
赤水河上游主要树种根际土壤调控水分功能研究
2015-04-20 11:52喻阳华李光容严令斌黄宗胜喻理飞
水土保持研究 2015年5期
喻阳华, 李光容, 严令斌, 黄宗胜, 喻理飞
(贵州大学 林学院, 贵阳 550025)
赤水河上游主要树种根际土壤调控水分功能研究
喻阳华, 李光容, 严令斌, 黄宗胜, 喻理飞
(贵州大学 林学院, 贵阳 550025)
为明确赤水河上游主要树种根际土壤调控水分能力,通过野外采样和室内分析方法,从容重、含水率和孔隙度等方面入手研究水文生态功能,并划分调控水分功能群。结果表明:32个树种0—60 cm的最大持水量为212.00~357.94 mm、有效贮水量为11.07~85.15 mm,是一个蓄水库;其持水速率大于失水速率,且二者与时间的关系均符合幂函数模型。采用RDA排序时,可将该区的树种划分为5类功能群:功能群Ⅰ包括杨梅、丝栗栲、茶、白栎、马尾松、慈竹;功能群Ⅱ包括杨树、杉木、构树、荚蒾、撑绿竹、枫香、火棘、山胡椒、乌桕、楝树、马桑;功能群Ⅲ包括枇杷、南天竹、李、柏木、梧桐、香椿;功能群Ⅳ包括檵木、黄荆、油桐、毛桐、南酸枣;功能群Ⅴ包括盐肤木、黄连木、柑橘、油茶。土壤容重、含水率、孔隙度等物理特征和持水过程是影响土壤水分调控能力的主因。研究结果可为赤水河上游水源涵养林树种的选择和配置提供理论依据。
树种根际土壤; 水文功能; 水分调控; 功能群; 赤水河上游
林地土壤最大蓄水量占总蓄水量的97%以上[1],它的孔隙状况反映了根系改善土壤物理性质的潜能,影响土壤蓄水保水和通气性能[2],一般以总持水量、毛管持水量和非毛管持水量作为蓄水性能指标[3]。其中,毛管孔隙度的大小反映了森林植被吸持水分用于维持自身生长发育的能力;非毛管孔隙的大小反映了森林植被滞留水分、发挥涵养水源功能和削减洪峰的能力[4-6]。根际是根系对土壤影响最直接、最强烈的区域[7],可用来反映根系对土壤孔隙和土壤质量的影响,揭示树种根系对土壤涵养水源的作用。水源涵养林功能的提高,取决于林分结构的优化,结构调整首先需要研究树种特性,正确选择树种,贯彻适地适树原则[8],即通过对种群和功能群尺度的研究揭示和上推群落尺度的作用机理。目前对森林土壤水文生态效应[4-6,9]研究主要集中在群落水平,以物种为单元、从功能群尺度研究土壤持水能力的报道较为鲜见。因此,本文以赤水河上游主要树种为单元、以根际土壤为研究对象,从功能群尺度考察其水文生态功能,主要研究以下3个问题:(1) 各树种土壤层水文效应的定量数据;(2) 各树种土壤层的持水过程和失水过程特征;(3) 根际土壤调控水分功能群组合。旨在为该区水源涵养林建设提供理论依据和技术支撑。
1 研究区概况
赤水河是长江中上游保持原生态状况的河流,国酒茅台主产区位于流域中下游,中上游区是水源涵养区,植被优劣影响下游河流生态系统,对维护国酒茅台特殊生产用水及酿造环境安全和区域生态安全及生态平衡有重要意义,并发挥着区域生物多样性和保持水土功能。本研究区主要为位于赤水河上游茅台水源功能区内的金沙县和仁怀市(105°21′32″—106°15′00″E, 27°39′48″—28°05′38″N),为亚热带季风湿润气候,多年平均气温为15℃,≥10℃的年积温为5 000~5 500℃,年日照时数1 400 h,无霜期约340 d,年均降水量>1 000 mm,土壤类型有石灰土、黄壤、紫色土等[10]。新中国成立以来,由于人为砍伐,使该区域原本脆弱的森林植被生产功能和承载功能下降,水土流失加剧,水源涵养能力降低,直接影响到茅台酒的酿造环境与品质。现存主要植被为暖性针叶林和常绿落叶阔叶林,丝栗栲林为演替顶极群落。目前赤水河水资源总量、多年平均径流量和最大年均径流量呈现减少趋势,也使该区开展水源涵养林建设具有重要性和紧迫性。
2 材料与方法
2.1 研究对象
根据《贵州植被》[11]及前人研究成果[10],确定该区域的乡土树种与地带性植物,结合实际调查,在乡土树种与地带性植物中选取如下32个树种作为研究对象:杨树(Populustrinervis)、盐肤木(Rhuschinensis)、杨梅(Myricaceaerubra)、丝栗栲(Castanopsisfargesii)、黄连木(Pistaciachinensis)、枇杷(Eriobotryabengalensis)、杉木(Cunninghamialanceolata)、南天竹(Nandinadomestica)、构树(Broussonetiapapyifera)、荚蒾(Viburnumchinshanense)、李(Prunusnalicina)、撑绿竹(Bambusapervariabilis)、茶(Camelliasinensis)、枫香(Liquidambaformosana)、白栎(Quercusaliena)、柏木(Cupressusfunebris)、梧桐(Platanusacerifolia)、马尾松(Pinusmassoniana)、檵木(Loropetalumchinense)、火棘(Pyracanthafortuneana)、乌桕(Sapiumsebiferum)、山胡椒(Linderaglauca)、慈竹(Neosinocalamusaffinis)、马桑(Coriarianepalensis)、柑橘(Citrusreticulata)、黄荆(Vitexnegundo)、油桐(Verniciafordii)、毛桐(Mallotusbarbatus)、南酸枣(Choerospondiasaxillaria)、楝树(Meliaazedarach)、香椿(Toonasinensis)、油茶(Camelliaoleifera),均以健康成年树为标准采集根际土壤。
2.2 研究方法
2.2.1 土壤调查与测定 选择天气条件较为一致的2014年3月6日至8日,乔木树种距主干≤50 cm范围,灌木树种移除整株后沿主根系挖掘1个60 cm深的土壤剖面进行观测记录,用环刀采集0—20,20—40,40—60 cm土层的土样,用烘干法和浸水法测定土壤自然含水率、土壤的各项物理性质和持水性能指标[3]。
2.2.2 土壤持水量与失水量测定 环刀浸水后静置,于0.5,1,2,4,8,12,24 h后称量持水质量;持水24 h后,将环刀带土置于平板上,于0.5,1,2,4,8,12,24 h后称量失水质量。每个树种设置3次重复。
2.2.3 土壤蓄水量的测定 土壤有效持水量和最大持水量采用如下公式计算[1]:
Wa=1000PnH
(1)
Ws=1000PtH
(2)
式中:Wa——土壤有效持水(mm);Pn——土壤非毛管孔隙度(%);H——土层深度(m);Ws——土壤最大持水量(m);Pt——土壤总孔隙度(%)。土壤总孔隙度、毛管孔隙度、非毛管孔隙度的计算见文献[12]。
2.2.4 功能群划分 选取容重、含水率、总孔隙度、毛管孔隙度、有效持水量、持水曲线常数项、持水曲线指数项、失水曲线常数项、失水曲线指数项9个指标进行功能群划分(由于非毛管孔隙度为总孔隙度与毛管孔隙度之差,为防止划分指标产生累积效应,因此不作为直接因子参与划分)。
2.3 数据处理
平均值计算用算术法,数据标准化预处理用归一化方法,均由Excel 2003完成;持水速率、失水速率与时间的拟合关系用回归分析方法,由SPSS 20.0软件完成;排序是在判断排序轴梯度长度的基础上,采用冗余分析(Redundancyanalysis,RDA)进行,由Canocofor Windows 4.5软件完成;图片处理采用修正法和填充法,由photoshop和Microsoft Office Visio 2003软件共同完成。
3 结果与分析
3.1 根际土壤持水能力
土壤的物理性质制约着它的持水与保水性能。如表1所示,不同树种0—60 cm土壤持水能力各异,饱和持水能力212.00~357.94 mm,有效贮水能力11.07~85.15 mm。就不同树种而言,根际土壤最大持水量最大为南天竹(357.94 mm)和枇杷(338.26 mm)、最小为丝栗栲(212.00 mm)和茶(221.64 mm),有效持水量最大为杉木(85.15 mm)和南天竹(73.54 mm)、最小为油桐(11.07 mm)和马桑(11.15 mm),表明它们改良土壤调控水分能力的潜力不同,进而影响土壤水分保持与运动及土壤对植物水分供应能力。产生这种现象的原因与枯落物组成、数量、分解程度及分解产物有关,也与地下根系发育与作用有关[2,5,13]。
同一树种各层的持水能力也具有差异,多表现为土层越深则有效持水能力越弱,原因可能为根系在不同厚度处的分布不均匀,也可能是不同土层有机质含量和土壤团聚性能不一样,进而形成土壤物理性质的差异[14]。此外,高大乔木根际土壤持水能力强于灌木或小乔木,这可能与高大乔木根系粗度和密度较大有关。马尾松、柏木持水能力低的原因可能是针叶树种凋落物不易分解,凋落物腐殖质后能够为表层土壤输入的有机质少,不利于毛管孔隙的形成[2],但杉木持水能力强的原因有待进一步研究。

表1 不同树种各土层持水能力 mm
3.2 根际土壤持水与失水过程
土壤持水速率和失水速率分别表征其持水、失水过程的快慢,二者共同反映了土壤延缓地表径流和截持降雨能力,决定森林土壤水分调控能力。本文是以0—60 cm土壤层代表其调控能力,实际的蓄水量和调控能力则更大。
如表2所示,1 h内,土壤持水速率和失水速率都较高,随着时间的推移而不断降低,4 h后趋于稳定。二者与时间具有一定的相关关系,均能用幂函数曲线进行拟合(y=axb),式中y为持水速率或失水速率[g/(kg·h)],x为时间(h),a,b为常数,R>0.99,相关系数较高。所有根际土壤同时段的持水速率均大于失水速率,表明土壤具有水源涵养、调节地表径流的功能,说明森林土壤水文生态效益显著,是森林生态系统的水分蓄库。持水速率高表征土壤滞留降雨的能力强,对水源涵养林树种选择具有重要参考价值。产生这一差异的因素包括:(1)树种类型。树种会通过枯落物和根系等影响土壤孔隙度性状,从而使其储蓄、调控水分能力不同;(2)砾石大小与含量[9]。影响土壤有效贮水能力的高低;(3)土壤密度[15]。制约土壤稳渗速率和水分贮存空间,对土壤水、气此消彼长的关系产生影响。
3.3 根际土壤调控水分功能群
植物功能群是用于研究基于植物功能特性的植物集群和物种反应的普遍法则[16],它的研究可把复杂生态系统通过功能团体划分而得以简单化[17]。赤水河上游生境要素的变化导致物种生态习性不同,在长期自然演替进程中形成稳定的功能组合。以协变量表示的背景变异为基础,大致将32个树种划分为5组植物功能群(图1)。
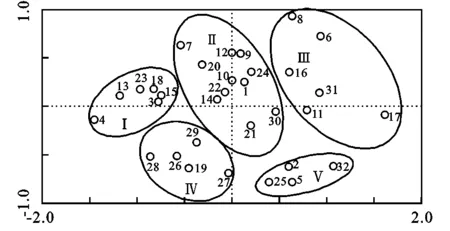
注:1杨树,2盐肤木,3杨梅,4丝栗栲,5黄连木,6枇杷,7杉木,8南天竹,9构树,10荚蒾,11李,12撑绿竹,13茶,14枫香,15白栎,16柏木,17梧桐,18马尾松,19檵木,20火棘,21乌桕,22山胡椒,23慈竹,24马桑,25柑橘,26黄荆,27油桐,28毛桐,29南酸枣,30楝树,31香椿,32油茶。
图1 根际土壤调控水分功能群划分结果
功能群Ⅰ包括杨梅、丝栗栲、茶、白栎、马尾松、慈竹,均生长于常态地貌(多为黄壤),面积大、分布广,林下更新幼苗数量较少,0—60 cm根际土壤有效持水量为30~50 mm,处于中等持水水平。

表2 土壤持水速率、失水速率与时间的拟合关系
注:**表示在0.01(双侧)上显著差异。
功能群Ⅱ包括杨树、杉木、构树、荚蒾、撑绿竹、枫香、火棘、山胡椒、乌桕、楝树、马桑,生长分布于常态地貌和喀斯特地貌的过渡区域,适生范围较广,但喀斯特地貌上的物种长势要差,根际土壤有效持水能力较低。
功能群Ⅲ包括枇杷、南天竹、李、柏木、梧桐、香椿,多为人工栽植的经济树种,受人为干扰程度较大,多分布于喀斯特地貌,根际土壤有效持水能力处于中等水平。
功能群Ⅳ包括檵木、黄荆、油桐、毛桐、南酸枣,均生长于喀斯特地貌,大面积分布但较分散,多与其它树种混交形成混交林,所构成的群落物种多样性水平较高,根际土壤有效持水能力处于中低水平。
功能群Ⅴ包括盐肤木、黄连木、柑橘、油茶,天然林中分布较少,为偶见种,根际土壤有效持水能力相对较低。
3.4 影响根际土壤持水能力的因素
如图2所示,从箭头连线的长短可知,在土壤厚度一定时,影响根际土壤持水能力的主要因素有容重、含水率、持水曲线常数项、毛管孔隙度和总孔隙度等,影响较小的是持水曲线指数项。土壤容重影响持水能力的原因是它指示土壤紧实度状况,表征土壤透水性、透气性和根系延展时的阻力大小[5];土壤含水率高低则可能制约饱和持水能力及土壤蓄水潜力与空间;土壤孔隙状况反映了本身结构和粒径分布特征,体现了土壤蓄水能力的高低。总孔隙度和毛管孔隙度的差值即为非毛管孔隙度,能迅速吸收降水并及时下渗,决定了土壤有效持水量,利于涵养水源[6]。
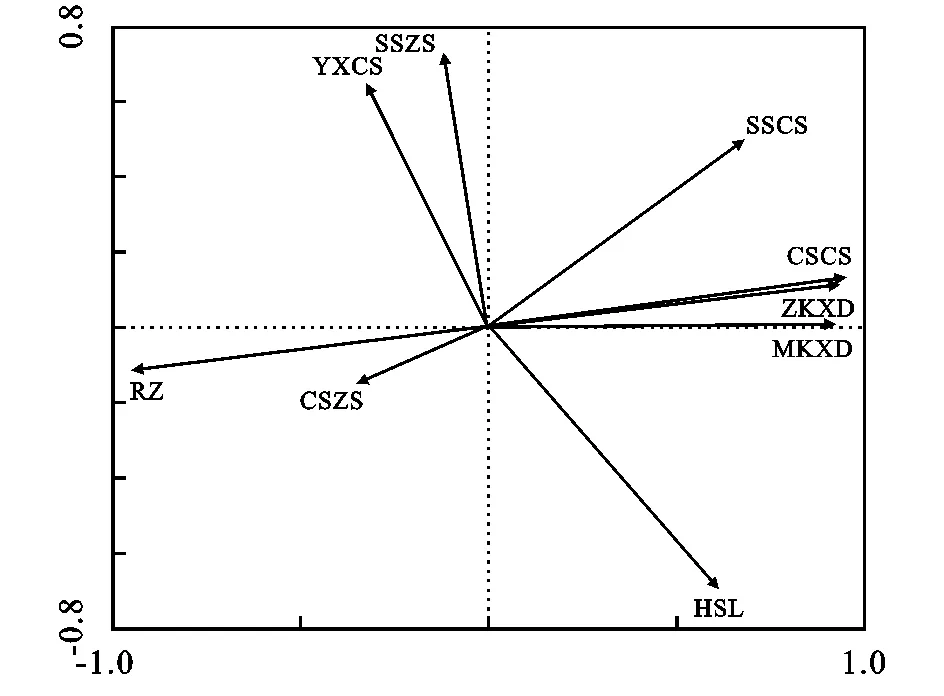
注:RZ容重、HSL自然含水率、ZKXD总孔隙度、MKXD毛管孔隙度、YXCS有效持水量、CSCS持水曲线常数项、CSZS持水曲线指数项、SSCS失水曲线常数项、SSZS失水曲线指数项。
图2 影响根际土壤调控水分能力的因素
4 结 论
赤水河上游32个树种0—60 cm根际土壤的持水速率均大于失水速率,且二者均与时间呈明显幂函数关系,相关性较好,具有较强的水分调蓄潜力,是一个蓄水库。将它们划分为5个功能群:功能群Ⅰ包括杨梅、丝栗栲、茶、白栎、马尾松、慈竹;功能群Ⅱ包括杨树、杉木、构树、荚蒾、撑绿竹、枫香、火棘、山胡椒、乌桕、楝树、马桑;功能群Ⅲ包括枇杷、南天竹、李、柏木、梧桐、香椿;功能群Ⅳ包括檵木、黄荆、油桐、毛桐、南酸枣;功能群Ⅴ包括盐肤木、黄连木、柑橘、油茶。划分结果能够较好地反映树种与持水能力的关系,辅助该区水源涵养林树种选择和布局。改善根际土壤物理性状应作为该区水源涵养林建设和植被恢复的措施之一。
[1] 丁访军,王兵,钟洪明,等.赤水河下游不同林地类型土壤物理特性及其水源涵养功能[J].水土保持学报,2009,23(3):179-183,203.
[2] 贾秀红,曾毅,周志翔,等.鄂中低丘区不同演替阶段森林凋落物和土壤水文特征[J].水土保持学报,2013,27(4):125-129.
[3] 赵建生,王永明,杨新兵,等.冀北山地森林土壤水文效应研究[J].水土保持研究,2013,20(3):201-205.
[4] 石媛,鲁绍伟,陈波,等.河北雾灵山不同密度油松人工林枯落物及土壤水文效应[J].水土保持学报,2014,28(3):92-97.
[5] 鲁绍伟,陈波,潘青华,等.北京山地不同密度侧柏人工林枯落物及土壤水文效应[J].水土保持学报,2013,27(1):224-229.
[6] 卢振启,黄秋娴,杨新兵.河北雾灵山不同海拔油松人工林枯落物及土壤水文效应研究[J].水土保持学报,2014,28(1):113-116.
[7] 薛建辉,王智,吕祥生.林木根系与土壤环境相互作用研究综述[J].南京林业大学学报:自然科学版,2002,26(3):79-84.
[8] 喻理飞,朱守谦,祝小科,等.乌江流域不同植被—土壤系统涵养水源功能研究[J].贵州农学院丛刊,1991,17(1):36-52.
[9] 徐丽宏,王彦辉,熊伟,等.六盘山典型植被类型土壤水文生态功能研究[J].林业科学研究,2012,25(4):456-463.
[10] 肖卫平.茅台水源功能区植物群落地上结构持水功能评价研究[D].贵阳:贵州大学,2012.
[11] 黄威廉,屠玉麟,杨龙.贵州植被[M].贵阳:贵州人民出版社,1988.
[12] 林大仪.土壤学试验指导[M].北京:中国林业出版社,2004.
[13] 邓继峰,丁国栋,吴斌,等.宁夏盐池地区3种林分枯落物层和土壤水文效应[J].北京林业大学学报,2014,36(2):108-114.
[14] 高琛,鲁绍伟,杨新兵,等.北京松山不同密度丁香天然林枯落物及土壤水文效应[J].水土保持研究,2013,20(6):59-65.
[15] 吴迪,辛学兵,赵明扬,等.北京九龙山不同林分枯落物及土壤水文效应[J].林业科学研究,2014,27(3):417-422.
[16] Juliane D, Römermann C, Bernhardt-Römermann M, et al. Adaptation of plant functional group composition to management changes in calcareous grassland[J]. Agriculture Ecosystems & Environment,2011,145(1):29-37.
[17] 胡楠,范玉龙,丁圣彦.伏牛山森林生态系统灌木植物功能群分类[J].生态学报,2009,29(8):4017-4025.
Soil Water Regulation Function in the Rhizosphere of Main Tree Species in the Upper Reaches of Chishui River
YU Yanghua, LI Guangrong, YAN Lingbin, HUANG Zongsheng, YU Lifei
(CollegeofForestry,GuizhouUniversity,Guiyang550025,China)
In order to understand the soil water regulation ability in rhizosphere of main tree species in the upper reaches of Chishui River, through field sampling and experimental analysis method, study on ecohydrological function was carried out based on the bulk density, water content, porosity and other aspects, and the division of water regulation function groups. The results showed that the maximum water holding capacity of 32 species in 0—60 cm layer is about 212.00~357.94 mm, available water content ranged from 11.07 mm to 85.15 mm, was the reservoir; water holding rate was greater than the rate of water loss, and the relationship between the two and the time followed the power function model. Using RDA ordination, tree species in this area could be divided into 5 types of functional group. Functional group ⅠincludedMyricaceaerubra,Castanopsisfargesii,Camelliasinensis,Quercusaliena,PinusmassonianaandNeosinocalamusaffinis. Functional group Ⅱ includedPopulustrinervis,Cunninghamialanceolata,Broussonetiapapyifera,Viburnumchinshanense,Bambusapervariabilis,Liquidambaformosana,Pyracanthafortuneana,Linderaglauca,Sapiumsebiferum,MeliaazedarachandCoriarianepalensis. Functional group Ⅲ includedEriobotryabengalensis,Nandinadomestica,Prunusnalicina,Cupressusfunebris,PlatanusacerifoliaandToonasinensis. Functional group Ⅳ includedLoropetalumchinense,Vitexnegundo,Verniciafordii,MallotusbarbatusandChoerospondiasaxillaria. Functional group Ⅴ includedRhuschinensis,Pistaciachinensis,CitrusreticulataandCamelliaoleifera. Physical characteristics like soil bulk density, water content and porosity and water holding process are the key factors which affect the ability of regulation of soil hydrology. The results can provide theoretical basis for the selection and configuration of tree species in the water conservation forest of upper Chishui River.
tree rhizosphere soil; hydrological function; regulate soil water; functional group; upper reaches of Chishui River
2014-10-11
2014-10-22
贵州茅台科技联合资助项目“退化植被生态功能评价及生态修复技术与示范研究”(黔科合茅科联字[2009]7007);贵州省环境保护厅环境保护项目“赤水河上游退化植被水源涵养功能评价及恢复对策”(2013-09);贵州省农业攻关项目[黔科合NY(2013)3055号]
喻阳华(1984—),男,贵州仁怀人,博士生,研究方向为退化森林生态系统恢复。E-mail:yuyanghua2003@163.com
喻理飞(1963—),男,贵州贵阳人,教授,博士生导师,主要从事喀斯特退化生态系统恢复研究。E-mail:gdyulifei@163.com
S152.7; S715.7
1005-3409(2015)05-0284-05
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02
华南农业大学学报(2021年4期)2021-07-24
——美丽赤水河
南风(2021年14期)2021-06-13
煤田地质与勘探(2021年2期)2021-05-23
干旱地区农业研究(2019年3期)2019-07-18
天然产物研究与开发(2018年3期)2018-05-07
山东工业技术(2017年21期)2017-11-04
中国蔬菜(2016年8期)2017-01-15
遵义(2016年8期)2016-07-18
西藏大学学报(自然科学版)(2015年1期)2015-12-25
