
不同色膜遮光对棉花功能叶光合效能的影响
2015-04-29 00:44:03张友昌张教海王孝纲夏松波宋成攀别墅
棉花科学 2015年1期
关键词:光合作用
张友昌 张教海 王孝纲 夏松波 宋成攀 别墅
摘要:试验选取长势一致的、在盛蕾期的棉花功能叶作为测定对象,通过三种不同色膜遮光处理,测定叶绿素含量、硝酸还原酶活力和光合效能,以探讨何种色膜有利于功能叶的光合作用。结果表明:功能叶的叶绿素含量以黄色透光膜遮光下最大,高于太阳光(对照);功能叶的硝酸还原酶活性在开始时以黄色透光膜遮光的处理较强,大于蓝色和红色,随着处理时间的延长,以红色透光膜遮光处理较强,且大于黄色透光膜和蓝色透光膜处理,但三者远小于对照处理;从胞间CO2浓度、外部光强、蒸腾作用、净光合作用这几个指标来看,黄色透光膜遮光处理的光合效能较好,仅小于太阳光(对照),所以在人工光源选择上,黄光是比红光和蓝光更适合植物进行光合作用的光源。
关键词:光处理;光合作用;硝酸还原酶
中图分类号:S562.01. 文献标志码: A 文章编号:2095-3143(2015)01-0012-06
DOI:10.3969/j.issn.2095-3143.2015.01.003
0 引言
光合作用是绿色植物将来自太阳的能量转化为化学能(糖)的过程。绿色植物在光合作用中捕获光能,并将其转变为碳水化合物(存储化学能)。光合作用是一系列复杂的代谢反应的总和,是生物界赖以生存的基础。光是影响光合作用最重要的因素,植物利用的光能主要来自太阳光,太阳光不是单一的光,绿色植物主要利用太阳光谱中的红光和蓝紫光,因为叶绿素a主要吸收红光,叶绿素b、胡萝卜素与叶黄素主要吸收蓝紫光。硝酸还原酶(Nitrate Reductase,简称NR)活力对光极为敏感,国内外研究证明光是诱导硝酸还原酶活性的重要因子,光敏色素参与NR的诱导[1-2]。在光形态建成或光控发育过程中,硝酸还原酶的活性是受不同光质影响的,如蓝光、红光、远红光都可诱导小麦幼苗硝酸还原酶的活性。黄化幼苗经红光照射后立即照射远红光,可逆转红光对硝酸还原酶活性的诱导效应[3]。不同色光对植物硝酸还原酶诱导能力不一样,从而对光合效能也不同,在余让才,等[4]的研究中,蓝光诱导NR 活性的能力最强, 红光最弱, 白光介于这2种光质之间,红光处理材料的NR活性只有蓝光处理的50% 左右;用 DCMU 处理后, 可以部分抑制蓝光和红光对硝酸还原酶的诱导, 其中对蓝光诱导酶活性的抑制作用更强,光对NR的诱导可能是多途径协同作用的结果。许德威,等[5]研究表明,棉花叶片用一层纱布、两层纱布和黑纸遮光处理一天后,叶片中NR活性分别为不遮光的74%、32.2%和0.2%,在荫蔽条件下叶片NR活力也远低于正常光照的。
目前,国内对光与NR的诱导研究很多[3-6],但是对在单色光处理下植物的光合作用和硝酸还原酶活性的变化研究的较少。人工光源一般都是单色光,本研究采用不同色的透光膜遮住生长发育期棉花的功能叶来测定叶片的光合作用、叶绿素含量及硝酸还原酶活力的变化,以期探明何种光有利于光合作用的进行和植物的生长发育,为设施栽培选膜及农业领域应对日益频繁的雾霾气候时采取何种人工光源来减少农作物损失提供参考。
1材料与方法
1.1试验基本情况
试验于2012年在湖北省农业科学院试验基地进行。试验田地势平坦,土壤肥力较均匀。试验于4月24日营养钵播种育苗,5月19日移栽。
1.2试验材料
试验供试棉花品种为标准棉Tm-1。光学膜由广州成烨节能科技有限公司提供,其红色透光膜的透光率为20%、阻隔红外线率为25%、阻隔黄外线率为99%,蓝色透光膜的透光率为28%、阻隔红外线率为25%、阻隔黄外线率为99%,黄色透光膜的透光率为35%、阻隔红外线率为22%、阻隔黄外线率为99%。硝酸还原酶(NR)测试盒由南京建成生物工程研究所提供。
1.3 试验设计
试验于7月24日进行,设4个处理,3个重复。处理1为红色(Red)透光膜遮蔽,处理2为蓝色(Blue)透光膜遮蔽,处理3为黄色(Yellow)透光膜遮蔽处理,处理4为无遮光处理,即太阳光(对照)。棉花田间管理按照一般规程进行,要求田间每个处理施肥浇水均衡一致。在时间上作了两个梯度,一是从早晨5:00开始连续遮光处理处理13个小时,二是从早晨5:00开始连续遮光处理处理26个小时。
1.4 调查与测定
1.4.1 农艺性状调查 在进行试验前1天开展棉花农艺性状的调查。内容包括株高、主茎真叶数等,观察试验用的棉株在生育指标上的一致性。
1.4.2 功能叶叶绿素含量测定 采用SPAD-502 叶绿素仪来通过测量叶片在两种波长光学浓度差方式650 nm 和940 nm来确定叶片当前叶绿素的相对数量。
1.4.3 功能叶硝酸还原酶活力测定 采用硝酸还原酶(NR)测试盒。测定原理是硝酸还原酶催化植物体的硝酸盐还原为亚硝酸盐。产生的亚硝酸盐与显色剂反应生成红色化合物,在540 nm处比色测定。
1.4.4功能叶光合效能的测定 光合作用采用LI-6400XT便携式光合作用测量系统进行测定,光源设置为自然光源,控制叶片周围CO2浓度、H2O浓度、温度、相对湿度、光照强度和叶室温度等相关环境因子在相同的水平。
2 结果与分析
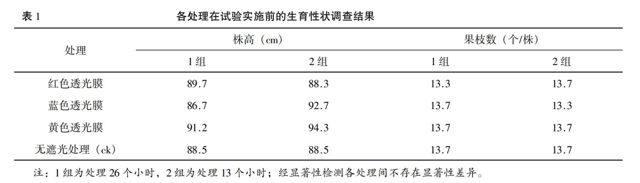
2.1 试验前棉花生育性状调查
从7月23日调查结果看(见表1),试验田棉花的果枝数在13~14个,株高为86.7~94.3 cm,小区间生育指标差异不显著,生长趋势一致,不存在差异显著性,符合试验要求。
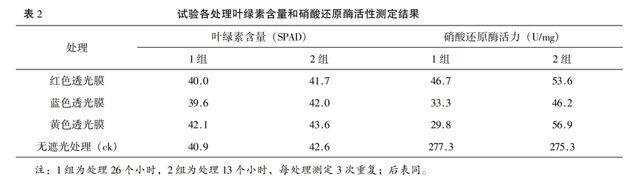
2.2 不同色膜遮光处理对棉花功能叶叶绿素含量的影响
从表2可以看出,在两组试验中均以黄色透光膜处理的叶绿素含量值最大,其次是对照,再次是蓝色透光膜,最后是红色透光膜,顺序是:黄色透光膜>无遮光处理(ck)>蓝色透光膜>红色透光膜,黄色透光膜处理的叶绿素含量最大值为45.5 SPAD,高于太阳光下的叶绿素合成,红色透光膜处理的最小值低至38.9 SPAD。随着色膜遮光处理时间延长,其叶绿素含量值低,如各处理第1组的值低于第2组,且差异达到显著性。结果表明黄色透光膜有利于激发叶绿素合成。
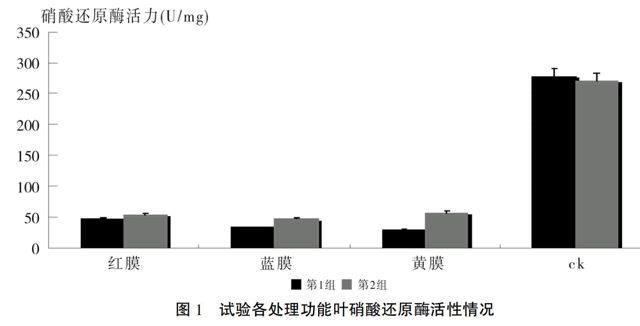
2.3 不同色膜遮光处理对棉花功能叶硝酸还原酶活性的影响
在不同色光处理下,功能叶的硝酸还原酶活性值依大到小,2组(处理13个小时)为无遮光处理(ck)>黄色透光膜>红色透光膜>蓝色透光膜,且3种色膜遮光处理与对照(无遮光处理)的比值分别为:黄色透光膜20.7%、红色透光膜19.5%、蓝色透光膜16.8%;而1组(处理26个小时)为无遮光处理(ck)>红色透光膜>蓝色透光膜>黄色透光膜,3种色膜遮光处理与对照(无遮光处理)的比值分别为:黄色透光膜10.7%、红色透光膜16.8%、蓝色透光膜12%(见表2、图1)。结果显示,色膜遮光处理时间长(第1组),以红色透光膜处理的功能叶硝酸还原酶活性值较蓝色透光膜和黄色透光膜大;而色膜遮光处理时间短(第2组),以黄色透光膜处理的功能叶硝酸还原酶活性值较红色透光膜和蓝色透光膜大,黄色透光膜处理是随着时间的延长先高后低,且他们之间的差异存在显著性。没有色膜遮光的处理(即太阳光)与遮光的3个处理间均存在极显著性差异,且3个色膜遮光处理的功能叶硝酸还原酶活性值均不及太阳光1/4。说明遮光处理抑制了功能叶硝酸还原酶活性。
2.4 不同色膜遮光处理对棉花功能叶光合效能的影响
太阳光经不同色光学膜透射后光强降低程度不同,从表3可以看出,红色透光膜处理的光强为381.5 μmol/m2·s,只有太阳光的29.5%;蓝色透光膜处理的光强为443.3 μmol/m2·s,只有太阳光的34.4%;黄色透光膜处理的光强为931.9 μmol/m2·s,占太阳光的72.1%。说明黄色透光膜处理光强削弱最少,其次是蓝色膜,红色膜削弱程度最大。
不同色光下蒸腾作用和胞间CO2浓度也会不同,试验第2组(处理13个小时后),红、蓝和黄3个不同色膜遮光处理的蒸腾作用分别为7.7 mmol/m2·s、6.3 mmol/m2·s、6.7 mmol/m2·s,差异不显著,而对照为14.3 mmol/m2·s,3个色膜遮光处理的蒸腾作用均约为太阳光的1/2。随着处理时间的增加,第1组(处理时间为26小时)红、蓝和黄3个色膜遮光处理功能叶的蒸腾作用分别为5.9 mmol/m2·s、9.5 mmol/m2·s、10.5 mmol/m2·s。第1组比第2组的蒸腾作用红色透光膜下降了23%,蓝色透光膜上升了50.8%,黄色透光膜上升了56.7%, 但是均低于对照,且具有显著性差异。说明红色透光膜不利于蒸腾作用,蓝色透光膜和黄色透光膜对蒸腾作用的削弱程度远小于红色光膜。
胞间CO2浓度是植物叶片(G)在光合生理生态研究中经常用到的一个参数。特别是在光合作用的气孔限制分析中,Ci的变化方向是确定光合速率变化的主要原因和是否为气孔因素的必不可少的判断依据[6]。在处理13小时后,色膜遮光处理下胞间CO2浓度红色透光膜为309.5 μμmol/mol,高于对照6.6%;蓝色透光膜为271.1 μmol/mol,低于对照6.7%,黄色透光膜为245.7 μmol/mol,低于对照18.2%。说明黄色透光膜有利于光合作用的进行,红色透光膜却抑制了光合作用的进行。随着色膜遮光处理时间的加长(处理26小时后),各处理胞间CO2浓度都升高了,红色透光膜升高了4.2%,蓝色透光膜升高了8.5%,黄色透光膜升高了25%。
净光合作用是衡量光合效能的最直接指标,在不同色膜遮光处理下,净光合作用差异明显。在处理13小时候后,红色透光膜处理的净光合作用为11.4 μmol/m2·s、蓝色透光膜为10.8 μmol/m2·s、黄色透光膜为13.4 μmol/m2·s、对照为25.4 μmol/m2·s,且各色膜遮光处理均小于对照。随着处理时间的延长,净光合作用发生了变化,处理26个小时后测定结果显示:红色透光膜的净光合作用下降6.1%,蓝色透光膜的净光合作用提高22.2%,黄色透光膜的光合作用提高58.2%。说明黄色透光膜处理最有利于叶片光合作用的进行。
在不同色膜遮光处理下,外部光强不一样,光量子大小不一,光合效能就呈现不一样的结果,胞间CO2浓度、蒸腾作用、净光合作用都呈现不一样的变化趋势。如表3所示,外部光强、蒸腾作用和净光合作用大小顺序均为:ck >黄色透光膜>蓝色透光膜>红色透光膜,且处理时间长,其值明显降低;胞间CO2浓度在色膜遮光处理下各色都呈现上升趋势,在开始时只有红色透光膜处理高于对照,随着处理时间的延长,3个色膜遮光处理都高于对照。
3讨论
试验选取长势一致的、在盛蕾期的棉花功能叶作为测定对象,通过不同色膜遮光处理,测定叶绿素含量、硝酸还原酶活力和光合效能,以探讨何种色光有利于功能叶的光合作用进行。
在本研究中,黄色透光膜遮光处理下的功能叶叶绿素含量高于对照,硝酸还原酶活性在处理13个小时后大于蓝色透光膜和红色透光膜,在处理26个小时后降低,低于红色和蓝色两种透光膜,但3个色膜遮光均远小于对照;光合效能(胞间CO2浓度、外部光强、蒸腾作用、净光合作用)也是黄色透光膜处理小于对照,而大于蓝色和红色两种透光膜处理。由此可见,叶片光合作用与叶绿素含量呈正相关,这与Leonard Beevers研究表明叶片光合作用与RuBP 羧化酶活性、叶片全氮含量、可溶性蛋白和叶绿素含量呈正相关的结果相一致[7]。
高等植物硝酸还原酶( NR, nitrate reductase, EC1. 6. 6. 1) 是植物氮代谢的关键酶和限速酶。 它是底物诱导酶, 同时也受到光的调控[8]。通常在有 NO3存在时, 在黑暗下植物也有硝酸还原酶活性( NRA, n-itrate reductase act ivity ) , 但活性很低; 一经照光, 硝酸还原酶活性大大提高。光照可通过加强植物对硝酸盐的吸收而提高NRA。光对植物 NRA 的快速调控, 对维持植物正常的生理代谢具有非常重要的意义[9]。在20世纪70 年代以来,国内外用体内测定法对大豆氮代谢中起重要作用的硝酸还原酶( NR) 进行了广泛研究, 认为光合作用与叶片NR 活性呈正相关[10]。该研究不同色膜遮光处理下硝酸还原酶活性变化趋势与净光合作用的变化趋势一致,进一步佐证了光合作用与叶片NR 活性呈正相关。光对硝酸还原酶的诱导起重要作用,蓝光的诱导相对稳定,红光诱导作用最弱,黄光在开始的时候比较强,随着处理时间加长而减弱。余让才,等[4]在研究不同光质对黄化小麦幼苗硝酸还原酶和乙醇酸氧化酶诱导的影响中得出:在蓝光、红光及白光中, 蓝光对硝酸还原酶和乙醇酸氧化酶诱导的能力最强, 而红光最弱;红光和蓝光诱导叶绿素积累的能力比较接近, 都低于白光,在光诱导的0~24 h 时期内, NR 活性上升较快, 而在24~48 h 时期内, 红光和白光下 NR 活性上升较小, 而蓝光处理还有一定的增加;在3种光质中, 蓝光诱导NR 活性的能力最强, 红光最弱, 白光介于蓝和红光质之间,红光处理材料的NR活性只有蓝光处理的50% 左右。本研究结果与这一结论相近。硝酸还原酶活性在采用光学膜遮蔽后,活性下降很大,仅仅只有对照的20%,这与许德威,等[5]的研究中得出的结论相一致。
不同光源对棉株光合作用的影响,从外部光强就可以看出黄色透光膜的光强最接近自然光源,其次是蓝色透光膜,最小的是红色透光膜。各色透光膜产生的光强不一样,其蒸腾速率、胞间CO2浓度、净光合作用就不一样,表现出黄色透光膜处理的光合效能仅次于自然光。在现代农业生产中,随着环境的恶化,雾霾越来越频繁,将成为人类和植物共同的环境灾害,不亚于干旱、暴雨等自然灾害。研究不同色光对植物光合作用的影响,将有助于人类在应对雾霾的气候时选择人工光源做出科学的依据。
致谢
本研究得到湖北省农业科学科院经济作物研究所棉花栽培团队的全力配合,武汉大学研究生宋成攀的协助,国家现代农业棉花产业技术体系、湖北省农业科技创新中心基金和湖北省农业科学院青年科学基金的经费资助,特此致谢!。
参考文献
[1] Sasakawa H,Yamamoto Y. Effects of red,far red and blue light on enhancement of nitrate reductase activity and on nitrate uptake in etiolated rice seedings[J]. Plant Physiol,1979(63):1098-1101.
[2] Vijayaraghavan SJ,Sopory SK,Guha-Mukherjee S. Role of light in the regulation of the nitrate xreductase level in wheat (Triticum aestivvm)[J]. Plant Cell Physiol,1979(20):1251-1261.
[3] 段远霖,李合生.不同光质和钙对小麦幼苗硝酸还原酶和谷氨酰胺合成酶活性的影响[J].植物生 理学通讯,1999(2):122-125.
[4] 余让才,范燕萍,李明启. 光对黄化小麦幼苗硝酸还原酶和乙醇酸氧化酶诱导的影响[J].中山大学学报论丛,1997(5):59-62.
[5] 许德威,周庆祺. 硝酸还原酶活力与棉株营养生长关系的研究[M]. 中国棉花学会第六次学术讨论会论文汇编,1986.
[6] 许大全.光合作用气孔限制分析中的一些问题.植物生理学通讯[J].1997(4):241-244.
[7] Leonard Beevers. The role of light and nitrate in the induction of nitrate reductase in radish cotyledons and maize seedings[J]. Plant Physiology,1965(40):691-695.
[8] 赵宏伟,马凤鸣,李文华.氮肥施用量对春玉米硝酸还原酶活性及产质量的影响[J].东北农业 大学学报, 2004,3(35):276-281.
[9] 余让才,范燕萍,李明启.蓝光对黄化小麦幼苗吸收及硝酸还原酶活性的影响[J].华南农业大 学学报,1996,17(3):70-74.
[10] Lenard Beevers, hageman R H.Nitrate reduction in haigher plants[J]. Ann Rev Plant Physilogy, 1969(20):495-498.
?收稿日期:2014-11-05
项目基金:国家现代农业棉花产业技术体系建设项目(CARS-18-22);湖北省农业科技创新中心资助项目(2007-620-001-03);湖北省农业科学院青年科学基金项目(2014NKYJJ21)。
作者简介:张友昌(1984-),男,硕士,zhych307@163.com。
通讯作者:别墅,研究员 bieshu02@163.com。
猜你喜欢
科学(2022年4期)2022-10-25 02:43:00
发明与创新(2019年43期)2019-11-16 06:59:44
学苑创造·A版(2018年5期)2018-05-28 12:26:30
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
求学·理科版(2016年4期)2016-09-02 04:04:04
Coco薇(2016年5期)2016-06-03 09:17:41
考试周刊(2016年6期)2016-03-11 08:14:32
少儿科学周刊·儿童版(2015年10期)2015-11-07 03:45:31
高中生学习·高二版(2014年5期)2014-07-03 14:50:01
青苹果(2014年1期)2014-04-09 20:31:00
