技(2016年5期)2016-07-05 00:43:00
