
不同龄组杉木生态公益林空间结构
2015-03-23 09:21曹小玉李际平张彩彩房晓娜
浙江农林大学学报 2015年1期
曹小玉,李际平,封 尧,张彩彩,房晓娜
不同龄组杉木生态公益林空间结构
曹小玉,李际平,封 尧,张彩彩,房晓娜
(中南林业科技大学 林学院, 湖南 长沙 410004)
为了优化杉木Cunninghamia lanceolata生态公益林空间结构,充分发挥其生态、经济和文化等多项功能,以湖南省福寿林场杉木生态公益林9个固定样地林分空间结构的实测数据,采用混交度、大小比数和角尺度对比分析了不同龄组杉木生态公益林的空间结构特征。结果表明:杉木幼龄林的平均混交度为0.19,为0度混交和弱度混交的中间状态,杉木中龄林和成熟林的平均混交度为0.09和0.04,接近0度混交状态,3个龄组杉木林的林分混交度都偏低,林分稳定性差;杉木幼龄林、中龄林和成熟林的平均大小比数分别为0.51,0.52和0.52,接近中庸状态,说明林木个体差异不大,林木分化不严重;平均角尺度分别为0.37,0.35和0.34,为均匀分布和随机分布的中间状态,不是理想的林木水平分布格局。图4表4参14
森林经理学;杉木;生态公益林;空间结构
优化森林结构是培育健康稳定的多功能森林的重要途径,而森林空间结构作为森林结构最直接的表现,它不仅能够提供与林木空间位置有关的空间信息,并且决定林分非空间结构,是实现森林结构优化最有可能调控的因子[1]。因此,分析森林空间结构是优化森林结构的基础,对实现森林的多功能经营具有重要的意义[2]。空间结构单元的确定是林分空间结构分析的基础,林木空间结构单元是由1株参照木及其n株相邻木构成[3]。大多数学者采用邻近木取4的固定空间结构单元,如惠刚盈等认为n=4可满足混交林空间结构分析的要求。可在现实林分中,参照木主要与直接邻体产生竞争,固定空间结构单元的方法可能将直接邻体排除在外。如果采用Voronoi图确定林木的邻近木可以克服这个缺点。王峥峰等[4]、赵春燕等[5]基于Voronoi图确定林木空间结构单元进行林分空间结构分析,认为这种方法更为合理。杉木Cunninghamia lanceolata生态公益林作为中国南方公益林的重要组成部分,具有举足轻重的地位,但由于杉木林树种单一,生物多样性低,病虫害和火灾时有发生,导致其生态功能低下、经济效益产出也低,因此,如何通过森林空间结构的调整促其健康稳定的发展,已成为林业工作者需要迫切解决的问题[6]。本研究采用Voronoi图确定林分空间结构单元,选用角尺度、混交度、大小比数等3个空间结构指标对不同龄组杉木生态公益林空间结构进行分析,以期为杉木生态公益林结构的优化提供依据。
1 研究区概况
福寿林场位于湖南省平江县南部的福寿山上,地处28°3′00″~28°32′30″N,113°41′15″~113°45′00″E,处于中亚热带向北亚热带过渡的气候带,属湿润的大陆性季风气候。场内植被繁茂,群落较多,有木本植物55科275种。研究样地所属的杉木林均为在皆伐迹地上营造的杉木人工林,但它们经过抚育间和林分的自然更新,自然稀疏,自然灾害(虫害、火灾),人为破坏(盗伐)等导致林分分布密疏不均。
2 研究方法
2.1 数据调查方法
在杉木林有代表性的地段,用罗盘仪设置20 m×30 m的标准地9块(幼龄林、中龄林和成熟林各设置3块样地),将每块标准地都用相邻网格法划分为6个10 m×10 m小样方作为调查单元,以每个小样方的西南角为坐标原点,用皮尺测量每株树木在本小样方内的相对位置坐标(x,y),x为东西方向坐标,y为南北方向坐标,并记录小样方内的每株树木的树种、胸径、树高及冠幅,然后将样地西南角设为样地坐标系的原点。把每个小样方内林木的相对位置坐标转换为整个样地范围内同一坐标系内的坐标,以确定每株林木在整个样地内的相对位置分布,并将转换后的林木坐标数据导入ArcGIS软件中绘制林木空间位置分布图。每个标准地的基本情况见表1。
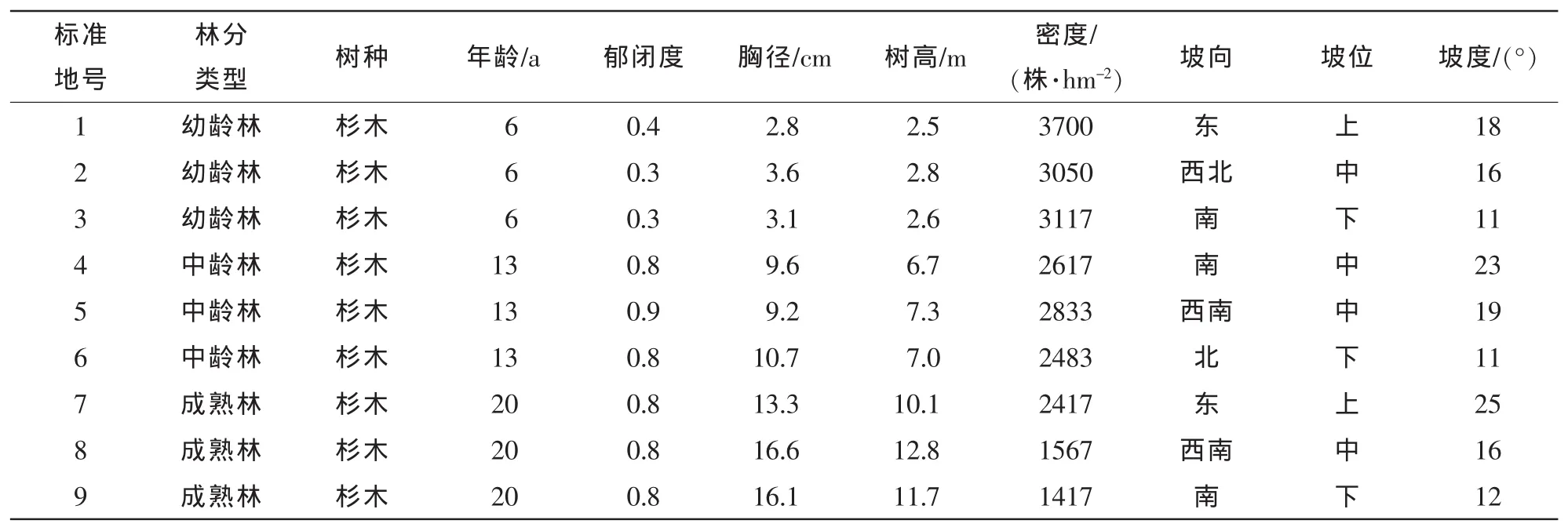
表1 调查样地基本情况表Table 1 Basic situation of sample plots
2.2 数据处理和分析方法
2.2.1 空间结构单元的确定 利用ArcGIS软件中创建泰森多边形工具,根据林木位置坐标数据生成Voronoi图。每个Voronoi多边形内只包含1株林木,其边数即为该林木的邻近木株数,其邻接Voronoi多边形所包含的林木即为该林木的邻近木具体分布位置,由此即可确定参照木的空间结构单元(图1)。
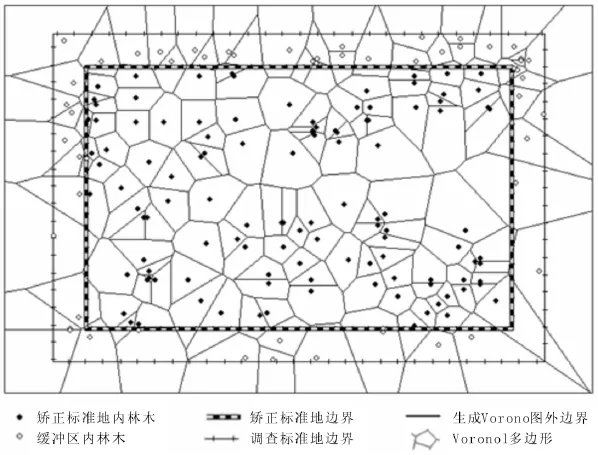
图1 基于林木点数据生成的加权Voronoi图Figure 1 Voronoi diagram based on tree data
2.2.2 边缘矫正 基于Voronoi图确定林分空间结构单元时处于样地边缘的边界木会受到边界的影响,且其邻近木可能处于样地外,故以边界木为中心木构建的空间结构单元是不完整的,会影响空间结构特征的分析结果。为了避免这种情况,必须对样地进行边缘矫正[7]。本研究采用距离缓冲区法,在原样地四周设置2.0 m宽的带状缓冲区。此宽度既可消除边界效应,又能充分利用样地内的调查数据。在缓冲区以外的林木为边缘木,只作为中心木的邻近木存在,而位于缓冲区内的林木均作为中心木参与计算。2.2.3 空间结构参数的计算 ①混交度。混交度是是描述树种间的相互隔离程度的参数[8-9],采用汤孟平等[10]提出的全混交度,描述树种间的相互隔离程度,计算公式为:
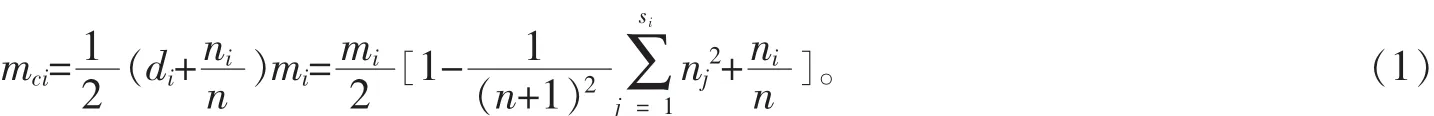
式(1)中:mci为中心木i点的全混交度;di为中心木i所在空间结构单元的Simpson指数;mi为中心木i点的简单混交度;n为邻近木株数;si为中心木i所在空间结构单元内的树种个数;nj为中心木i所在空间结构单元内第j树种的株数;ni为邻近木中不同树种的个数。计算林分或某一树种平均混交度时采用公式(2),其中N为林分或者某一树种中心木的株数。

②大小比数。大小比数是指冠幅、树高或者胸径大于做中心木的邻近木数占最近n株邻近木的比例(本研究采用胸径),用公式表示为[11]:

式(3)中:ui其取值为计算林分或某一树种的平均大小比数采用公式(4),其中N为林分或者某一树种中心木的株数。

③角尺度。角尺度是被定义为琢角(邻近木的较小夹角)小于标准角琢0的个数占所考察的 n个琢角的比例。它是描述空间结构单元内相邻木围绕中心木的水平分布分布格局[12],用下式表示如下:

式(5)中:wi取值为计算林分或某一树种的平均角尺度用式(6),式中N为林分或者某一树种中心木的株数。

3 结果与分析
3.1 混交度分析
从表2可以看出:杉木幼龄林不同样地的平均混交度为0.17~0.20,林分的平均混交度为0.19,为0度混交和弱度混交的中间状态,杉木中龄林不同样地的平均混交度为0.08~0.13,林分的平均混交度为0.09,接近0度混交状态,杉木成熟林不同样地的平均混交度为0.02~0.06,林分的平均混交度为0.04,接近0度混交状态,说明3个龄组杉木林林分混交程度都非常低,林分树种单一,物种多样性低,抗御病虫害、火灾等自然灾害的能力低,导致林分稳定性差。成熟林和中龄林混交度较幼龄林更低主要是由于成熟林和中龄林0度混交所占比例很大所导致的。在中龄林和成熟林中,0度混交比例高达43%和72%,说明成熟林和中龄林基本是相同树种的林木单种聚集在一起。同时杉木幼龄林、中龄林和成熟林的极强度混交比例都为0,说明在3个龄组的杉木林分中,没有任何一株林木完全被不同树种的邻近木所包围。
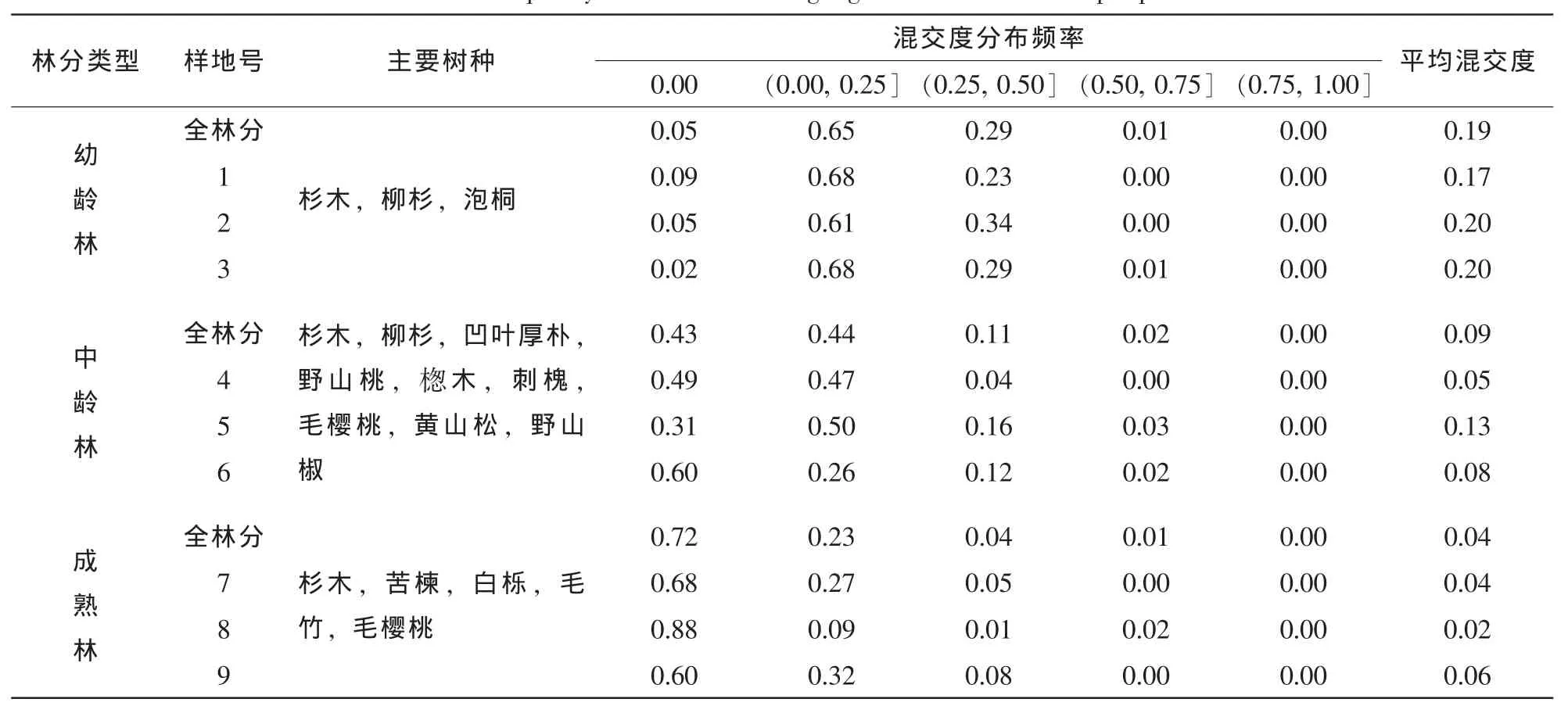
表2 林分及各样地混交度分布频率Table 2 Frequency distribution of mingling of the stand and sample plots
从不同树种看(图2),在杉木幼龄林、中龄林和成熟林林分中,杉木的平均混交度分别为0.17,0.06和0.02。为0度混交向弱度混交的过渡状态和0度混交状态。这说明杉木基本上是与同树种聚集生长在一起。在杉木幼龄林中,柳杉Cryptomeria fortunei的平均混交度为0.23,接近弱度混交状态,泡桐Paulownia的平均混交度为0.39。为弱度混交和中度混交的中间状态。在杉木中龄林林分中,柳杉,刺槐Robinia pseudoacacia,野山椒Capsicum frutescens的平均混交度分别为0.27,0.24和0.20,为弱度混交状态,野山桃Prunus davidiana的平均混交度为0.18,为0度混交向弱度混交的中间状态,黄山松Pinus taiwanensis,毛樱桃Cerasus tomentosa和凹叶厚朴Magnolia officinalis平均混交度分别为0.45,0.48,0.51为中度混交状态,楤木Aralia chinensis的平均混交度为0.65,为中度混交向强度混交过度状态。说明这8种树木在林分中没有出现单种聚集情况,均不同程度的与其他树种混交。在杉木成熟林中,苦楝Melia azedarach,毛竹Phyllostachys edulis和白栎 Quercus fabri的平均混交度分别为0.33,0.29和0.29,大致为弱度混交状态,毛樱桃的混交度为0.44,大致为中度混交状态。
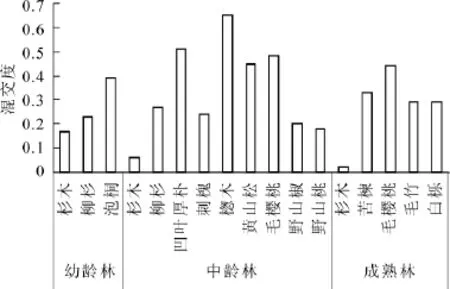
图2 不同树种平均混交度Figure 2 Average value of mingling degree of each tree species
3.2 大小比数分析
从表3可以看出:杉木幼龄林不同样地的平均大小比数为0.50~0.52,林分的平均大小比数为0.51,杉木中龄林不同样地的平均大小比数为0.48~0.52,林分的平均大小比数为0.52,杉木成熟林不同样地的平均大小比数为0.51~0.52,林分的平均大小比数为0.52,3个龄组的杉木林林分平均大小比数都接近中庸状态,说明3个龄组杉木林林分林木个体差异不大,林木分化不严重。

表3 林分及各样地大小比数分布频率Table 3 Frequency distribution of neighborhood comparison of the stand and sample plots
从不同树种看(图3):在杉木幼龄林、中龄林和成熟林林分中,杉木的平均大小比数分别为0.45,0.50和0.50。接近中庸状态。这说明杉木的胸径差异不明显。但杉木在不同龄组的林分中占的比例极大,其优势度非常明显。在杉木幼龄林中,柳杉的平均大小比数为0.60,为中庸和劣态的中间状态,说明柳杉在整个林分树木的竞争中处于相对劣势,泡桐的平均大小比数为0.28,接近亚优势的状态,说明泡桐在整个林分树木竞争中具有一定的优势。在杉木中龄林林分中,柳杉的平均大小比数为0.64,为中庸和劣态的中间状态,凹叶厚朴和野山椒的平均大小比数为0.80和0.83,为劣态和绝对劣态的中间状态,在与其他树种的竞争中处于不利地位。楤木的平均大小比数为1.00,为绝对劣态,在竞争中处于绝对劣势。刺槐的平均大小比数为0.20,为亚优势状态,说明其竞争优势明显。毛樱桃和野山桃的平均大小比数为0.46和0.57,接近中庸状态。黄山松的平均大小比数为0.34,为亚优势向中庸过渡状态。在杉木成熟林中,苦楝的平均大小比数为0.33,为亚优势向中庸过渡状态,毛樱桃的平均大小比数为0.6,为中庸向劣态过渡状态,毛竹的平均大小比数为0.78,为劣态,在整个林分树木的竞争中处于劣势。白栎的平均大小比数为1.00,为绝对劣态。
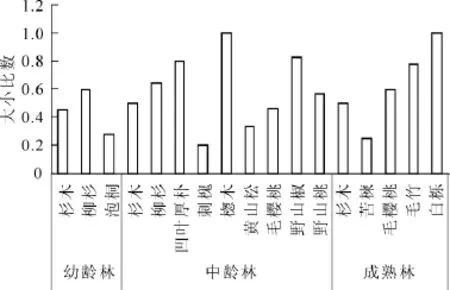
图3 不同树种平均大小比数Figure 3 Average value of neighborhood comparison of DBH of each tree species
3.3 角尺度分析
从表4可以看出:杉木幼龄林不同样地的平均角尺度为0.35~0.40,林分的平均角尺度为0.37,杉木中龄林不同样地平均混交度和林分的平均混交度为0.31~0.38,林分的平均角尺度为0.35,杉木成熟林不同样地的平均混交度为0.33~0.34,林分的平均混交度为0.34,说明3个龄组的杉木林林分平均角尺度都为均匀分布和随机分布的中间状态。需要通过结构调整是其由均匀分布向随机分布过渡。
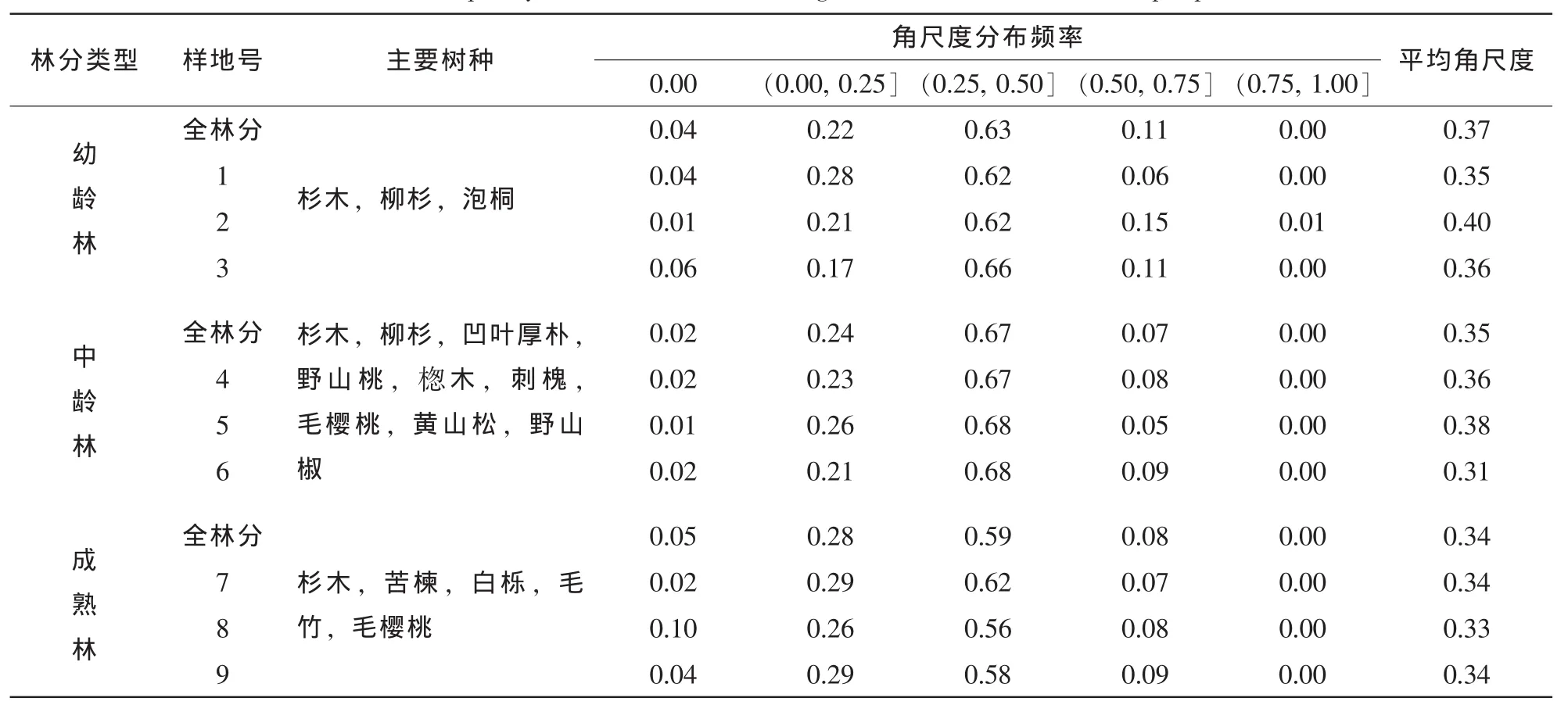
表4 林分及各样地角尺度分布频率Table 4 Frequency distribution of uniformangle index of the stand and sample plots
从不同树种看(图4):在杉木幼龄林、中龄林和成熟林林分中,杉木的平均角尺度为0.37,0.35和0.33。为均匀分布和随机分布的中间状态。这说明杉木的水平分布格局需要调整使其理想状态发展。在杉木幼龄林中,柳杉和泡桐的平均角尺度为0.37和0.40,为均匀分布和随机分布的中间状态。在杉木中龄林林分中,柳杉和黄山松的平均角尺度为0.37和0.34,为均匀分布和随机分布的中间状态,毛樱桃和野山椒的平均角尺度为0.50,为随机分布状态,是一种比较理想的分布状态。凹叶厚朴和野山桃的平均角尺度为0.40和0.43,为均匀分布和随机分布的中间状态,楤木平均角尺度为0,为绝对匀分布,刺槐的平均角尺度为0.20,接近均匀分布状态。在杉木成熟林中,白栎的平均角尺度为0.50,为随机分布状态,毛樱桃、苦楝和毛竹的平均混交度分别都接近0.35,为均匀分布和随机分布的中间状态。
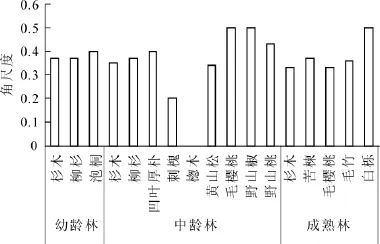
图4 不同树种的平均角尺度Figure 4 Average value of uniform angle index of each tree species
4 结论与讨论
本研究采用混交度、大小比数和角尺度分析了不同龄组杉木生态公益林的空间结构特征。结果表明:杉木幼龄林的平均混交度为0.19,为0度混交和弱度混交的中间状态,杉木中龄林和成熟林的平均混交度为0.09和0.04,都接近0度混交状态,3个龄组的林分混交程度都很低,林分稳定性差,需要通过引进阔叶树种,增加林分的混交度;杉木幼龄林、中龄林和成熟林的平均大小比数分别为0.51,0.52 和0.52,接近中庸状态,说明林木个体差异不大,林木分化不严重;平均角尺度分别为0.37,0.35和0.34,为均匀分布和随机分布的中间状态,不是理想的水平分布格局。
采用n=4构成的空间结构单元来分析林分的空间结构特征,这一标准被很多学者采用,但Pommerening[13]认为不同林分状况下确定的林分空间结构单元大小应该是不同的,Voronoi图能根据林分的实际情况,灵活确定各空间结构的单元的大小。这在一定程度上克服了固定空间结构单元所导致的有偏估计缺陷[14]。基于此,本研究采用Voronoi图确定林分的空间结构单元。但是当利用Voronoi图来确定空间结构单元时却没有计算角尺度的相应理论标准,致使不同研究人员在采用基于Voronoi图计算角尺度时,因没有统一的标准而导致研究结果相差较大。因此,如何基于Voronoi图计算角尺度的新的评判标准,是进行角尺度研究的一个重要方向。
简单混交度分析了不同龄组的杉木生态公益林林分内不同树种间的隔离程度,仅考虑了中心木与4株邻近木之间的树种异同关系,未曾考虑4株邻近木彼此之间的树种异同。因此,需要提出一个全新的混交度指标来代替它。本研究采用汤孟平等[10]提出的全混交度来分析杉木生态公益林的混交程度,它不仅考虑中心木与邻近木之间以及邻近木彼此之间的树种隔离关系,还引入了生物多样性Simpson指数来描述树种多样性,以提高树种多样性的区分度,是进行混交度分析比较理想的指标。
[1] NORTH M,CHEN Jiquan,OAKLEY B,et al.Forest stand structure and pattern of old-growth western hemlock/Douglas-fir and mixed-conifer forests[J].For Sci,2004,50(3):299-311.
[2] MASON W L,CONNOLLY T,POMMERENING A,et al.Spatial structure of semi-natural and plantation stands of Scots pine in northern Scotland[J].Forest,2007,80(5):567-586.
[3] 汪平,贾黎明,魏松坡,等.基于 Voronoi图的侧柏游憩林空间结构分析[J].北京林业大学学报,2013,35(2):39-44.WANG Ping JIA Liming,WEI Songpo,et al.Analysis of stand spatial structure of Platycladus orientalis recreational forest based on Voronoi diagram method[J].J Beijing For Univ,2013,35(2):39-44.
[4] 王峥峰,安树青,朱学雷,等.热带森林乔木种群分布格局及其研究方法的比较[J].应用生态学报,1998,9 (6):575-580. WANG Zhengfeng,AN Shuqing,ZHU Xuelei,et al.Distribution pattern of tree populations in tropical forest and comparison of its study method[J].Chin J Appl Ecol,1998,9(6):575-580.
[5] 赵春燕,李际平,李建军.基于Voronoi图和Delaunay三角网的林分空间结构量化分析[J].林业科学,2010,46(6):78-84. ZHAO Chunyan,LI Jiping,LI Jianjun.Quantitative analysis of forest stand spatial structure based on Voronoi diagram &Delaunay triangulated network[J].Sci Silv Sin,2010,46(6):78-84.
[6] 汤孟平.森林空间结构研究现状与发展趋势[J].林业科学,2010,46(1):117-122. TANG Mengping.Advances in study of forest spatial structure[J].Sci Silv Sin,2010,46(1):117-122.
[7] 周红敏,惠刚盈,赵中华,等.林分空间结构分析中样地边界木的处理方法[J].林业科学,2009,45(2):1-5. ZHOU Hongmin,HUI Gangying,ZHAO Zhonghua,et al.Treatment methods of plot boundary trees in spatial forest structure analysis[J].Sc Silv Sin,2009,45(2):1-5.
[8] 惠刚盈,胡艳波.混交林树种空间隔离程度表达方式的研究[J].林业科学研究,2001,14(1):23-27. HUI Gangying,HU Yanbo.Measuring Species spatial isolation in mixed forests[J].For Res,2001,14(1):23-27.
[9] GADOW K V,FÜLDNER K.Bestandesbeschreibung in der forsteinrichtung tagungsbericht der arbeitsgruppe forsteinrichtung klieken bei dessau[J].Forest,1992,30(5):267-286.
[10] 汤孟平,娄明华,陈永刚,等.不同混交度指数的比较分析[J].林业科学,2012,48(8):46-53. TANG Mengping,LOU Minghua,CHEN Yonggang,et al.Comparative analysis on different mingling indices[J].Sci Silv Sin,2012,48(8):46-53.
[11] 惠刚盈,von GADOW K,ALBERT M.一个新的林分空间结构参数——大小比数[J].林业科学研究,1999,12(1):1-6. HUI Gangying,von GADOW K,ALBERT M.A new parameter for stand spatial structure neighbourhood comparison[J].For Res,1999,12(1):1-6.
[12] 惠刚盈.角尺度:一个描述林木个体分布格局的结构参数[J].林业科学,1999,35(1):37-42. HUI Gangying.The neighbourhood pattern:a new structure parameter for describing distribution of forest tree position[J].Sci Silv Sin,1999,35(1):37-42.
[13] POMMERENING A.Evaluating structural indices by reversing forest structural analysis[J].For Ecol Manage,2006, 224(3):266-277.
[14] 汤孟平,陈永刚,施拥军,等.基于Voronoi图的群落优势树种种内种间竞争[J].生态学报,2007,27(11):4707-4716. TANG Mengping,CHEN Yonggang,SHI Yongjun,et al.Intraspecific and interspecific competition analysis of community dominant plant populations based on Voronoi diagram[J].Acta Ecol Sin,2007,27(11):4707-4716.
Stand spatial structure for an uneven-aged Cunninghamia lanceolata ecological forest
CAO Xiaoyu,LI Jiping,FENG Yao,ZHANG Caicai,FANG Xiaona
(School of Forestry,Central South University of Forestry&Technology,Changsha 410004,Hunan,China)
To optimize the spatial structure of a Cunninghamia lanceolata ecological forest and use of its ecological,economic,and cultural functions,nine permanent sample plots(20 m×30 m)at the Fushou Forest Farm in Hunan Province were analyzed for variation in spatial structure of uneven age-group stands using three spatial structure indexes:the mingling index,the neighborhood comparison,and the uniform angle index. Results showed an average mingling degree for juveniles of 0.19 which belonged to the transitional type between zero and weak mingling.The average degree of mingling for medium(0.09)and mature(0.04)trees was close to zero mingling.The average neighborhood comparison for stand DBH was juvenile (0.51),medium (0.52),and mature trees (0.52)which were close to the moderate state.The average stand uniform angle indexes were juvenile(0.37),medium(0.35),and mature(0.34)belonging to the transitional types of uniform and random distributions.Overall for the C.lanceolata stands,mingling showed that the ecological forest rating was low,and the stability of the stand structure was weak;the neighborhood comparison indicated that the number of trees in diameter classes was roughly equal;and the uniform angle index revealed poor horizontal distribution patterns in juvenile,medium,and mature stands.[Ch,4 fig.4 tab.14 ref.]
forest management;Cunninghamia lanceolata;ecological forest;spatial structure
S757.1
A
2095-0756(2015)01-0084-08
浙 江 农 林 大 学 学 报,2015,32(1):84-91
Journal of Zhejiang A&F University
10.11833/j.issn.2095-0756.2015.01.012
2014-04-29;
2014-08-22
“十二五”国家科技支撑计划资助项目(2012BAD22B0505);湖南省教育厅资助项目(12C0440)
曹小玉,讲师,博士研究生,从事森林经理等研究。E-mail:cxy7723@aliyun.com。通信作者:李际平,教授,博士生导师,从事森林经理等研究。E-mail:lijiping@vip.163.com
猜你喜欢
林业勘查设计(2021年2期)2021-04-08
锦绣·下旬刊(2020年12期)2020-01-03
农业与技术(2018年16期)2018-11-28
农村经济与科技(2018年15期)2018-05-14
森林工程(2018年5期)2018-05-14
现代农业科技(2017年10期)2017-07-12
湖北林业科技(2016年3期)2016-08-10
