
草地生态系统碳通量特征及其影响因子研究
2015-03-20 00:55:24李晓琳
中低纬山地气象 2015年6期
李晓琳
(河南省三门峡市气象局,河南 三门峡 472000)
草地生态系统碳通量特征及其影响因子研究
李晓琳
(河南省三门峡市气象局,河南 三门峡 472000)
该文以内蒙古科尔沁草原为研究对象,利用涡动相关系统观测的2011年生长季(5—9月)的碳通量以及同期常规气象观测数据,分析该生态系统的碳通量的日变化、季节变化及造成碳通量变化的原因,结果表明草地生态系统碳通量日变化呈单峰型;生长季碳通量具有明显的季节变化规律,基本呈“U”型;净辐射和气温是影响草地生态系统碳通量的主要气象因子。
碳通量;草地;涡动相关;影响因子
1 引言
自20世纪70年代以来,亚洲工业排放的 CO2增长了 60% ,增长率高于其他所有地区,而我国则成为亚洲地区第一CO2排放大国,高于工业发达国家日本[1]。草地生态系统作为地球上最重要的陆地生态系统之一,世界草原总面积约占地球陆地总面积的六分之一,中国是世界上草原资源最丰富的国家之一,草原总面积将近4亿hm2,占全国土地总面积的40%[2],为现有耕地面积的3倍。草地生态系统不仅对于发展畜牧业、保护生物多样性以及保持水土等有重要意义,其对地球碳循环的重要意义也十分突出[3]。本研究选取我国四大草原之一的科尔沁草原,通过对其碳通量变化情况以及影响因子进行研究和分析,以更好的揭示科尔沁草原的碳循环规律,为更好的发挥草地生态系统的生态价值提供一定的依据[4]。
2 研究地点和方法
2.1 研究地点
研究地点选取是内蒙古科尔沁草原。位于42°5′~43°5′N,117°30′~123°30′E,海拔250~650 m。气候冬寒冷、夏炎热,春风大。年均降水量360 mm,年际变化较大,年内分配不均,多集中6—8月份。冬季以西北风为主,春秋则对西南风为主,年平均风速3.5 m/s,大风日数常达30 d左右。全区年日照时数2 900 h,日照率达67% ,无霜期140~150 d。植被主要有白草、沙蓬、小黄柳、差巴嘎蒿等[5]。
2.2 观测仪器
研究地建有3 m微气象观测塔,装有一套开路涡动相关系统(Open Path Eddy Covariance,简写为OPEC)和常规气象观测系统,对草地生态系统CO2通量和相关气象要素进行同步测量(表1)。

表1 涡度相关、常规气象和太阳辐射仪器介绍
2.3 研究方法
涡动相关技术是以湍流传输理论和雷诺平均法则为理论基础,某物理量的垂直湍流输送通量可表示为:
(1)
式中,Fc为 CO2的垂直湍流输送通量,w′为垂直风速脉动量,c′为CO2浓度的脉动量。上横线表示在一定时间间隔上的平均。
对于叶面积指数测定,选取试验地每个物种具有代表性植株各30株,然后摘下所选植株叶片,并记录植株叶片数及其长宽等数据,然后称重,最后将其放于印有0.5 cm×0.5 cm方格的A4纸上,通过数字图像处理法测定各植株叶片面积[6]。5—9月的叶面积指数变化范围为0.32~5.93。
空气的饱和水汽压差(VPD),通过空气的水汽压(ea)和相对湿度(Rh)来计算。根据关系式
VPD=es-ea
(2)
(3)
式中,es为饱和水汽压(kPa)
由上面两公式可得:
(4)
由此公式求得空气的饱和水汽压差的数据用于本文的分析。
3 结果分析
3.1 碳通量的日变化和季节变化
通过应用涡度相关系统的测量可以得到所研究草地生态系统中CO2通量在草地生态系统与大气系统之间的交换状况。分析观测所得数据可以得出草地生态系统的光合作用强度与其呼吸作用的强弱关系,当草地生态系统的光合作用大于其呼吸作用时,此时CO2通量视为负值;反之当生态系统的光合作用强度小于其呼吸作用强度时,此时的CO2通量表现为正,如果光合强度与呼吸强度基本在同一水平线上时,则整个草地生态系统与大气生态系统之间的净CO2交换为零。
3.1.1 碳通量的日变化 由于晴天和多云天碳通量日变化的不同,在该草地生态系统3个不同生长时期分别选择1个典型的晴天和多云天进行碳通量日变化的分析。
科尔沁草地7月份晴天碳通量具有明显的日变化规律(图1a),日振幅较大,从07时开始随着太阳辐射的增强和气温的升高(图2a、3a),净光合速率逐渐增大。其中CO2净吸收峰值出现在当日的14时左右,峰值约为1.32 mg·m-2·s-1。下午随着太阳辐射和气温的降低(图2a、3a),净光合速率逐渐降低,至18时左右变为正值。而整个夜间碳通量都为正值,生态系统表现为CO2净排放,在22时30分左右达到排放最大值,为0.61 mg·m-2·s-1。生长初期的5月份和生长末期的9月份,碳通量日变化幅度较小,但有较为明显的日变化规律,碳通量在白天随着太阳辐射和气温的升高(图2a、3a),光合速率不断增强,在正午时分达到峰值,而后又随太阳辐射和气温的降低(图2a、3a),碳通量又开始下降。5月份和9月叶面积指数较低(5月4日为0.39,9月1日为1.87),气温较低(图2a),太阳辐射稍低(图1a), 因而生态系统的光合速率较低。
7月份叶面积指数达到最大,气温也最高,各项环境因素都有利于植被的光合作用,因而净固碳能力最强[7]。图1b为多云天,碳通量在3个不同生长时期的日变化图。由图可明显发现多云天碳通量变化与晴天的差异。相对于晴天,3个时期多云天的碳通量日变化幅度小于同时期晴天的变化幅度,并且阴天的碳通量日变化波动较大,变化曲线不平滑。这主要是由于阴天的净辐射和气温值均小于同期晴天的值(图2b、3b),并且阴天净辐射和气温的波动相较于晴天明显较大(图2b、3b)。
3.1.2 碳通量的季节变化 计算得整个生长季碳通量的日总量,作图分析其季节变化,如图4a所示,并添加趋势线。正值表示CO2的净排放,负值表示CO2的净吸收。在生长季,科尔沁草地生态系统碳通量具有明显的季节变化规律,基本呈“U”型。碳通量随时间的推移而逐渐增大,在6—7月份达到最大,随后又逐渐减小,在9月份以后碳通量变为正值,释放CO2。通过计算得出碳通量月值,用月值作图4b,由图可更直观看出碳通量随季节的变化。在5月份,碳通量月总量绝对值为160.29 g·m-2,而后碳通量月总量绝对值变大,6—7月份相差不大,7月份达到最大值517.41 g·m-2,较5月份值翻了3倍还要多,随后开始下降,到8月份碳通量月总量绝对值为226.70 g·m-2,9月份碳通量月总量已变为正值,变为CO2净排放,值为83.08 g·m-2。
出现上述变化特征,主要受叶面积指数、气温和净辐射的影响。在生长季的初期,主要是5月,叶面积指数为0.33~2.78,气温较低,月平均值为15.61℃,因而植被的光合速率较低,5月碳通量的月总量较小。随后气温不断上升(图5)和叶面积指数也不断增大,在6—7月,叶面积指数大约为5~6,气温在20~25℃之间,净辐射充足,这段时间的植被生长情况和各项环境因子都达到了植被光合作用所需的最佳要求,因此这段时间CO2的净吸收达到整个生长季的最高峰。旺盛期过后,8月叶面积指数开始降低,气温和净辐射也出现下降趋势,导致植被生态系统呈现不断衰退状态,CO2净吸收量不断变小,8月的碳通量已不及7月的一半。而在9月,气温明显降低,净辐射明显减弱(图5),叶面积指数也迅速减少,多种原因导致生态系统9月变为碳源,净排放CO2。
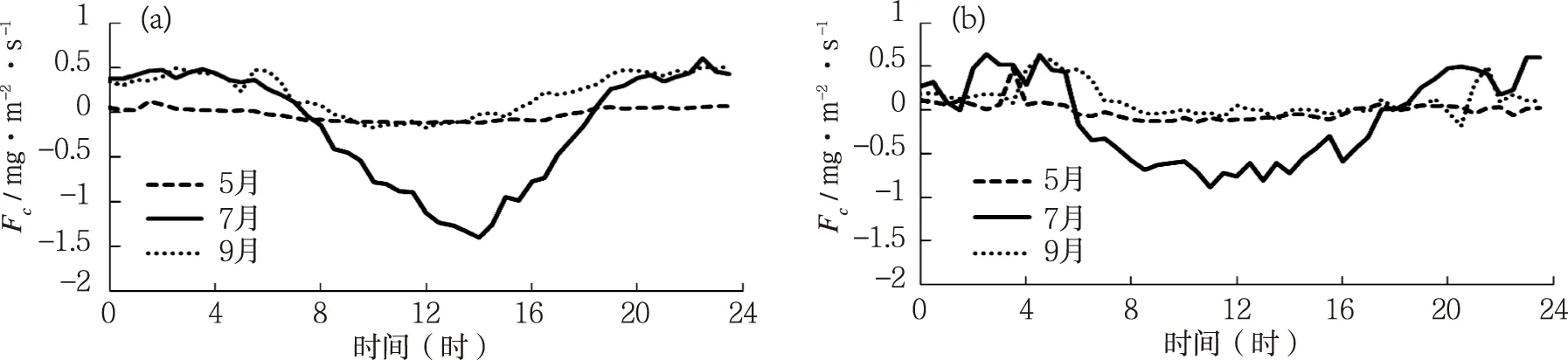
图1 不同生长时期晴天和多云天碳通量的日变化特征
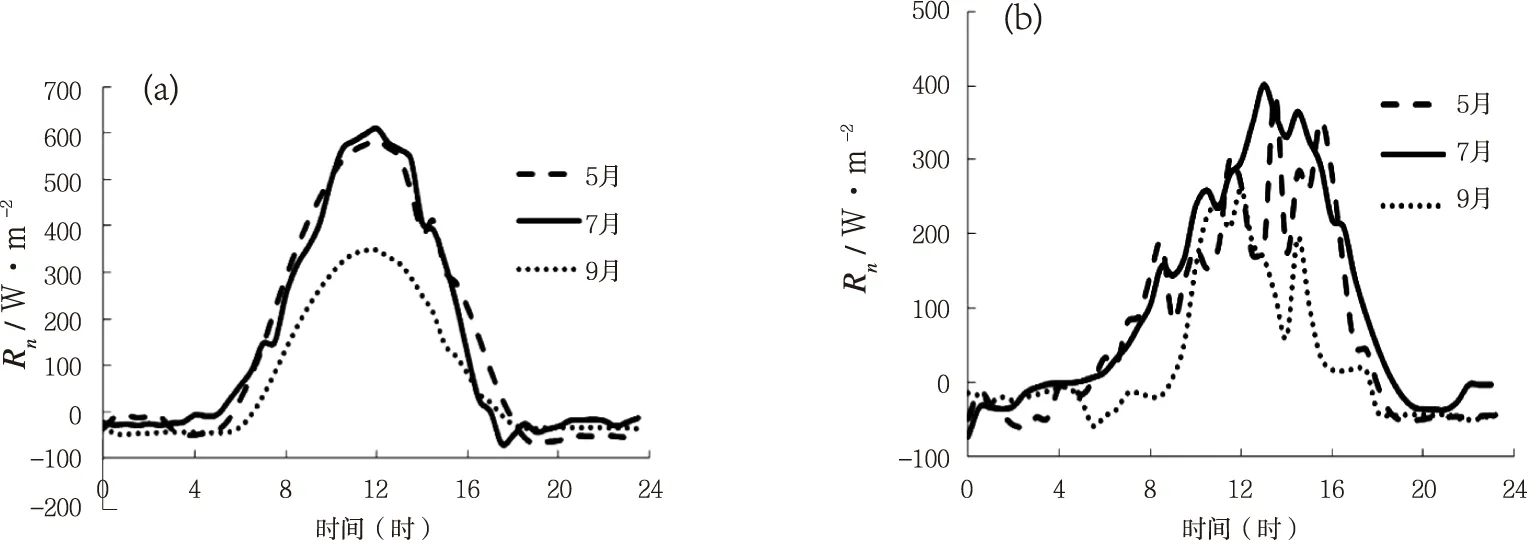
图2 净辐射日变化

图3 气温日变化

图4 草地碳通量季节变化特征

图5 净辐射的季节变化特征
3.2 CO2通量与环境因子的关系
3.2.1 碳通量与气象因子的关系 从以上所分析的结果中我们初步得知,影响草地生态系统碳通量的气象因子主要是净辐射和气温。2011年科尔沁草地生态系统生长季(5—9月)碳通量与净辐射和气温的关系,分别如图6、图7所示。

图6 净辐射与碳通量的关系

图7 气温与碳通量的关系
从图6我们可以看出,图中的点较为密集的集中在趋势线上下,由此可见碳通量受净辐射影响很大。净辐射处于-100~100 W·m-2附近时,图中的点分布十分密集,并且上下波动较大,这片区域,呼吸作用起主导作用,而后随着太阳辐射增强,植被光合作用增强,在净辐射值约为500 W·m-2时,碳通量绝对值达到峰值,约为1.8 mg·m-2·s-1,此时为光合作用最强点,随后随着净辐射增强,碳通量绝对值略微变小。净辐射对碳通量的影响,主要通过影响植被的光合作用,充足的净辐射可以提供植被光合作用所需的光能,从而促进植被的光合作用强度,增强其固碳能力[8]。此外,在净辐射达到500 W·m-2以后,碳通量随净辐射的增强而变化的趋势略有放缓,这主要是由于植被光合作用出现了光饱和现象。
从图7可以看出,碳通量与气温也存在其与净辐射类似的关系,随着气温的升高,碳通量绝对值不断变大,在气温低于10℃时,碳通量与温度的变化规律不明显,主要是由于气温较低时,植被的光合作用受到抑制,难以体现出明显的变化规律[9]。而后随着气温的升高,碳通量的变化变大,在气温约为20℃时,碳通量绝对值迅速增大到1.5 mg·m-2·s-1。气温从20~30℃,碳通量均维持在较高水平,说明这个温度段适宜植被的光合作用的进行,在27℃左右,碳通量绝对值达到最大值,约为1.8 mg·m-2·s-1。在气温>30℃以后,碳通量绝对值又逐渐减小,此时温度过高,植被为避免蒸腾作用过强而关闭部分气孔,导致光合作用强度减弱,从而出现此现象。
分别将2011年生长季每半小时的净辐射、气温、饱和水汽压差、土壤温度(5 cm)和风速与碳通量做线性回归分析,各气象因子与碳通量的方程式和R2如表2所示。根据表中的R2值我们可以看得出,对草地生态系统碳通量影响最大的气象因子是净辐射,其次分别是气温、饱和水汽压差和土壤温度,风速对碳通量影响较小。而且对所有气象因子与碳通量的显著性检验结果显示,所有气象因子都达到了极显著水平。

表2 碳通量与气象因子的相关性
**表示p<0.01,*表示p<0.05(Rn为太阳净辐射,Ta为气温,VPD为饱和水汽压差,Ts为土壤温度,Wnd为风速)
3.2.2 碳通量与环境因子的多元回归分析 将碳通量与净辐射、气温、叶面积指数、风速、地温和饱和水汽压差利用SPSS软件进行多元回归分析,如表3所示。由表中内容可知,输入的6个可能影响碳通量的因子共建立了3个模型,其中,以第3个模型(LAI,Ts和Rn)的R2值最大,为0.652,即此模型的拟合度最好;其次是第2个模型(LAI和Ts),R2值为0.606;最后是第1个模型,R2值为0.522。同时由分析结果得知3个模型的回归系数均达到极显著水平。由此可得出结论:碳通量主要受叶面积指数、土壤温度和净辐射影响,而气温、风速和饱和水汽压差对碳通量影响不明显。

表3 碳通量与环境因子的多元回归分析
**表示p<0.01,*表示p<0.05(LAI为叶面积指数,Ts为土壤温度,Rn为太阳净辐射)
由表3可以看出,模型3的拟合度最好,对模型3中3个自变量的数据进行计算得出预测值(Fcp),并作图分析预测值与实际值的关系,如图8,并添加趋势线,并显示公式和R2,预测值与实测值拟合效果较好。
Fcp= 0.657 7Fc- 2.941 6R2= 0.657 7
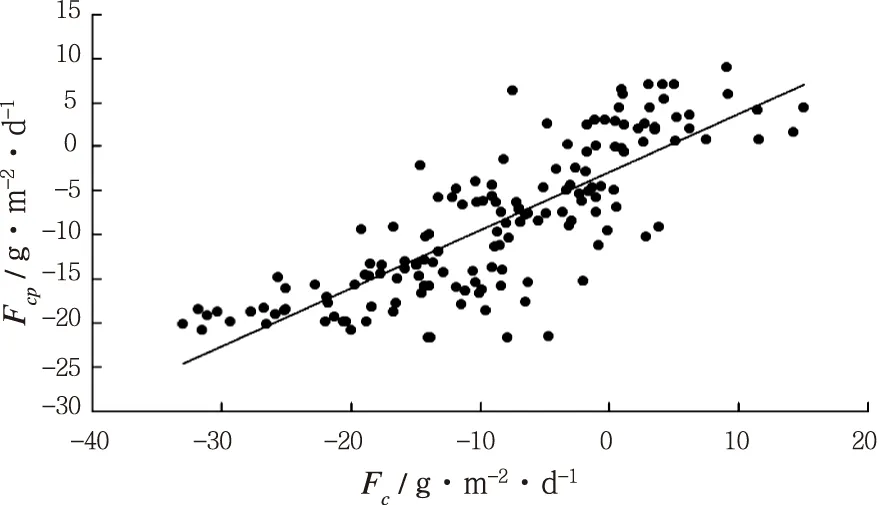
图8 模型预测值Fcp与实测值Fc的关系
4 结论
本研究主要利用涡动相关技术对科尔沁草甸草地生态系统碳通量进行观测,通过观测数据分析碳通量的日变化和季节变化特征,并讨论分析了碳通量与环境因子之间的关系,为科尔沁草地生态系统的碳通量变化特征及其影响因子进行了一定的研究分析。主要结论如下:
① 草地生态系统各生长时期碳通量日变化特征相似,呈单峰型日变化。自日出后,碳通量绝对值逐渐增大,在14时前后达到最大值,此时光合作用最强,然后随着气温和净辐射的不断减小,碳通量绝对值也不断减小。此外,不同的天气状况和不同的生长时期也都会影响碳通量的日变化情况:多云天碳通量变化幅度小于晴天,且波动较大;生长旺盛期碳通量日变化幅度大于生长季初期和末期。
② 草地生态系统碳通量呈明显季节变化,其日值变化曲线接近于“U”型。5月份植被叶面积指数较小、气温较低,碳通量值较小;随着植被叶面积指数的增大和气温的升高,草地生态系统在6—7月份碳通量达到最大值;随后,植被叶面积指数、气温开始降低,8月份碳通量值明显减少;最后的9月份,由于自然和人为(牧民收草)原因导致叶面积指数迅速降低且净辐射和气温的降低,9月份碳通量值为正,排放CO2。整个2011年生长季(5—9月),草地生态系统固定CO2值为1 314.7 g·m-2。
③ 多种气象因子会影响草地生态系统的碳通量,其中净辐射和气温对其影响最大,其次按顺序分别为饱和水汽压差、土壤温度和风速。除气象因子外,植被的叶面积指数对碳通量的影响很大。
[1] UN EP.1999.Global Environment Outlook 2000[R].United Nations Environment Programme,7-8.
[2] 杜容,吕达仁,王庚辰.天然温带典型草原N2,O2和CH4通量的时间变化特征[J].自然科学进展,2015,15(3):313-320.
[3] 谢高地,张钇锂,鲁春霞,等.中国自然草地生态系统服务价值[J].自然资源学报,2001,16(1):47-53.
[4] 苏大学.中国草地资源的区域分布与生产力结构[J].草地学报,1994,2(1):71-77.
[5] 李胜功,赵爱芬,常学礼.科尔沁沙地植被演替的几个问题[J].中国沙漠,1997,17(增刊):25-32.
[6] 李亚军,纪澍琴,张守伟,等.羊草叶面积的测定方法[J].安徽农业科学,2009,37(15):6819-6819.
[7] 李琪,王云龙,胡正华,等.基于涡度相关法的中国草地生态系统碳通量研究进展[J].草业科学,2010,27(12):38-44.
[8] 关德新,吴家兵,于贵瑞,等.气象条件对长白山阔叶红松林 CO2通量的影响[J].中国科学:D辑,2004,34(Ⅱ):103-108.
[9] 项虹艳,朱波,王小国,等.蔬菜地CO2排放特征及其影响因素[J].生态环境,2006,15(1):73-74.
[10] 姚正兰,赵大清,姚章福.遵义市近63 a气象干旱变化趋势及成因分析[J].贵州气象,2015,39(5):08-13.
[11] 陈林琴,刘佳,成春丽等.浅析金沙县气候条件与大棚蔬菜种植的关系[J].贵州气象,2014,38(51):27-28.
2015-07-01
李晓琳(1983—),女,助工,主要从事应用气象研究工作。
1003-6598(2015)06-00019-05
S162
A
猜你喜欢
科学(2022年4期)2022-10-25 02:43:00
农业灾害研究(2022年1期)2022-05-07 01:31:04
湖南农业大学学报(自然科学版)(2021年3期)2021-07-02 01:48:06
新疆农业科学(2020年1期)2020-02-14 03:22:46
青苹果·教育研究版(2016年7期)2016-12-12 05:19:52
Coco薇(2016年5期)2016-06-03 09:17:41
考试周刊(2016年6期)2016-03-11 08:14:32
中国麻业科学(2015年5期)2015-12-28 06:22:11
化工进展(2015年6期)2015-11-13 00:26:29
少儿科学周刊·儿童版(2015年10期)2015-11-07 03:45:31
