
放牧和围封对赖草草地群落空间分布特征的影响
2015-04-25 10:17任国华张宾宾张智袁赵祥
山西农业大学学报(自然科学版) 2015年5期
任国华,张宾宾,张智袁,赵祥
(山西农业大学 动物科技学院,山西 太谷030801)
草地植物群落的空间分布异质性是物种间相互关系及其利用环境资源状况综合作用的结果,是草地植被更新的生态学基础[1]。天然草地的科学管理和持续利用是草地畜牧业研究者的关注焦点。放牧利用作为草地管理的主要方式之一,不仅使草地植物的物种组成和群落结构发生改变,也引起草地功能的变化[2~5]。目前,国内学者从不同角度研究了草地植物种群动态、群落结构和功能对放牧的响应,如郑伟等[6]研究发现适口性较好的优良牧草优势度随着放牧强度的增加而降低,适口性差或家畜不食的植物优势度增加;赵景学等[7]对藏北地区3类高寒草地的放牧研究表明,植物群落盖度、高度和地上生物量均显著低于围栏封育样地,而短期禁牧对退化高寒草地群落物种多样性影响不显著;石福孙[8]和左万庆等[9]认为围栏封育能增加草地的生产力,但会影响草地群落结构和功能的完整性。因此,了解草地利用和管理方式如何作用于草地生态系统群落异质性,确定人类活动对草地空间格局的影响范围,对指导受损草地生态系统恢复和重建具有十分重要的意义。
不同地理区域环境条件差异较大,生态背景和草地类型各异,放牧利用和管理方式对草地生态系统异质性的影响也不同[10]。赖草(Leymus secalinus)草地是晋北地区重要的生态屏障和草地资源,是极具特色的草原畜牧业生产基地。晋北地区地处黄土高原东缘,草地生态系统脆弱且抗干扰能力差,近年来由于不合理的放牧利用以及对草地资源科学管理的忽视,草地系统的结构和功能不断退化,草畜矛盾日益突出,严重威胁晋北地区草地畜牧业的可持续发展和生态系统功能的稳定性。围栏封育作为一种简便易行且有效的草地恢复措施,在我国草地管理中越来越得到人们的共识,目前对草地放牧管理的研究主要集中在群落特征方面,群落中植物种群的空间异质性分布对放牧的响应研究不多。本研究以山西北部右玉县长期放牧和围封6年赖草草地为研究对象,探讨不同放牧管理方式对主要植物种群空间分布格局的影响,为赖草草地资源的保护性利用和可持续发展提供理论依据。
1 材料与方法
1.1 研究区概况
本研究在山西农业大学雁门关草地生态系统定位研究站进行,位于山西省西北部右玉县境内。其地理位置为北纬39°59′,东经112°19′,海拔1 350m。气候为温带大陆性季风气候,年均气温4℃,7月平均温度20℃,1月平均温度-13℃。年平均降水量420mm左右,主要集中分布于6-9月,年蒸发量约为1 000~1 500mm。全年日照时数2 650h,年均无霜期约为100~120d。土壤为苏打草甸碱土,全盐含量为0.44%,植被类型主要是低地草甸类盐化低地草甸亚类,以禾本科赖草属的赖草(Leymus secalinus)为优势种,主要伴生种有鹅绒委陵菜(Potentilla ansrina)、碱地风毛菊(Saussurea amara)、米 蒿 (Artemisia dalailamae)、车前(Plantago asiatica))、西伯利亚蓼(Polygonum sibiricum)、海乳 草 (Glaux maritima)、芦苇(Phragmites communis)等[10]。
1.2 研究方法
1.2.1 调查与取样
围栏草地封育时间为2008年,围栏内禁牧,围栏外放牧强度中等,放牧强度约为每公顷4.5只羊。2014年7月和8月调查草地植被特征,围栏内外分别选取100m×100m的样地,采用1m×1m的样方沿对角线依次取样。分别调查每个样方内物种种类、盖度、高度和密度;地上生物量采用刈割烘干称重法,留茬高度为1cm,烘干后称重。
1.2.2 植物多样性测定
群落中物种丰富度指数(richness index)、均匀度指数(evenness index)、优势度指数(dominance index)和多样性指数(diversity index)根据物种重要值计算,公式如下:
重要值=(相对盖度+相对高度+相对密度+相对生物量)/4
丰富度指数:R=S
Shannon-Wiener多样性指数:
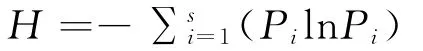
式中:S为每个样方中的出现的总物种数;Pi为样方中种i的重要值。
1.2.3 物种多度拟合计算
物种多度等级分布拟合分别采用分割线段模型(Broken-stick model)、生态位优先占据模型(Niche preemption model)、对数正态分布模型(Log-normal model)、Zipf模 型 (Zipf model)和Zipf-Mandelbrot 模 型 (Zipf-Mandelbrot model)[11,12]。根 据 每 个 模 型 的 赤 池 信 息 量 准 则(Akaike’s Information Criteria,AIC)选择物种多度分布模式最优的拟合模型,AIC值越小,模型拟合效果越好[13]。
1.2.4 半方差函数计算
草地群落中赖草的空间异质性采用线性模型(Linear model)、指数模型(Exponential model)、球状模型(Spherical model)和高斯模型(Gaussian model)进行拟合,根据决定系数(R2)和残差平方和(RSS)评价4个模型拟合效果。半方差函数主要参数包括:块金值(C0),基台值(C0+C)、结构比(C/C0+C)、范围参数(a0)、决定系数(R2)和残差平方和(RSS)等[14~16]。其中,块金值表示随机性因素引起的空间异质性;基台值为研究变量在整个系统中的最大变异程度;结构比代表结构方差在空间总异质性变异中的大小;范围参数反映调查变量在空间上的自相关程度[11,12]。
2 结果与分析
2.1 放牧和封育对草地群落组成的影响
调查结果显示,自由放牧样地和围栏封育样地群落建群种均为赖草,且重要值所占比例较大,分别为0.64和0.79;放牧区主要伴生种为鹅绒委陵菜、米蒿和冷蒿,封育区主要伴生种为披碱草、芦苇和碱茅;样地间物种相似度指数为0.433,说明群落组分种在草地长期封育过程中发生了一定程度的改变(表1)。封育措施对草地群落特征产生明显影响,由表2可见,封育草地的盖度、高度和地上生物量显著高于放牧地(P<0.01);放牧草地相对于围封样地具有较高的优势度指数和多样性指数(P<0.05)。

表1 围栏内外草地群落物种重要值Table 1 Important values of plant species under enclosure and grazing plots
2.2 放牧和封育对草地物种-多度格局的影响
分别采用分割线段模型、生态位优先占据模型、对数正态分布模型、Zipf模型和Zipf-Mandelbrot模型对放牧样地与封育样地群落的物种多度分布格局进行拟合,根据各个模型的AIC值(Akaike’s Information Criteria)最小原则选择拟合的最优模型。从表3中可以看到,放牧和封育对草地群落物种多度等级分布模式的影响不同,放牧草地群落物种多度分布格局符合对数正态分布模型和Zipf-Mandelbrot模型,围封草地中符合生态位优先占据模型。
2.3 放牧和封育对赖草空间格局的影响
不同处理区赖草空间分布变异函数的参数值如表4所示,放牧样地和封育样地的块金值分别为0.165和0.001,表明放牧样地赖草空间异质性主要由放牧家畜采食和践踏等随机性因素决定,几乎不受结构性因素(如气候、地形、土壤母质和利用方式等)影响;结构比分别为0.626和0.997,表明赖草密度的空间分布在封育样地中具有强烈的空间自相关性,即封育措施影响了赖草空间分布的结构属性。根据决定系数和剩余残差大小,放牧样地赖草空间分布半方差函数中,高斯模型拟合效果较好,围栏封育区最适空间变异函数为球状模型。

表2 围栏内外草地群落盖度、高度、生物量、物种丰富度、均匀度、优势度和多样性指数的变化Table 2 Change of cover,height,biomass,species number(S),evenness index(J),domination index(D)and diversity index(H)between enclosure and grazing plots

表3 围栏内外草地群落物种多度分布模型拟合AIC值Table 3 AICvalues of different models under enclosureand grazing plots
3 讨论
围栏封育是草地生态系统最常见的管理方式,是影响草地生产力和物种多样性的重要因素[4]。本研究表明,围栏封育6年后的赖草草地总盖度、平均高度和地上生物量显著高于围栏外自由放牧样地,这与李媛媛等[3]、刘晓妮等[10]的研究结果一致。本实验中放牧样地物种丰富度和多样性明显高于围封样地,这也与“中度干扰理论”[17,18]相符合。在草地生态系统中,植物种群的空间格局是群落构建的基本生态过程之一,是空间异质性的具体体现[14]。本研究中放牧样地赖草空间分布格局主要由随机性因素引起,即由家畜的选择性采食、践踏和游走等牧食行为异质性决定[15]。另外,围栏封育使赖草最大空间变异程度减小,空间格局趋于简单化。这与禁牧使草地植物多样性降低,不同样点处的物种密度趋向统一有关。

表4 围栏内外草地中赖草空间分布的半方差函数分析Table 4 The semi-variance analysis of Leymus secalinus under enclosure and grazing plot
草原植物群落的物种多度和空间分布变化也是草原响应放牧干扰的重要特征之一,群落组分更替和植被类型改变是草地处于不同演替阶段的标志[2],围栏内外建群种没有发生更替,赖草种群均在群落中处于绝对支配地位,但组分种在群落中的地位和作用发生较大改变。通过对比放牧样地和围封样地的物种多度分布格局发现,对数正态分布模型和Zipf-Mandelbrot模型都可以拟合放牧样地中物种多度分布模式,围封样地则符合生态位优先占据模型。对数正态分布假说认为物种的存在受到微气候、空间和食物资源等诸多环境因素的影响,导致物种多度呈正态分布;Zipf-Mandelbrot假说认为物种对生态位的占有基于环境潜在多样性;而生态位优先假说认为第一个物种占用群落生态位空间的大部分,第二个物种占用余下的大部分,依此类推,末位的只能占有很少的空间[11]。草地在长期围封禁牧作用下,可以预测,潜在群落会向赖草单顶级群落方向演替,不干扰策略会逐步增加其群落的稳定性,不利于草地植物多样性的保护与维持。
草地植被经历较多的干扰时会导致群落内部物种间相互作用发生变化,因而使其构成的群落空间格局改变,外部作用替代草地群落内部植物间或与微环境的相互影响,进而形成新的草地群落空间格局[19]。杨晓晖等[20]研究显示,短期围栏封育草地盖度和地上生物量均有增加,但草地长时间围封并不能增加草地的生产力;左万庆等[9]发现长期草地围栏会导致草地植物多样性下降;较多研究[7,17,20]表明,草地过度放牧会造成生物多样性下降和草地生态破坏。由于本实验仅为中度放牧与围栏封育两种情形,随机因素和结构因素在草地围栏内外引起的植物群落空间异质性变化还有待于进一步研究和考证。放牧试验年限、放牧组合和放牧强度等具体参数的量化,对维持放牧草地系统结构和功能的稳定性,逐步建立可持续发展的畜牧业具有重要的理论和实践意义。
[1]Audrey A,Vincent B,Sandrine P.Spatial patterns of weeds along agradient of landscape complexity[J].Basic and Applied Ecology,2012,13:328-337.
[2]刘雪明,聂学敏.围栏封育对高寒草地植被数量特征的影响[J].草业科学,2012,29(1):112-116.
[3]李媛媛,董世魁,李小艳,等.围栏封育对黄河源区退化高寒草地植被组成及生物量的影响[J].草地学报,2012,20(2):275-286.
[4]高宁宁,陈俊,张鹏莉,等.放牧对西藏高寒嵩草草甸地上生物量空间分布的影响[J].草地学报,2014,22(2):256-260.
[5]益西措姆,许岳飞,付娟娟,等.放牧强度对西藏高寒草甸植被群落和土壤理化性质的影响[J].西北农林科技大学学报(自然科学版),2014,42(6):27-33.
[6]郑伟,董全民,李世雄,等.放牧对环青海湖高寒草原主要植物种群生态位的影响[J].草业科学,2013,30(12):2040-2046.
[7]赵景学,祁彪,多吉顿珠,等.短期围栏封育对藏北3类退化高寒草地群落特征的影响[J].草业科学,2011,28(1):59-62.
[8]石福孙,吴宁,罗鹏,等.围栏禁牧对川西北亚高山高寒草甸群落结构的影响[J].应用与环境生物学报,2007,13(6):767-770.
[9]左万庆,王玉辉,王风玉,等.围栏封育措施对退化羊草草原植物群落特征影响研究[J].草业学报,2009,18(3):12-19.
[10]刘晓妮,李刚,赵祥,等.放牧对赖草群落生物量及植物多样性的影响[J].草地学报,2014,22(5):942-948.
[11]程佳佳,米湘成,马克平,等.亚热带常绿阔叶林群落物种多度分布格局对取样尺度的响应[J].生物多样性,2011,19(2):168-177.
[12]闫琰,张春雨,赵秀海.长白山不同演替阶段针阔混交林群落物种多度分布格局[J].植物生态学报,2012,36(9):923-934.
[13]Burnham KP,Anderson DR.Model selection and Multi-Model Inference.A Practical Information-Theoretic Approach[M].New York:Springer-Verlag,2002:49-96.
[14]吴艳玲,陈立波,卫智军,等.不同放牧压短花针茅荒漠草原群落植物种的空间异质特征[J].干旱区资源与环境,2012,26(7):110-115.
[15]希吉日塔娜,吕世杰,卫智军,等.不同放牧制度下短花针茅草原主要植物种群的空间变异[J].中国草地学报,2013,35(2):76-82.
[16]吕世杰,刘红梅,吴艳玲,等.放牧对短花针茅荒漠草原建群种与优势种空间分布关系的影响[J].应用生态学报,2014,25(12):3469-3474.
[17]李强,杨劼,宋炳煜,等.不同围封年限对退化大针茅草原生产力和土壤碳、氮贮量的影响[J].生态学杂志,2014,33(4):896-901.
[18]Doyle JJ,Doyle JL.A rapid DNA isolation procedure for small quantities of fresh leaf tissue[J].Phytochemical Bulletin,1987,19:11-15.
[19]刘振国,李镇清.不同放牧强度下冷蒿种群小尺度空间格局[J].生态学报,2004,24(2):227-234.
[20]杨晓晖,张克斌,侯瑞萍.封育措施对半干旱沙地草场植被群落特征及地上生物量的影响[J].生态环境,2005,14(5):730-734.
猜你喜欢
林业资源管理(2022年6期)2023-01-18
天津农林科技(2022年2期)2022-04-19
河南畜牧兽医(2022年3期)2022-04-13
现代园艺(2021年23期)2021-12-01
今日农业(2021年2期)2021-11-27
绿色科技(2021年10期)2021-06-23
林业勘查设计(2020年1期)2021-01-18
新农业(2020年18期)2021-01-07
好孩子画报(2019年8期)2019-09-19
草业学报(2019年2期)2019-02-25
