
黑尾近红鲌3种同工酶的组织特异性研究
2015-04-29 12:43:52陈见等
安徽农业科学 2015年29期
关键词:同工酶
陈见等

摘要[目的]了解黑尾近红鲌的遗传特性。[方法]采用聚丙烯酰胺凝胶垂直平板电泳,对黑尾近红鲌肝、肾、心、脑、肌和眼6种组织中乳酸脱氢酶(LDH)、酯酶(EST)及苹果酸脱氢酶(MDH)进行电泳分析。[结果]LDHB4在心脏和脑中优势表达,LDHA4在肌肉中优势表达,而LDHC只在肝脏中检测到;EST在肾、心、肝中活性较强,其中脑和肝组织有6条带,肾组织有5条带;mMDH在心脏中表现为典型的二聚体2条酶带,sMDH在所分析的6种组织都有分布。[结论]黑尾近红鲌6种组织中3种同工酶酶带的组成及活性均有所不同,表现出较明显的组织特异性。
关键词黑尾近红鲌;同工酶;组织特异性
中图分类号S965.199文献标识码A文章编号0517-6611(2015)29-049-03
黑尾近红鲌(Ancherythroculter nigrocauda Yih et Wu)又名高尖、黑尾鲌、黑尾、大白刁等,隶属鲤形目、鲤科、鲌亚科、近红鲌属,属于肉食性偏杂食性鱼类,是长江上游特有经济鱼类,其营养丰富、肉质细嫩、味道鲜美,深受养殖户喜爱。该品种具有养殖成本低、生长速度快、耐低氧能力强、易捕捞、易活鱼上市、食性杂、抗病力强等优势,是淡水养殖领域中新的优良、高效养殖对象之一。
同工酶是指具有相同的催化活性且在电场中的运动存在多种可分离形式的酶。同工酶在鱼类各组织中具有显著特异性,已广泛地用于鱼类分类鉴定、个体发育、杂种与种质鉴定、种群遗传多样性及进化分析、遗传育种等方面[6-7]。
笔者采用聚丙烯酰胺凝胶电泳技术,对黑尾近红鲌的乳酸脱氢酶(LDH)、酯酶(EST)及苹果酸脱氢酶(MDH)等生化遗传特性进行研究,以期为黑尾近红鲌的种质鉴定及选择育种提供一定的基础数据。
1材料与方法
1.1试验材料
试验用黑尾近红鲌来自国家级“湖北武汉鲌鱼良种场”。
1.2仪器与试剂
1.2.1仪器。北京六一DYY-6C型电泳仪电源、北京六一DYCZ-24A型双垂直电泳槽、北京六一WD-9405B型脱色摇床、上海恒平JY6002型电子天平、德国Eppondorf公司5810R型台式高速冷冻离心机玻璃制匀浆器。
1.2.2试剂。丙烯酰胺、N,N′—甲叉双丙烯酰胺、TEMED催化剂、过硫酸铵、溴酚蓝、缓冲液和染色液相关试剂。
1.3方法
1.3.1样品液的制备。
对黑尾近红鲌采用断鳃放血方法致死,在冰浴条件下取出其肝、肾、心、脑、肌和眼6种组织并洗净。在冰上进行组织匀浆,匀浆缓冲液为pH 7.0、浓度 0.1 mol/L磷酸缓冲液,将已匀好的匀浆液在4 ℃条件下静置1 h后,15 000 r/min、4 ℃下离心20 min后取上清液,置于-20 ℃条件下保存备用。
1.3.2聚丙烯酰胺凝胶垂直电泳。
采用不连续聚丙烯酰胺凝胶垂直电泳,凝胶制备参照《现代分子生物学实验技术》,电极缓冲液为pH8.3的Tris-甘氨酸溶液,在4 ℃条件下进行预电泳30 min。
经预电泳后,以5∶1的比例混合的酶液和溴酚蓝指示剂,用微量取液器取50 μl混合好的酶液加入点样孔。在4 ℃条件下正式电泳,当样品在浓缩胶迁移时电压控制为50 V,等到样品进入分离胶后电压升至200 V,直至溴酚蓝指示剂距分离胶底部1 cm时停止电泳。
1.3.3染色和固定。
乳酸脱氢酶、酯酶和苹果酸脱氢酶3种酶的染色方法分别参照张庆朝、赵赣等[10-11]。将凝胶于37 ℃染色液中避光染色,显色完成后,将凝胶板用去离子水漂洗2~3次,在7%的冰醋酸溶液中固定,对染色和固定好的凝胶板用相机进行拍照并进行分析。
2结果与分析
2.1乳酸脱氢酶(LDH)
目前,已知编码鱼类的LDH同工酶基因座包含3个位点,存在多种形式,其中LDH-C位点编码单独编码C蛋白且靠近阴极[12]。从图1可以看出,黑尾近鲌LDH同工酶活性具有组织特异性,在肾、心、脑、肌、眼组织中均可见5条酶带,其中LDH-B4在心脏和脑中优势表达,LDH-A4在肌肉中优势表达,而LDH-C只在肝脏中检测到,这种特异性与不同组织的相应生理功能密切相关。
2.2酯酶(EST)
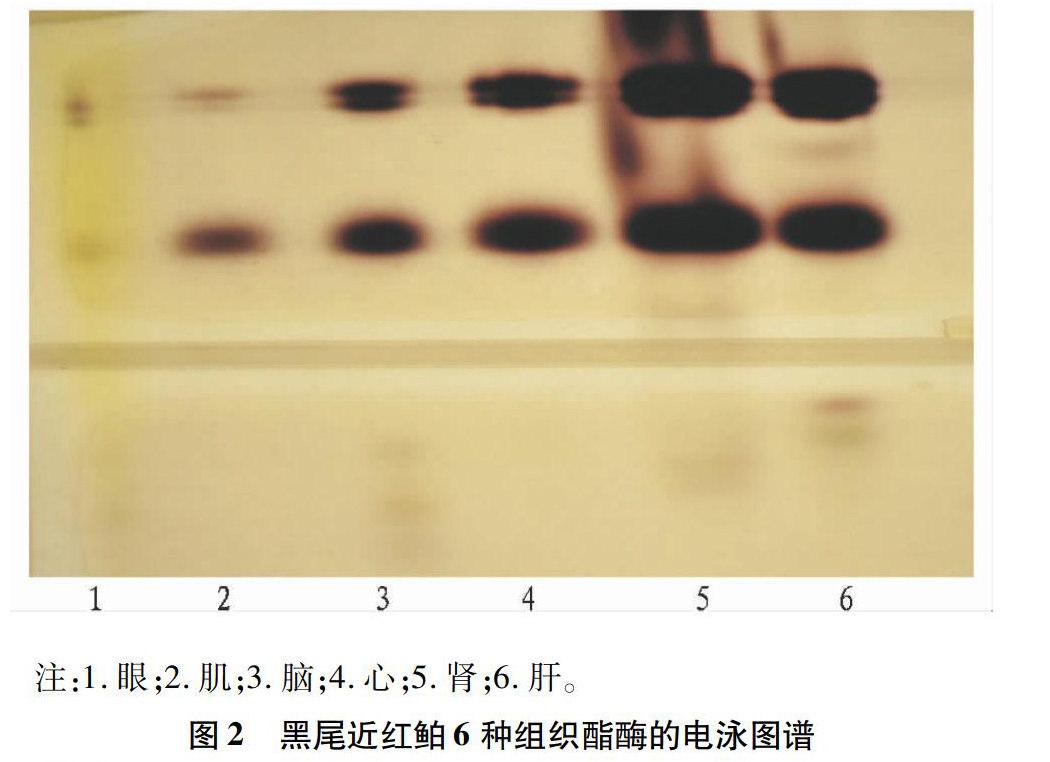
鱼类酯酶比较复杂,以单体酶或多聚体酶等形式存在,且其编码基因位点较多,呈现出普遍的多态性。从图2可以看出,EST在黑尾近红鲌的6种组织中均有分布,在肾、心和肝中活性较强,其中脑和肝组织有6条带,肾组织有5条带,这3种组织中的酯酶分布形式最多。
2.3苹果酸脱氢酶(MDH)
MDH同工酶在鱼类以二聚体形式存在,包括细胞质型(s-MDH)和线粒体型(m-MDH)2种类型,且均由2个基因位点编码[14]。从图3可以看出,黑尾近红鲌的m-MDH在心脏中表现为典型的二聚体2条酶带,在脑和肌中亦有分布。s-MDH在6种组织都有分布,其中肝、肾、心和脑均为2条酶带,而其他组织为1条酶带,尤其在肝、肾及脑中表达活性最强。
3讨论
该试验结果表明黑尾近红鲌的6种组织中3种同工酶在组成和活性上表现出明显的组织特异性,这与相应组织器官的生理功能密切相關[15]。黑尾近红鲌同工酶组织特异性主要表现在以下方面:①同工酶表达的位点多态性,某些位点的同工酶仅在特定组织表达;②同工酶表达强弱不同,主要表现在同一位点不同组织的表达强弱不一样;③同工酶表达的等位基因不同,同一位点的在不同组织中会有不同的同工酶。
LDH是糖酵解过程中的重要酶,可将无氧下产生的丙酮酸还原为乳酸并产能供机体所需。鱼类的LDH同工酶研究比较深入,已证明LDH是四聚体,编码鱼类的LDH同工酶基因座包含3个位点,分别为LDHA、LDHB、LDHC,通常情况下四聚体LDH由A、B 2个亚基组合成5种同工酶,分别为B4、AB3、A2B2、A3B、A4,B亚基迁移速率大于A亚基,C位点编码的蛋白质电荷与A、B不同且靠近阴极。LDHB4主要位于好氧性器官组织(如心脏)中,LDHA4主要位于厌氧性器官组织肌肉中。该研究中的黑尾近红鲌肌肉组织中的LDHA4表达要强于其他组织,这与晁珊珊等[16]对齐口裂腹鱼研究结果及马波等[17]对兴凯湖翘嘴红鲌研究结果类似。C位点编码的蛋白在鱼眼中首先被发现,随后在一些鱼类肝脏中也发现C位点的表达。该研究中LDH-C仅在黑尾近红鲌肝脏中特异性表达,可能与其肝脏代谢旺盛密切相关。在对翘嘴鲌、蒙古红鲌、青梢红鮊及海南红鲌[17-19]的研究中也发现肝脏具有C酶带。
EST是一种水解类酶,可催化酯类化合物的水解,参与能量代谢,还可代谢大量非正常存在的酯类,起到解毒功能。一般认为,在硬骨鱼类中EST是以单体或二聚体形式存在,由多个位点编码,存在多态现象。鱼类肝和肾等组织不仅作为主要的能量代谢组织,也是重要解毒组织。该研究中黑尾近红鲌肾脏、肝脏及心脏中EST的含量比较丰富,这与其提供能量并降解代谢废物、毒素等复杂的生理功能相适应,在淡水黑鲷、白甲鱼[14]、岩原鲤 、刺鲤研究中有着相似的结果。
MDH是生物氧化重要酶类,其负责使苹果酸脱氢为草酰乙酸参与糖酵解后的有氧代谢[13,22];因此MDH在一些有氧代谢旺盛的组织(如肝、肾等)中含量丰富、染色深。黑尾近红鲌中的MDH存在细胞质型(sMDH)和线粒体型(mMDH)2种,在对褐点石斑鱼[22]和松江鲈鱼研究中也发现存在2种类型的MDH。这2种MDH都是在黑尾近红鲌肝脏和心脏中染色较深,这与其有氧代谢旺盛有着密切关联性。
参考文献
[1] 湖北水生生物研究所鱼类研究室.长江鱼类[M].北京:科学出版社,1976:115-116.
[2] 薛正楷.濑溪河黑尾近红鲌生物学的初步研究[D].重庆:西南师范大学,2001.
[3] 谭德清,王剑伟,严太明,等.黑尾近红鲌人工繁殖研究[J].长江流域资源与环境,2004,13(2):193-197.
[4] 李伟东,黄峰,王贵英,等.饲料脂肪水平对黑尾近红鲌生长性能及鱼体成分的影响[J].粮食与饲料工业,2014(11):56-59.
[5] 彭仁海,张国强,卢全伟,等.兴国红鲤同工酶的研究[J].安阳工学院学报,2006(2):29-32.
[6] 王宏伟,王安利,王维娜,等.鱼类同工酶研究进展[J].动物学报,2009,47(S1):101-105.
[7] 刘鸿艳,谢从新.鱼类同工酶应用及研究进展[J].水利渔业,2006,26(5):1-3.
[8] 卢圣栋.现代分子生物学实验技术[M].2版.北京:中国协和医科大学出版社,1999:383-384.
[9] 张庆朝.鱼类乳酸脱氢酶同工酶聚丙烯酰胺凝胶电泳技术研究[J].生物技术,1994,4(5):38-40.
[10] 赵赣,钱芳.酯酶同工酶的一种新染色法的初步研究[J].江西农业大学学报,2002,24(3):380-382.
[11] 赵赣,张守全.一种新的苹果酸脱氢酶同工酶染色法[J].江西农业大学学报,2000,22(4):589-590.
[12] 杨玲,卢红,刘羽清,等.东平湖鲤8种同工酶的组织特异性研究[J].中国农学通报,2014,30(14):26-32.
[13] 李娴,王成武,安丽,等.淡水黑鲷5种同工酶的组织特异性研究[J].中国农学通报,2011,27(7):380-384.
[14] 王红叶,张娟,蔡焰值,等.白甲鱼不同组织同工酶的组织特异性研究[J].湖北农业科学,2011,50(23):4912-4915.
[15] 段彪,刘鸿艳.细鳞裂腹鱼同工酶组织特异性研究[J].西南大学学报(自然科学版),2010(6):27-30.
[16] 晁珊珊,宫佳琦,刀筱芳,等.齐口裂腹鱼及草鱼乳酸脱氢酶和草果酸脱氢酶同工酶比较研究[J].水产科学,2013,32(8):467-470.
[17] 马波,石连玉,尹家胜.兴凯湖翘嘴红鲌同工酶研究[J].水产学杂志,2000,13(2):63-68.
[18] 戈志强,宋学宏,许爱国,等.紅鲌属3种鲌鱼同工酶的比较研究[J].水利渔业,2005,25(6):23-24.
[19] 朱华平,黄樟翰,卢迈新.翘嘴红鲌和海南红鲌同工酶的比较研究[J].大连水产学院学报,2003,18(3):175-179.
[20] 王红叶,张娟,蔡焰值,等.白甲鱼不同组织同工酶的组织特异性研究[J].湖北农业科学,2011,50(23):4912-4915.
[21] 冯为慧,官少飞,欧阳敏,等.刺鲤同工酶的研究[J].安徽农业科学,2012,40(7):4029-4031.
[22] 尹绍武,廖经球,黄海,等.褐点石斑鱼不同组织4种同工酶的研究[J].海洋通报,2007,26(1):41-44.
[23] 王金秋,石椿.松江鲈鱼不同组织同工酶研究[J].复旦学报(自然科学版),2001,40(5):465-470.
猜你喜欢
遗传(2023年8期)2023-08-25 06:59:00
宁夏医学杂志(2020年3期)2021-01-21 08:23:38
微生物学杂志(2020年6期)2020-03-05 07:31:54
安徽农业科学(2019年7期)2019-03-16 02:30:13
现代检验医学杂志(2016年5期)2016-08-20 03:17:04
上海蔬菜(2016年6期)2016-04-04 18:57:33
湖南师范大学学报·自然科学版(2015年5期)2015-10-20 23:57:44
中国当代医药(2015年21期)2015-03-01 02:04:50
茶叶通讯(2014年2期)2014-02-27 07:55:39
癌变·畸变·突变(2014年4期)2014-02-27 06:14:34
