
未来黄、东海营养盐浓度变化情景预测*
2015-03-22 00:58赵一丁
海洋与湖沼 2015年5期
赵一丁 杨 波 ① 魏 皓 赵 亮
(1. 中国海洋大学海洋环境学院 青岛 266100; 2. 天津科技大学海洋科学与工程学院 天津 300457)
联合国政府间气候变化专门委员会(Intergovernmental Panel on Climate Change, IPCC)第四次评估报告(IPCC,2007)通过一系列耦合环流模式对未来温室气体增加的不同情景下全球气候变化作了预测, 其结果显示未来全球气候变暖趋势仍会持续, 升温带来的生物过程变化会影响海水中营养盐的分布。河流物质通量的研究显示, 由于农业生产(Bouwmanetal, 2010)、污水排放(Van Drechtetal, 2009)和水文气候(Feketeet al, 2010)变化, 未来全球河流无机氮、无机磷排放量将显著增加, 河流营养盐结构随之发生变化(Garnieretal, 2010; Seitzingeretal, 2010)。在气候和河流营养盐排放变化的影响下, 未来海水营养盐浓度发生怎样的变化?营养盐的分布与变化对生物生产力和生物资源量的变动有何影响?这些是当前生态环境研究共同关心的热点问题。
黄、东海是我国重要的陆架海区, 以生产力高而著称, 是我国经济社会发展的重要支撑系统之一。过去几十年, 黄、东海水温有明显上升趋势(袁承仪,2011; 李家星等, 2012)。随着经济的发展和人类活动加剧, 河流携带入海的陆源物质显著增加, 生态环境变化受到气候变化和人类活动的双力推进, 黄、东海生态系统面临的压力骤增, 生态灾害频发(唐启升等, 2001)。针对未来黄、东海营养盐浓度变化趋势的预测是未来生态环境变化研究的重要组成部分,也可为国家制定海洋生态环境保护和可持续发展政策提供参考。为此, 我们采用降尺度(downscaling)方法, 利用气候模式给出的气候预测结果来驱动区域海洋水动力模式, 得到未来黄、东海流场、湍流混合等水动力条件, 结合预测得到的未来河流营养盐载荷, 再与生态系统模型进行耦合, 预测未来黄、东海生态环境状态。本文集中讨论营养盐的预测结果。
1 模式与方法
本文采用的三维物理-生物耦合模型包括水动力模块和生态模块两个部分(Zhaoetal, 2011)。水动力模块为海洋环流模式POM (Princeton Ocean Model),空间分辨率为 1/18°, 垂向采用σ坐标共分为 21层,该模块为生态模块提供水温、环流、湍流混合等条件。生态模块基于海洋生态动力学模式 NORWECOM(Aksnesetal, 1995; Skogenetal, 1995), 包括三类营养盐(溶解无机氮 DIN、溶解无机磷 DIP和硅酸盐SIL)、两类浮游植物(硅藻(DIA)和鞭毛藻FLA)和两类生物有机物(碎屑DET和生物硅SIS)。该模块包含了浮游植物对营养盐的吸收、呼吸释放以及水体矿化再生过程, 底边界考虑了碎屑的沉降和再悬浮过程。由于底营养盐再生涉及过程和参数较复杂, 以往研究中并没有给出明确的底通量, 且本文主要关心河流的影响, 因此, 未考虑无机营养盐的底再生过程。模型边界条件、初始条件及生态参数的设定可参见Zhao等(2011)。营养盐、叶绿素a周年变化模拟已与观测数据进行对比, 校验结果表明此生态模型可以反映黄、东海营养盐分布及变化的规律。
本文针对 IPCC报告中提出的四个未来情景, 选择大气温室气体增长的中端稳定路径RCP4.5情景(到2100年辐射强迫稳定在 4.5W/m2左右)作为对未来气候变化的预估情景。我们从 IPCC报告提出的海气耦合模式中选取FGOALS_s2.0模式(the second spectral version of the Flexible Global Ocean-Atmosphere-Land System model)输出的未来(2026—2075年)和现代(1951—2000年)平均的风场及热通量结果作为强迫场来驱动水动力模式POM及生态模块。
本研究的海区营养盐初始条件、黑潮开边界条件和大气干湿沉降条件等设定与 Zhao等(2011)相同,研究中仅改变未来营养盐河流输入量。基于千年生态系统评估(Millennium Ecosystem Assessment, MEA)对未来社会经济发展规划的四个情景(Alcamo, 2006),Qu等(2010)以及Strokal等(2014), 应用GlobalNEWS模式(Global Nutrient Export from Watersheds)预测并分析了未来中国河流营养盐排放, 我们选取其中两个极端情景(GO和AM), 根据2050年相对2000年河流排放无机氮、无机磷增加的比例, 计算未来各河流营养盐的浓度作为未来河流输入边界条件。未来径流量变化很小(Yangetal, 2005), 控制模式中10条河流流量不变(海洋图集编委会, 1993)。由于硅酸盐的保守性, 未来河流硅酸盐浓度变化不大, 且主要受筑坝节流的影响, 模型中设其保持不变。2000年河流营养盐浓度数值参考 Zhou等(2008)及 Liu等(2009)的研究。模式由初始状态计算两年之后基本稳定, 本文采用第三年输出结果进行分析。
本文将黄、东海分区统计, 浮游植物的环境因子主要为温度、盐度、营养盐和光照等, 其中河流营养盐随淡水排放, 与盐度有较好的相关性。利用模式现代(2000年)的年均表层和底层温盐数据, 按水团分析方法中的聚类分析(图1及表1)将研究区域划分为黄海沿岸、黄海中部、长江口邻近海区和东海陆架区四个区域(图2)。黄海沿岸海区直接受河流排放影响, 营养盐浓度较高, 水深较浅, 属于黄海沿岸水系。黄海中部海区的突出特征为夏季存在冷水团, 该海区温跃层强,层结稳定, 生态环境独特, 营养盐来自邻近海区的输运。长江口邻近海区主要受长江冲淡水及苏北沿岸水的影响, 有充足的营养盐补充; 东海陆架区受台湾暖流及黑潮的影响, 呈现高温高盐的水文特征。本文针对未来气候变化和河流营养盐排放变化的影响, 研究各海区未来营养盐的分布与变化并探讨其原因。

表1 黄、东海各海区温盐特征Tab.1 The characteristics of temperature and salinity in the Yellow Sea and East China Sea
2 结果
2.1 未来水动力场的变化
张维娟等(2014)采用FGOALS模式的未来(2026—2075年)和现代(1951—2000年)的风场及热通量结果作为强迫场来驱动水动力模式 POM, 并将其得到的现代的结果与《渤黄、东海水文图集》(海洋图集编委会, 1993)进行了对比校验, 结果表明POM模式对现代水文状态的模拟基本符合观测事实。本研究采用张维娟等(2014)未来水动力场的预测结果作为未来水动力条件。

图1 黄、东海表(左)底层(右)温盐聚类分析Fig.1 Cluster analysis based on temperature and salinity data at the surface (left panel) and bottom (right panel) layers in the Yellow Sea and East China Sea

图2 黄、东海地形及生态分区, 紫色三角表示模式中包括的10条河流Fig.2 Topography and ecological division in the Yellow Sea and East China Sea. Ten rivers included in the model are denoted by purple triangles
对比未来与现代的黄、东海水动力环境, 发现在RCP4.5情景下, 2050年黄、东海年均水温相比现代高2.1 C, 而黄海冷水团受冬季水温升高的影响, 冷水团面积减少, 核心温度升温幅度为 1.9 C。随着冬季风的减弱, 具有补偿性质的黄海暖流也相应减弱, 同时表层流和沿岸流也减弱。具体分析详见张维娟等(2014)。
2.2 未来黄、东海入海河流营养盐载荷
近 50年来, 中国河流中的无机氮、无机磷载荷迅速增加(Wang, 2006; Lietal, 2007), 未来该趋势还将持续。利用 GlobalNEWS模式(Quetal, 2010;Strokaletal, 2014)的结果计算得到的2050年GO、AM两个情景下中国河流营养盐浓度见表2。由浓度与河流径流量的乘积得到营养盐入海通量即载荷。
GO情景下, 流入渤海的河流(黄河、辽河、滦河、海河)无机氮和无机磷营养盐载荷分别为现代的 1.34倍和 2.71倍, 而流入黄、东海的河流(长江、闽江、鸭绿江、淮河、汉江、钱塘江)无机氮和无机磷营养盐载荷分别为现代的1.58倍和1.60倍。
AM情景下, 流入渤海的河流(黄河、辽河、滦河、海河)无机氮和无机磷营养盐载荷分别为现代的 1.06倍和 3.31倍, 而流入黄、东海的河流(长江、闽江、鸭绿江、淮河、汉江、钱塘江)无机氮和无机磷营养盐载荷分别为现代的1.13倍和2.83倍。相比GO情景, AM 情景下无机氮增长减缓, 无机磷增长则更为迅速, 但是没有改变大部分河流无机氮过剩的状况。
营养盐入海通量与径流量密不可分。由于长江径流量巨大, 其每年携带入海的物质通量是最大的。长江无机氮、无机磷输入均占河流输入总量的80%。河流携带入海的营养盐比例差异也很大, GO情景下十条河流营养盐均为表现为氮过剩, 而在AM情景下淮河表现为磷过剩。本文仅讨论该营养盐载荷下的未来黄、东海营养盐分布变化。河流的独特营养盐结构会影响其邻近海域浮游植物的群落组成和生物过程,这将在未来的研究中深入讨论。
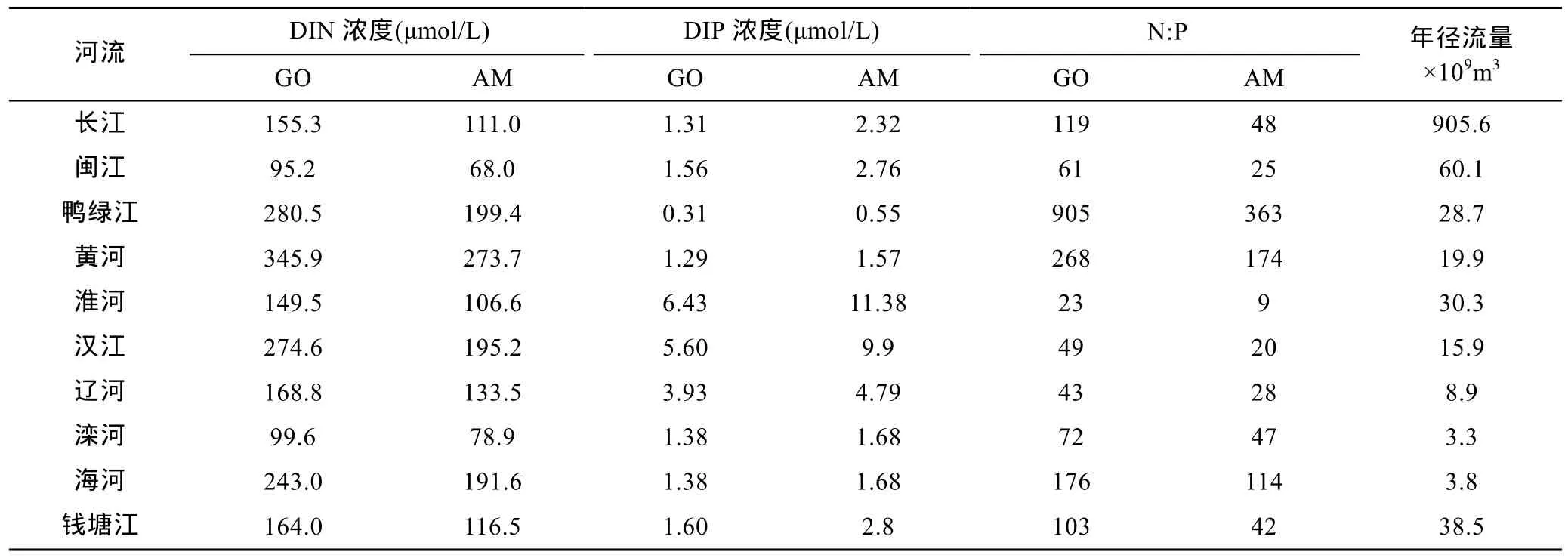
表2 2050年黄、东海入海河流营养盐浓度、径流量及氮磷比预测Tab.2 Prediction of nutrient concentration, runoff and N/P ratio export by rivers to the Yellow Sea and East China Sea in 2050
2.3 未来营养盐分布与变化的情景预测
本文利用以上水动力场和河流营养盐载荷数据,模拟现代和预测未来不同情景下无机氮、无机磷营养盐浓度水平分布及变化情况。
2.3.1 无机氮浓度的分布与变化 未来两个情景下无机氮浓度分布情况与现代情景相似, 这里仅列出各情景与现代的差值。
现代情景下, 表层无机氮浓度在河流邻近海域冬、夏季均为高值(图 3), 可达 40μmol/L, 但是其高值随着河流羽流扩展范围不同。冬季 DIN浓度较高的海水随沿岸流到达台湾海峡西岸, 夏季则随冲淡水向东北扩展。东海中陆架冬、夏季 DIN均小于2μmol/L。相比现代, GO情景下冬季水深小于30m的近岸海区表层DIN浓度增长迅速, 其在汉江口、鸭绿江口、淮河口、长江-钱塘江口的增量可达 10μmol/L,比现代浓度增加约 20%, 增量的高值随低盐水在近岸扩展; 夏季受冲淡水扩展的影响, 黄海中部的DIN浓度比现代高5μmol/L。AM情景下冬季30m以浅近岸海区表层DIN浓度增长缓慢约为2μmol/L, 黄海中部的DIN浓度与现代相比增加和减小的区域呈斑块状分布; 夏季比较显著的特征是黄海中部DIN浓度减小2μmol/L。与现代情景相比, 东海中陆架DIN浓度变化较小。
现代情景下, 河流在底层影响区域减小, 高 DIN浓度仅出现在河口范围内和沿岸流区, 羽流区外底层 DIN浓度大于表层。黄海中部和济州岛以西海域底层DIN浓度较高, 这可能与黄海暖流补充、九州岛西南黑潮向陆架入侵(Zhaoetal, 2011)及夏季底层矿化再生过程有关。底层另一个突出现象是黑潮区域的DIN浓度较高(>30μmol/L)。另外, 东海中陆架夏季底层 DIN 浓度(>5μmol/L)高于台湾海峡(<2μmol/L), 这与携带高营养盐的黑潮次表层水入侵至东海陆架有关(Guoetal, 2006; Yangetal, 2011)。夏季黑潮次表层水分为内外分支时, 在长江口以东内外分支之间形成了一个DIN浓度较低的孤立水体。相比现代情景,GO情景底层 DIN浓度增加的区域与表层基本一致,而AM情景下表层和底层DIN浓度变化明显不同。夏季除河口近岸地区DIN浓度增加外, 黄海中部DIN浓度也有所增加(2—5μmol/L), 位于黄海中部西侧,与表层DIN减少区域一致, 冬季该区域依然存在DIN浓度增加的现象。
长江口邻近海区及东海陆架区的无机氮分布情况如31.5°N断面分布图(图4)所示。东海内陆架(水深<50m)DIN浓度高于 20μmol/L的区域主要受到长江羽流的影响, 冬季贴近岸边, 夏季则随着羽流北转。长江浅滩上冬季 DIN浓度较高主要是因为受到黄海沿岸流的影响, 夏季的低 DIN浓度则与浮游植物的消耗有关。陆架区清楚地显示了底层黑潮次表层水的涌升和跃层附近营养盐的消耗。GO情景下, 冬季50m 以浅 DIN浓度上升幅度均大于 2μmol/L, 夏季DIN浓度的增加集中体现在羽流区。AM情景下DIN浓度变化小于GO情景。
从 35°N断面来看, 夏季黄海上层受西岸淮河、长江输入的影响, DIN浓度较高; 次表层浮游植物生长消耗了营养盐, 所以跃层以下DIN浓度低于上层。冬季垂向混合过程把沿岸河流输入的营养盐带入深层, DIN垂向分布均匀。未来两个情景中DIN浓度变化存在显著差异。GO情景中, 夏季黄海上层的 DIN浓度上升约5μmol/L, AM情景中DIN的浓度则下降1—4μmol/L。两种情景下, 黄海底层的DIN浓度均有所增加, AM情景增幅大于GO情景。

图3 现代情景下表(2m)底(σ=20)DIN以及未来情景的相对变化ΔDIN的水平分布(单位: μmol/L), 从上到下依次为现代情景、GO情景与现代情景之差以及AM情景与现代情景之差, 下同Fig.3 Horizontal distribution of modern DIN (μmol/L) and the relative change between the future and modern time at surface (2m) and bottom (σ=20) layers Panels from top to bottom: the modern scenario, the scenario with the change of GO, and the scenario with the change of AM

图4 现代情景下31.5°N断面(左)、35°N断面(右)DIN浓度以及未来情景的DIN浓度相对变化ΔDIN的垂向分布(单位:μmol/L), 红线表示真光层深度Fig.4 Vertical distribution of modern DIN (μmol/L) and the relative change in the future along the section at 31.5°N (left panel) and 35°N (right panel). Red lines denote euphotic depth
2.3.2 无机磷浓度分布与变化 现代黄、东海表层无机磷浓度分布(图 5)表明, 冬季淮河、长江-钱塘江影响区及黄海中部均保持较高的 DIP浓度(>0.5μmol/L), 东海中陆架 DIP浓度范围为 0—0.2μmol/L。夏季黄海中部DIP消耗殆尽, 仅长江口外近岸区DIP浓度较高。由于AM情景中河流DIP载荷的增幅大于GO情景, AM情景中高值区扩展范围大于GO情景。
底层无机磷浓度分布特征与无机氮相似: 近岸河口区、黑潮区、黄海中部浓度较高(>0.5μmol/L)且非羽流区底层浓度大于表层, 东海陆架黑潮次表层水的影响范围(>0.5μmol/L)更加清晰。AM情景夏季黄海中部底层 DIP浓度同样有所增加(增加0.2μmol/L), 位置与底层DIN浓度升高区域一致。断面分布(图 6)显示了黄海中部上层 DIP全部被消耗,并且从图中可以看出长江、黑潮次表层水携带的高磷水的影响。两个情景DIP浓度分布相似, 但是AM情景黄海中部底层DIP增幅大于GO情景。
3 讨论
3.1 未来营养盐平均浓度与结构
本文将黄、东海划分为黄海沿岸、黄海中部、长江口邻近海区和东海陆架区(图 2), 统计两个情景下各海区未来氮、磷营养盐平均浓度预测值(表3)。
GO情景下, 黄海沿岸DIN和DIP的浓度分别增长30.6%和7.0%, 黄海中部分别增长29.8%和10.8%,长江口邻近海区则分别增长39.2%和11.7%。由于本文未考虑未来情景下开边界处黑潮和台湾暖流的变化, 东海陆架区DIN和DIP的浓度变化很小。虽然河流无机氮、无机磷载荷增长幅度较大, 但是海水的缓冲能力强, 加之生物过程消耗, 三个海区的营养盐浓度平均仅增加33%(DIN)和10%(DIP), 无机氮浓度增幅大于无机磷, 各海区磷限制更加显著。长江口邻近海区不断输入的大量营养盐未被充分消耗, 所以增幅大于其他海域。
AM情景下, 黄海沿岸的DIN和DIP浓度分别增长6.3%和21.1%, 黄海中部的DIN和DIP浓度分别增长 8.4%和 23.1%, 长江口邻近海区的 DIN和 DIP浓度分别增长9.3%和36.4%。三个海区的营养盐浓度总体平均增加 8%(DIN)和 27%(DIP), 无机磷浓度增幅大于无机氮。由于AM情景下河流无机磷载荷增长迅速, 黄海沿岸的氮磷比由25.6下降至22.5, 黄海中部由22.6下降至19.9, 长江口邻近海区由34.4下降至24.8。由此可见, 该情景下整个海区的营养盐结构得到调整, 各海区氮过剩的状况也有所减轻。然而由于生物可利用营养盐总浓度的增加, 海区富营养化程度进一步加强。

图5 现代情景下表层(2m)和底层(σ=20)的DIP浓度分布情况以及未来情景的相对变化ΔDIP的水平分布(单位: μmol/L), 从上到下依次为现代情景、GO情景下的变化、AM情景下的变化Fig.5 Horizontal distribution of modern DIP (μmol/L) and the relative change in the future at surface (2m) and bottom (σ=20) layers Panels from top to bottom: the modern scenario, the scenario with the change of GO, and the scenario with the change of AM
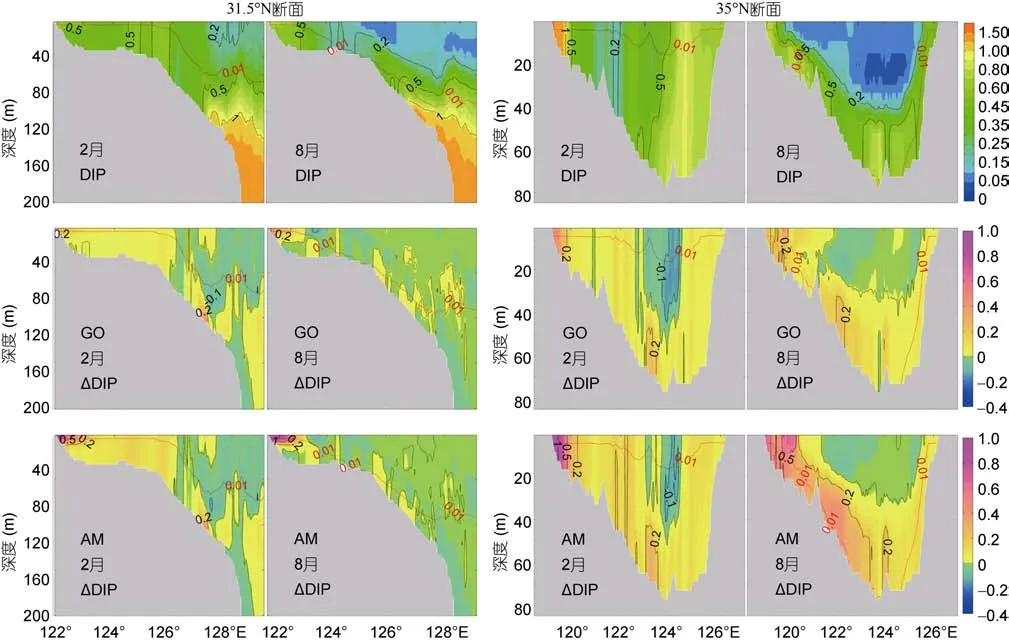
图6 现代情景下31.5°N断面(左)、35°N断面(右)DIP以及未来情景的相对变化ΔDIP的垂向分布(单位: μmol/L)Fig.6 Vertical distribution of modern DIP (μmol/L) and the relative change in future along the section at 31.5°N (left panel) and 35°N(right panel)
3.2 水动力环境对未来营养盐变化的贡献
我们通过一个敏感性实验来讨论在上述未来营养盐浓度改变过程中水动力环境与河流营养盐输入在不同海区的作用。该敏感性实验中生态模式仅采用未来(2026—2075年)水动力条件驱动, 河流营养盐输入仍然采用现代(2000年)浓度, 其他开边界条件不变。
实验结果与现代模拟的对比见表3、表 4。敏感性实验中各海区营养盐冬、夏季的分布与现代情景类似, 但营养盐浓度略有增加。黄海沿岸 DIN和 DIP浓度分别增长1.3%和1.8%, 黄海中部分别增长4.1%和4.6%, 长江口邻近海区分别增长1.5%和1.3%。由此可见, 相比 GO、AM 情景, 在河流影响的沿岸区,仅改变水动力环境的情况比同时考虑未来河流载荷增加的情况增幅小很多, 黄海中部水动力条件的改变对营养盐浓度增长有一定贡献。因此, 未来河流营养盐排放量的增长是未来沿岸海区营养盐浓度增加的主要原因。尽管黄海中部没有河流的直接注入, 但其营养盐的变化仍然主要来自于河流输入导致的周围环境浓度变化。
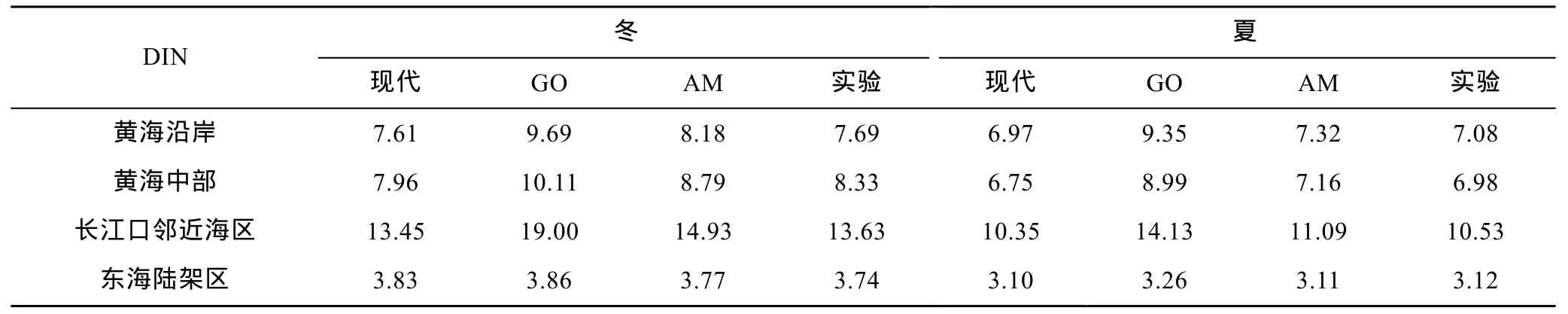
表3 各海区冬、夏季无机氮浓度统计结果(单位: μmol/L)Tab.3 Statistical results of DIN concentration (μmol/L) in each box in winter and summer
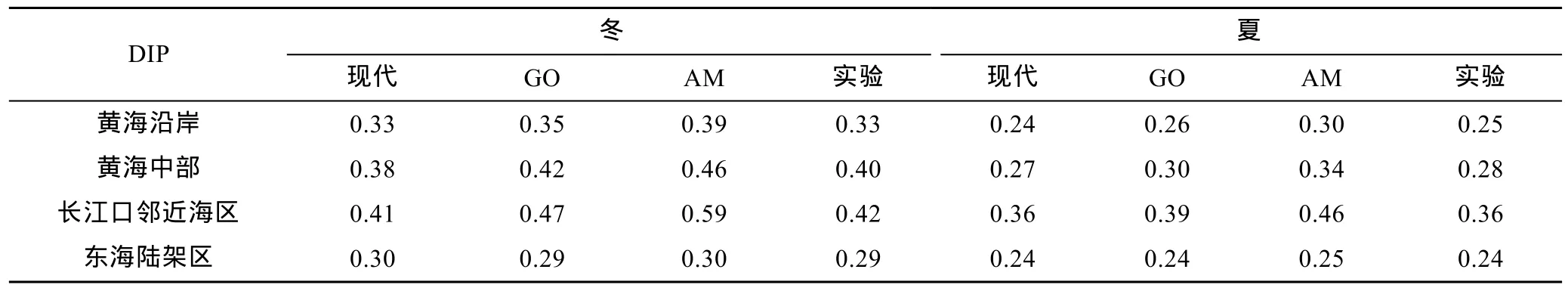
表4 各海区冬、夏季无机磷浓度统计结果(单位: μmol/L)Tab.4 Statistical results of DIP concentration (μmol/L) in each box in winter and summer
4 未来黄、东海营养盐收支
营养盐浓度是非保守的, 既受外界输运影响, 又存在其内部循环过程。对不同海区营养盐收支进行分析, 可以帮助我们更好地了解导致其未来变化的主要因素。
根据 Moll(1998)和赵亮等(2002)构建的氮、磷循环模型, 营养盐循环概念模型可概括为: 浮游植物在其生命过程中通过光合作用和呼吸作用分别吸收和释放营养盐, 其死亡后在水体及水底形成碎屑, 水底碎屑通过沉降与再悬浮与水体进行交换, 再通过矿化过程补充水体中的营养盐。
根据物质守恒, 有:

积分右端各项可得到浮游植物光合作用吸收、呼吸作用释放、水体矿化等营养盐内部循环过程, 以及水平对流输运、垂直扩散和河流输入等外部循环过程对各海区营养盐收支的贡献, 其中光合作用吸收与呼吸释放之差就是浮游植物的净初级生产量。
由各区域全年无机氮营养盐收支情况(表5)可见,GO、AM 两个情景下光合作用吸收、呼吸释放、水体矿化以及河流输入总量(有河流注入的区域)均有所增加。对流输运量除东海陆架区对流输入量减少外,其他各区域均呈现增加的趋势。营养盐含量的升高导致了生物过程的增强, 此时主要通过内部循环过程的消耗来达到平衡, 且在AM情景下无机磷增长较快,缓解了海区磷限制, 促进了氮的消耗, 因此生物过程较GO情景更强。

表5 各海区物理过程和生物过程无机氮收支的贡献(单位: ×103t/a)Tab.5 Contribution of physical and biological process to the budgets of DIN (×103t/a) in each box
黄海沿岸区, GO和AM两个情景下无机氮河流输入分别增加91.0×103t/a和20.1×103t/a, 对流输入分别增加 54.5×103t/a和 131.7×103t/a, 两者共同导致了该区域无机氮浓度的增加, 由此净初级生产分别增长13%和31%。由于AM情景生物过程较强, 夏季无机氮被大量消耗, 该海区接收的无机氮输运高于 GO情景。黄海中部没有河流直接输入, 依靠对流输运补充无机氮, GO和 AM 情景对流输入分别增加 92.7×103t/a和 37.2×103t/a。夏季长江冲淡水向该海区输入了大量无机氮, 无机氮过剩而未被完全消耗(图 3)。GO情景对黄海中部的输运高于 AM情景, 导致 GO情景无机氮含量增速较快。GO和AM情景净初级生产增长分别为 6%和 17%, 矿化随之分别增加 87.6×103t/a和238.5×103t/a。前文分析表明黄海中部层化季节AM情景下底层无机氮浓度增加显著, 且与表层无机氮浓度减少区域一致, 这可能与生物量升高碎屑沉降在底层矿化有关。由于底层水团的稳定性, 再生的无机氮未被消耗, 导致底层营养盐浓度升高。在长江口邻近海区, 长江-钱塘江是重要的无机氮源, GO和 AM 情景下河流输入导致的无机氮增加分别为683.5×103t/a和152.2×103t/a。该海区无机氮浓度较高,且向其它海区大量输出, 对外输出分别增加 656.6×103t/a和42.5×103t/a。由于AM情景下无机磷载荷快速增长, 消耗了大量无机氮, 因此净初级生产增加了34%, 高于GO情景(15%)。东海陆架区两个情景下对流输入均有所减少, 这是环流场改变的结果, 应与未来气候情景冬季风减弱、夏季风增强有关。
未来与现代情景下无机磷的收支变化与无机氮并不对应(表6), 各区域光合作用吸收、呼吸释放、水体矿化增加, 与无机氮收支中增加比例一致。而对流输运变化不同, AM情景无机磷输运高于GO情景, 河流输入增幅更大。相对于现代情景, 黄海沿岸GO和AM情景物理过程导致的无机磷增加分别为 13.7× 103t/a和 28.3×103t/a, 其中对流输运对该海域无机磷增加贡献较大; 黄海中部对流输入导致无机磷分别增加了6.8×103t/a和 18.7×103t/a, 使该海区无机磷浓度增加;长江口邻近海区物理过程导致无机磷增量分别为10.4×103t/a和 17.0×103t/a, 支撑了无机磷浓度的增加; 东海陆架区对流输入的无机磷减少, 无机磷浓度变化不大。

表6 各海区物理过程和生物过程无机磷收支的贡献(单位: ×103t/a)Tab.6 Contribution of physical and biological process to the budgets of DIP (×103t/a) in each box
收支分析可以用来了解海区未来营养盐浓度变化的主要来源以及内部循环过程随河流输入增加、对流输运改变的调整。GO和AM两个情景比较, GO情景下无机氮增幅较大, 但大部分海区氮已过剩, 相比之下, AM情景下无机磷的迅速增长更能影响各海区的生物过程, 使其大幅增强。
5 结论
本文基于 FGOALS模式对未来气候情景的预测结果, 结合未来河流营养盐载荷特征, 利用黄、东海水动力模型和生态模型并采用降尺度的方法对未来GO和AM情景下黄、东海营养盐的分布特征进行了情景预测, 并通过敏感性实验并结合收支分析, 对各海区水动力未来变化以及不同情景下河流载荷变化的相对贡献进行了评估, 分析了各海区未来两个情景下营养盐浓度相对于现代情景改变的主要影响因素。主要结论如下:
(1) GO情景下, 无机氮含量增加较快, 河口邻近海区、苏北浅滩及黄海中部无机氮浓度显著增加, 各海区磷限制更加显著; AM 情景下, 无机磷含量增加较快, 夏季黄海中部表层无机氮浓度随生物消耗而显著下降, 海区氮磷比有所下降, 但营养盐浓度增加会使海区富营养化加剧, 生物过程大幅增强。
(2) 水动力环境变化与河流营养盐排放量增加这两个因素, 后者是未来黄海沿岸、黄海中部和长江口邻近海区营养盐浓度增加的主要原因。东海陆架区营养盐浓度变化很小。
(3) 分析各海区营养盐内部循环和外部输运各收支分量, 未来黄海沿岸营养盐浓度增加是对流输入增加和河流输入增加导致的; 黄海中部表层受夏季冲淡水输运影响, 底层营养盐浓度增加则是由于夏季生物量升高造成更多碎屑沉降在底层矿化以及冬季对流、混合过程将沿岸河流输入的营养盐带入底层造成的; 长江口邻近海区受河流输入增加的影响,营养盐浓度增加主要发生在 10m以浅的水层; 东海陆架区生物、物理过程收支平衡, 营养盐浓度不变。
由于未来黑潮等开边界流量、营养盐浓度变化尚不明确, 本研究仅考虑了上边界大气动力-热力强迫和河流营养盐载荷的未来情景, Zhao等(2011)已通过敏感性实验研究了黑潮所携带营养盐输入的变化对黄、东海营养盐和初级生产的影响, 本文不再赘述。目前未来预测的大气强迫分辨率很低, 将来需要在区域大气模式和大小区水动力-生态模式嵌套方面进一步考虑降尺度方法, 完善预测模式。另外, 目前的气候预测有很大的不确定性, 该工作只是一个未来海洋生态环境情景预测的尝试, 未来的研究中将具体分析生态环境如黄、东海初级生产以及浮游植物群落组成的未来状态。
李家星, 魏 皓, 罗晓凡, 2012. 东海陆架表层水温年际变化时空特征分析. 海洋与湖沼, 43(3): 624—634
张维娟, 杨 波, 魏 皓, 2014. 四个耦合模式在模拟和预测东亚季风系统方面的对比分析. 海洋科学, 38(9): 96—108
赵 亮, 魏 皓, 冯士筰, 2002. 渤海氮磷营养盐的循环和收支. 环境科学, 23(1): 78—81
唐启升, 苏纪兰, 2001. 海洋生态系统动力学研究与海洋生物资源可持续利用. 地球科学进展, 16(1): 5—11
海洋图集编委会, 1993. 渤海、黄海、东海海洋图集(水文分册).北京: 海洋出版社, 1—530
袁承仪, 2011. 黄海水温与环流变化特征及机制研究. 青岛:中国海洋大学硕士学位论文, 1—97
Aksnes D L, Ulvestad K B, Balino B Metal, 1995. Ecological modelling in coastal waters: Towards predictive physicalchemical-biological simulation models. Ophelia, 41(1): 5—36
Alcamo J, Van Vuuren D, Cramer W, 2006. Changes in ecosystem services and their drivers across the scenarios.In:The Millennium Ecosystem Assessment ed. Ecosystems and Human Well-Being: Scenarios. Washington D C, USA:Island Press: 279—354
Bouwman A F, Beusen A H W, Billen G, 2010. Human alteration of the global nitrogen and phosphorus soil balances for the period 1970-2050. Global Biogeochem Cycles, 23(4),GB0A04, doi:10.1029/2009GB003576
Fekete B M, Wisser D, Kroeze Cetal, 2010. Millennium Ecosystem Assessment Scenario drivers (1970-2050):Climate and hydrological alterations. Global Biogeochem Cycles, 24(4), GB0A12, doi:10.1029/2009GB003593
Garnier J, Beusen A, Thieu Vetal, 2010. N:P:Si nutrient export ratios and ecological consequences in coastal seas evaluated by the ICEP approach. Global Biogeochem Cycles, 24(4),GB0A05, doi:10.1029/2009GB003583
Guo X Y, Miyazawa Y, Yamagata T, 2006. The Kuroshio Onshore Intrusion along the Shelf Break of the East China Sea: The Origin of the Tsushima Warm Current. J Phys Oceanogr,36(12): 2205—2231
IPCC, 2007. Climate Change 2007: Synthesis Report. Geneva,Switzerland: IPCC, 104
Li M T, Xu K Q, Watanabe Metal, 2007. Long-term variations in dissolved silicate, nitrogen, and phosphorus flux from the Yangtze River into the East China Sea and impacts on estuarine ecosystem. Estuar Coast Shelf Sci, 71(1—2):3—12
Liu S M, Hong G H, Zhang Jetal, 2009. Nutrient budgets for large Chinese estuaries. Biogeosciences, 6(10): 2245—2263
Moll A, 1998. Regional distribution of primary production in the North Sea simulated by a three-dimensional model. J Marine Syst, 16(1—2): 151—170
Qu H J, Kroeze C, 2010. Past and future trends in nutrients export by rivers to the coastal waters of China. Sci Total Environ, 408(9): 2075—2086
Seitzinger S P, Mayorga E, Bouwman A Fetal, 2010. Global river nutrient export: A scenario analysis of past and future trends. Global Biogeochem Cycles, 24(4), GB0A08,doi:10.1029/2009GB003587
Skogen M D, Svendsen E, Berntsen Jetal, 1995. Modeling the primary production in the North Sea using a coupled three-dimensional Physical-Chemical-Biological Ocean Model.Estuar Coast Shelf Sci, 41(5): 545—565
Strokal M, Yang H, Zhang Y Cetal, 2014. Increasing eutrophication in the coastal seas of China from 1970 to 2050. Mar Pollut Bull, 85(1): 123—140
Van Drecht G, Bouwman A F, Harrison Jetal, 2009. Global nitrogen and phosphate in urban wastewater for the period 1970 to 2050. Global Biogeochem Cycles, 23(4), GB0A03,doi:10.1029/2009GB003458
Wang B D, 2006. Cultural eutrophication in the Changjiang(Yangtze River) plume: History and perspective. Estuar Coast Shelf Sci, 69(3—4): 471—477
Yang S L, Gao A, Hotz H Metal, 2005. Trends in annual discharge from the Yangtze River to the sea (1865-2004).Hydrolog Sci J, 50(5): 825—836
Yang D Z, Yin B S, Liu Z Letal, 2011. Numerical study of the ocean circulation on the East China Sea shelf and a Kuroshio bottom branch northeast of Taiwan in summer. J Geophys Res, 116(C5), C05015, doi:10.1029/2010JC006777
Zhao L, Guo X Y, 2011. Influence of cross-shelf water transport on nutrients and phytoplankton in the East China Sea: a model study. Ocean Sci, 7: 27—43
Zhou M J, Shen Z L, Yu R C, 2008. Responses of a coastal phytoplankton community to increased nutrient input from the Changjiang (Yangtze) River. Cont Shelf Res, 28(12):1483—1489
猜你喜欢
草业学报(2022年5期)2022-07-08
海洋信息技术与应用(2021年3期)2022-01-17
草业学报(2018年10期)2018-10-19
草业科学(2018年7期)2018-07-26
建筑热能通风空调(2018年5期)2018-07-09
草业科学(2017年7期)2017-08-11
中国工程咨询(2016年9期)2016-02-13
广东海洋大学学报(2015年3期)2015-12-22
应用海洋学学报(2015年3期)2015-11-22
西安建筑科技大学学报(自然科学版)(2014年2期)2014-11-12
