
云南省绒鼠属(Eothenomys)的形态适应研究
2015-03-21 11:28朱万龙
生物学杂志 2015年1期
沐 远, 朱万龙
(云南师范大学 生命科学学院, 昆明 650500)
云南省绒鼠属(Eothenomys)的形态适应研究
沐 远, 朱万龙
(云南师范大学 生命科学学院, 昆明 650500)
绒鼠属(Eothenomys)是亚洲横断山地区的特有类群。对分布于云南地区的昭通绒鼠(E.oliter)、玉龙绒鼠(E.proditor)、西南绒鼠(E.custos)、滇绒鼠(E.eleusis)、黑腹绒鼠(E.melanogaster)、大绒鼠(E.miletus)、克钦绒鼠(E.cachinus)和中华绒鼠(E.chinensis)形态学数据进行了测量,利用判别分析、形态的系统发生等方法对其进行分析。结果表明云南横断山垂直分布的特点决定了海拔对绒鼠形态的影响比纬度更加明显,绒鼠中出现的不同表现型是其不同生活方式和对不同生境长期适应的结果,但在形态上滇绒鼠和黑腹绒鼠有重叠。
绒鼠属;形态学;扩散
研究表明环境在动物形态的表型可塑性上起到了重要的作用[1-2]。动物的栖息环境一旦发生改变可以引起表型的多样性随之改变[3-4]。动物表型上的改变显示出对其不同环境适应的结果[5],甚至可以通过形态特征的变化来推断动物在不同环境下是怎样进化的[6]。随着地理分布不同,如纬度、海拔的改变,不同地区气候差异,长期适应的结果使得一些小型哺乳动物在种内或种间出现形态学上的差异[7]。这些差异一定程度上是由于环境温度等生态因子的改变进一步影响植被的变化,最终长期积累后使得动物在利用不同的自然资源时出现了不同的适应结构,这些影响一定程度表现在其骨骼和身体大小的变化[8]。
绒鼠属(Eothenomys)是亚洲横断山脉地区的特有类群[9](图1)。一直以来,绒鼠属的分类地位及有多少个有效种一直存在争议[10-15]。绒鼠主要栖息于高原山林区,绒鼠化石最早发现于第三纪,从化石上看,其总体形态与现今种类的差异不大[9]。但随着时间的推移,纬度、海拔的变化会在一定程度上影响绒鼠形态发生变化[16]。关于中国绒鼠属形态学适应研究还未见报道。本文以云南分布的几种绒鼠为研究对象,对其形态学数据进行测量,为探讨横断山地区小型哺乳动物的形态适应提供一些基础数据。
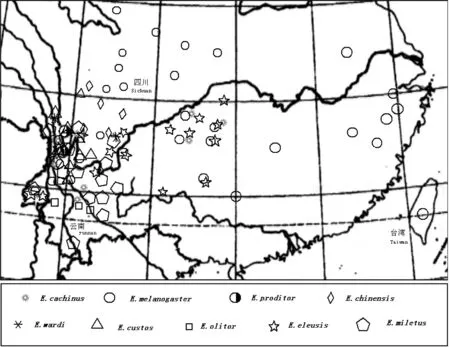
图1 中国绒鼠的分布图(根据《中国动物志》绘制)
1.1 动物来源
实验动物分别捕自剑川(2550~2590 m;N26°15′~26°45′;E99°40′~99°55′)、昭通(1952~1983 m;E103°68′~103°87′;N29°34′~29°60′)、丽江(2416~2483 m;E100°5′~100°25′;N26°84′~26°95′)、德钦(3362~3410 m;N28°29′;E98°54′)和贡山(1755~1789;E98°45′~98°63′;N27°53′~27°89′)的农田和灌木丛中,所有绒鼠均为成年健康个体。
1.2 外型及头骨测量
称重处死后用游标卡尺分别测量了编号动物13项数据,包括:体长(head and body length, HBL);尾长(tail length, TL);耳长(ear length, EL);后足长(hind foot length, HFL);颅全长(greatest length, GTL);基长(basal length);腭长(mandibular length, ML);颧宽(zygomatic breadth, ZB);眶间宽(narrowest width of the skull across the interorbital region, IC);颅高(greatest high, GH);听泡长(length of tympanic bulla, TB);上齿列长(length of the upper cheek teeth row, UM);下齿列长(length of the lower cheek teeth, LM)。精确到0.01 mm。
1.3 数据处理
采用SPSS13.0方差分析、判别分析进行试验数据的统计。文内数值均以平均值±标准误(Mean±SE)表示,P<0.05为差异显著。应用Mesquite中的Brownian model对形态学数据进行分析构建系统树[17]。
2 结果
2.1 物种鉴定
所捕捉的绒鼠经鉴定后为昭通绒鼠(Eothenomysoliter)、玉龙绒鼠(E.proditor)、西南绒鼠(E.custos)、滇绒鼠(E.eleusis)、黑腹绒鼠(E.melanogaster)、大绒鼠(E.miletus)、克钦绒鼠(E.cachinus)和中华绒鼠(E.chinensis)。
2.2 形态学数据
外形数据中,昭通绒鼠的体重与其它物种差异极显著(P<0.01),滇绒鼠和黑腹绒鼠差异不显著,克钦绒鼠、中华绒鼠、西南绒鼠、大绒鼠和玉龙绒鼠之间差异不显著。体长:昭通绒鼠与其它种差异极显著(P<0.01),而滇绒鼠、西南绒鼠和玉龙绒鼠之间差异不显著,玉龙绒鼠、克钦绒鼠和大绒鼠之间差异不显著;大绒鼠和中华绒鼠差异不显著。尾长:昭通绒鼠与玉龙绒鼠差异不显著,黑腹绒鼠和滇绒鼠、西南绒鼠间差异不显著,克钦绒鼠和大绒鼠差异不显著,中华绒鼠与其它物种差异均达到了显著水平(P<0.05)。后足长度:昭通绒鼠、滇绒鼠和黑腹绒鼠差异不显著,西南绒鼠、克钦绒鼠和玉龙绒鼠间差异不显著,克钦绒鼠、玉龙绒鼠和大绒鼠之间差异不显著,中华绒鼠和其它差异均达显著水平(P<0.05)。耳长:昭通绒鼠、黑腹绒鼠和滇绒鼠之间差异不显著,黑腹绒鼠、滇绒鼠和克钦绒鼠间差异也不显著水平,克钦绒鼠和玉龙绒鼠见差异不显著,玉龙绒鼠、西南绒鼠和大绒鼠差异不显著,中华绒鼠与其它物种差异达到了显著水平(P<0.05)。
在头骨数据中,昭通绒鼠的颅全长与其它种类达到了显著水平(P<0.05),滇绒鼠、黑腹绒鼠和西南绒鼠之间差异不显著,西南绒鼠和克钦绒鼠差异不显著,克钦绒鼠、玉龙绒鼠间差异不显著,玉龙绒鼠和中华绒鼠的差异也不显著。基长:昭通绒鼠与其它种类达到了显著水平(P<0.05),黑腹绒鼠、滇绒鼠、西南绒鼠见差异不显著,西南绒鼠和克钦绒鼠差异不显著,克钦绒鼠和玉龙绒鼠间差异不显著,玉龙绒鼠、大绒鼠与中华绒鼠间差异不显著。腭长和颧宽方面:昭通绒鼠与其它种类差异同样达到了显著水平(P<0.05),黑腹绒鼠与西南绒鼠、滇绒鼠之间的差异仍不显著,西南绒鼠和克钦绒鼠间差异不显著,克钦绒鼠,玉龙绒鼠间差异不显著,玉龙绒鼠、大绒鼠和中华绒鼠间差异不显著;但在上下齿列长方面昭通绒鼠仍然和其它种类有着显著的差异,但在玉龙绒鼠出现了差异,黑腹绒鼠、西南绒鼠、滇绒鼠和玉龙绒鼠上齿列长没有显著差异,而玉龙绒鼠下齿列长却与之有显著差异,玉龙绒鼠和克钦绒鼠上齿列长差异不显著,中华绒鼠和大绒鼠之间差异也不显著,下齿列长玉龙绒鼠和克钦绒鼠、中华绒鼠差异不显著,中华绒鼠和大绒鼠间差异不显著。

表1 8种绒鼠的测量数据
2.2 形态学数据的系统分析
经判别分析,中华绒鼠和昭通绒鼠不论是在外型上还是在头骨方面都能和其他物种有很大的区别,能比较容易地将其区分出来。而玉龙绒鼠、西南绒鼠、滇绒鼠、黑腹绒鼠、大绒鼠和克钦绒鼠较集中,而且有些个体之间会出现重叠,如黑腹绒鼠和滇绒鼠基于头骨的判别分析中就基本重叠在一起。
基于形态学数据构建的系统树发现,系统树分为两大支,其中中华绒鼠和西南绒鼠聚为一大支,而其他6种绒鼠聚为另一大支。6种绒鼠中滇绒鼠和大绒鼠又和黑腹绒鼠等、昭通绒鼠以及克钦绒鼠、玉龙绒鼠分为两支(图2)。

图2 基于形态学数据构建的系统树
3 讨论
自1896年Miller首次建立了Eothenomys亚属,绒鼠属分类的讨论一直没有停止[12- 13]。其中,在绒鼠属与其近似类群在属级和亚属级分类上相当的混乱,各学者的观点不一。在属下分类上,大部分学者认为应分为3个亚属[11];而Ellerman[15]、王应祥[18]、马勇[19]和罗泽珣[20]认为绒鼠属只分为2个亚属。而且在种级分类上主要的争议在9个种[21]。通过形态学数据的判别分析后,发现绒鼠在外形和头骨基本形态学数据上明显区分昭通绒鼠和中华绒鼠,玉龙绒鼠、西南绒鼠、大绒鼠和克钦绒鼠均能从基本形态上加以区别。但其中黑腹绒鼠和滇绒鼠在形态上发生了重叠,这种重叠可能与其栖息环境有关。通过系统发生分析结果表明:它们两者之间形态系统发生中存在着差异。研究发现大绒鼠和滇绒鼠虽然在分子系统演化上不是独立的分类单位[22],但从形态上来看,两者之间的差异是明显的,但通过形态系统分析后发现两者在进化上的地位是比较接近的。
“是否形态结构能指示出其生态情况”的问题在很早就提出了[23],个体的不同形态和其生态环境之间的关系是关联的[24]。绒鼠属由于其外形差异不大,部分种类牙齿形态不稳定,分类上意见分歧较大。根据化石记载,其分化可能在上新世[9]。随着时间推移,喜马拉雅造山运动使横断山区具有多种垂直分布带,植被的地域分异明显,且物种丰富[25]。地理变化成为许多地方脊椎动物形成的重要影响因素[26-28]。研究结果表明:绒鼠仅仅只用体长和尾长之间的关系是很难区分中华绒鼠和西南绒鼠、西南绒鼠和玉龙绒鼠,但通过运用体长、尾长、耳长、后足长、颅全长、基长、腭长、颧宽、眶间宽、颅高、听泡长、上齿列长和下齿列长等参数的判别函数分析能够将中华绒鼠和西南绒鼠、西南绒鼠和玉龙绒鼠分开,而且易于区分昭通绒鼠和中华绒鼠。但黑腹绒鼠和滇绒鼠不仅在体形外部出现重叠,而且在头骨形态上也出现重叠,此时头骨形态上部分也与西南绒鼠重叠,这种重叠可能是由于其栖息环境和行为特征有关。运用颅全长和颅高来分析绒鼠体形大小及变化,中华绒鼠都大于其余种类,可能由于云南平均气温,除干热河谷外,基本是从南往北渐减[28]。中华绒鼠主要分布在四川西部的川西山地,比较其他种类靠云南北部,生活于温度相对低的地区,其体形有变大的趋势。特别是在冬季和春季,保证其有充足的食物,就会影响动物生活史,增加动物体大小[7]。中华绒鼠分布的海拔从300~3100 m的阴湿阔叶林,稀树灌丛及农耕地附近[24],云南的四季不分明的气候特征保证其有充足的食物,这也对其有较大的体形有一定的影响,单位体长的体重上明显低于玉龙绒鼠。由此推测,其虽然栖息于云南偏北地区,但由于生活纬度比黑腹绒鼠等较低,横断山地区的垂直分布对绒鼠的形态影响更大。玉龙绒鼠和大绒鼠在体形大小仅次于中华绒鼠,但在单位体长上的重量却又都高于中华绒鼠。在单位体长上的重量,玉龙绒鼠明显高于其他种类。玉龙绒鼠主要分布于中国云南丽江横断山北段高山峡谷地区,海拔为2800~4200 m的林地或草甸[24],是几种绒鼠中分布海拔最高。这些地方年均温在10℃左右,处于寒温带和暖温带地区,所以需要增加隔热性来抵抗低温。低温是影响动物大小的一个主要因素[29]。面对温度的压力,玉龙绒鼠不仅体形较大,而且增加隔热性来抵抗低温,使得其在单位体长上的重量处于最高的水平。而对于大绒鼠来说,其它绒鼠主要是在晨昏外出活动、觅食,但是大绒鼠多在夜间、晨昏外出活动、觅食[27]。并且自第四纪以来,由于冰川的活动和云贵高原的抬升,导致气候特征从湿润、温暖向干燥、寒冷的方向演化,并使气温年温差小,日温差大。这样,夜间活动的绒鼠就需要有一定的生理生态适应方式,使得其体形和单位体长的重量都比较高。西南绒鼠、滇绒鼠、黑腹绒鼠和克钦绒鼠在体形大小上的变化都不是很明显。中国动物志记载,这4种绒鼠主要栖息地海拔分别是1100~3800 m、1000~3000 m、1000~3000 m和2000~3000 m的阴湿林地[21],且几乎都是黄昏后觅食,所以在形态上的差别不是很明显,但在单位体长体重来看,滇绒鼠和黑腹绒鼠要偏低。本研究中,滇绒鼠和黑腹绒鼠是分布比较靠东,在中国大部分南部都分布有黑腹绒鼠,滇绒鼠也广泛地分布到了华中地区,贵州、四川、湖北及湖南等。在这些地方,气候条件明显与云南有差别,所以在冬季和春季又面临着食物的压力,其在单位体长的体重上与西南绒鼠和克钦绒鼠有着明显的差异。昭通绒鼠在这几种绒鼠中,无论在体形大小还是在单位体长的体重上都是处于最低水平。通过对玉龙绒鼠和中华绒鼠的比较后发现,在云南横断山垂直分布特征明显的特点可能海拔高度对绒鼠形态的影响比纬度更加明显。
形态构建的系统树表明大绒鼠和滇绒鼠在形态上比较靠近。玉龙绒鼠和克钦绒鼠聚在一起,中华绒鼠和西南绒鼠在一起。黑腹绒鼠和昭通绒鼠又较为靠近。可以看出中华绒鼠栖息的海拔300~3100 m,大绒鼠和滇绒鼠都是生活于海拔为1000~3000 m的地区,而玉龙绒鼠和德钦绒鼠分布的海拔分别为2800~4200 m和2400~4300 m等海拔较高的地区[21],推测绒鼠在分化过程中,可能一部分往高海拔区域扩散,而另一部分则向低处延伸逐步适应其生活环境。黑腹绒鼠分布最广,可能由于其在向平原地区扩散的过程比较晚。由此推断可能绒鼠在整个扩散过程起始比较晚,所以大部分还集中分布于中国西部的横断山,但横断山具有多种垂直分布带和独特的地形地貌最终导致物种对环境的适应,形成现在的分布格局和形态特征。
综上,除了滇绒鼠和黑腹绒鼠在形态上有重叠外,其余物种在一定程度上都能通过判别函数分析区分出来,且这些差异和其所栖息的环境之间有着重要的联系。
[1]Ane T L, Anssi L K, Ingemar J, et al. Do common frogs (Ranatemporaria) follow Bergmann′s rule?[J] . Evolutionary Ecology Research, 2005, 7:717-731.
[2]Miguel A R, Irene L L, Bradford A H. The geographic distribution of mammal body size in Europe[J]. Global Ecology and Biogeography, 2006, 15 (2):173-181.
[3]Hadany L, Beker T. On the evolutionary advantage of fitness-associated recombination[J]. Genetics, 2003, 165:2167-2179.
[4]Price T D, Qvarnstrom A, Irwin D E. The role of phenotypic plasticity in driving genetic evolution[J]. Proceedings of the Royal Society of London, 2006, 270B:1433-1440.
[5]Rutherford S L. Between genotype and phenotype: Protein chaperones and evolve-ability[J]. Nature Reviews, 2003, 4:263-274.
[6]Sabrina R, Jacques M, Daniela N, et al. Morphological evolution, ecological diversification and climate change in rodents[J]. Pro R Soc B, 2005, 272: 609-617.
[7]Yoram Y T, Shlomith Y T. Climatic change and body size in two species of Japanese rodents[J]. Journal of the Linnean Society, 2004, 82: 263-267.
[8]Smith F A, Browning H, Shepherd U L. The influence of climate change on the body mass of wood rats Neotoma in arid region of New Mexico, USA[J]. Cerography, 1998, 21: 140-148.
[9]郑少华.川黔地区第四纪啮齿动物化石[M].北京:科学出版社, 1993.
[10]Allen G M. Some Chinese vertebrates, mammalian[J]. Memories of the Museum of Comparative Zoology of Harvard College, 1912, 40: 201-247.
[11]Allen G M. Mammals of China and Mongolia[M]. New York: American Museum of Natural History, 1940.
[12]Hinton M A C. On the voles collected by Mr. G. Forrest in Yunnan; with remarks upon the generaEothenomysandNeodonand upon their allies[J]. Annals and Magazine of Natural History, 1923, 9(11): 145-162.
[13]Hinton M A C. Monograph of the voles and lemmings (Microtinae), living and extinct[M]. London: British Museum (Natural History), 1926.
[14]Ellerman J R. The families and genera of living rodents[M]. London: British museum (Natural History), 1941.
[15]Ellerman J R, Morrison S. Checklist of Palaearctic and Indian mammals 1758 to 1946[M]. London: British Museum (Natural History), 1951.
[16]Yukibumi K. Morphological variation and geographical and altitudinal distribution inEothenomysmelanogasterandE.ucronatus(Rodentia, Arvicolinae) in China, Taiwan, Bruma, India, Thailang, and Vietnam[J]. Mammal Study, 2002, 27:31-63.
[17]Maddison W P, Maddison D R. Mesquite: a modular system for evolutionary analysis[M]. Version 1.1. http://mesquiteproject.org, 2006.
[18]王应祥.中国哺乳动物种和亚种分类名录与分布大全[M]. 北京: 中国林业出版社, 2003.
[19]马 勇, 姜建青. 绒鼠属Caryomys地位的恢复(啮齿目:仓鼠科:田鼠亚科)[J].动物分类学报, 1996, 21(4):493-497.
[20]罗泽珣, 陈卫等. 中国动物志:兽纲第六卷啮齿目下册仓鼠科[M].北京:科学出版社, 2000.
[21]刘少英, 刘 洋. 绒鼠系统学研究概况[J]. 四川动物, 2005, 24(1): 98-103.
[22]Luo J, Y D, Suziki H, et al. Molecular phylogeny and biogeography of oriental voles: genus Eothernomys (Muridae, Mammalia)[J]. Molecular Phylogenetics and Evolution, 2004, 33:349-362.
[23]Losos J B. Geomorphology, performance capability, and scaling of West IndianAnolislizarda: an evolutionary analysis[J]. Ecol Monogr, 1990, 60:369-388.
[24]Wainwright P C. Functional morphology as a tool in ecological research. In: ecological morphology: integrative organismal biology[M]. Chicago: University of Chicago Press, 1994.
[25]Pang J F, Wang Y Z, Zhong Y, et al. A phylogeny of Chinese species in the genusPhrynocephalus(Agamidae) inferred from mitochondrial DNA sequences[J]. Mol Phylogenet Evol, 2003, 27:398-409.
[26]Bergmann C. Systematic and the origin of species[M]. New York: Columbia University Press, 1847.
[27]McNab B K. On the ecological significance of Bergman′s rule[J]. Ecology, 1971, 52: 845-854.
[28]许祥誉.云南主要生态类型及开发[M]. 昆明: 云南大学出版社. 1993.
[29]Muller E F. Basal metabolic rate in primate′s the possible role of phylogenetic and ecological factor[J]. Comp Biochem Physiol, 1985, 81(4): 707-711.
Study of morphological adaptation of genusEothenomys(Muridae, Mannalia) in Yunnan province
MU Yuan, ZHU Wan-long
(School of Life Science, Yunnan Normal University, Kunming 650500, China)
Eothenomysare small mammals inhabiting in Hengduan mountain regions. In the present study, morphological examinations ofE.oliter,E.proditor,E.custos,E.eleusis,E.melanogaster,E.miletusandE.cachinusfrom Yunnan were measured. Discriminate analysis and phylogeny of morphology were used to analysis the ecological adaptations and group spread. Results showed thatEothenomysmorphological divergence was prime importance in the interaction of the organism with its environment and life history, and the altitude was more important than latitude. Taxonomic differentiation was commonly associated with morphological divergence, exceptE.eleusisandE.melanogaster.
Eothenomys; morphology; spread
2014-06-17;
2014-08-08
国家国际科技合作项目(No. 2014DFR31040);十二五国家支撑计划(No. 2014BAZ0481500);国家自然科学基金资助项目(No.31360096; 31260097);云南省应用基础研究项目(No.2013FA014)
沐 远,硕士研究生,研究方向为动物生理生态,E-mail: islam19891001@163.com;
朱万龙,副教授,研究方向为动物生理生态,E-mail: zwl_8307@163.com。
Q951+.4
A
2095-1736(2015)01-0014-04
doi∶10.3969/j.issn.2095-1736.2015.01.014
猜你喜欢
昭通学院学报(2022年1期)2022-05-28
考试与评价·高二版(2021年2期)2021-09-10
汉语世界(The World of Chinese)(2019年2期)2019-04-19
启蒙(3-7岁)(2017年4期)2017-06-15
水科学与工程技术(2016年3期)2016-07-10
新时代职业教育(2016年4期)2016-02-06
大众考古(2015年6期)2015-06-26
西南学林(2014年0期)2014-11-12
西南学林(2014年0期)2014-11-12
海外英语(2013年7期)2013-11-22
