
Effects of Trawl Selectivity and Genetic Parameters on Fish Body Length Under Long-Term Trawling
2015-03-15 01:43YUYangSUNPengCUIHeSHENGHuaxiangZHAOFenfangTANGYanliandCHENZelin
YU Yang, SUN Peng, CUI He, SHENG Huaxiang ZHAO Fenfang TANG Yanli and CHEN Zelin
1) College of Environmental Science and Engineering, Ocean University of China, Qingdao 266100, P. R. China
2) Fisheries College, Ocean University of China, Qingdao 266003, P. R. China
3) Chinese Academy of fishery sciences, Beijing 100141, P. R. China
Effects of Trawl Selectivity and Genetic Parameters on Fish Body Length Under Long-Term Trawling
YU Yang1), SUN Peng2),*, CUI He3), SHENG Huaxiang2), ZHAO Fenfang2), TANG Yanli2), and CHEN Zelin2)
1) College of Environmental Science and Engineering, Ocean University of China, Qingdao 266100, P. R. China
2) Fisheries College, Ocean University of China, Qingdao 266003, P. R. China
3) Chinese Academy of fishery sciences, Beijing 100141, P. R. China
© Ocean University of China, Science Press and Springer-Verlag Berlin Heidelberg 2015
Long-term fishing pressure affects the biological characteristics of exploited fish stocks. The biological characteristics of hairtail (Trichiurus lepturus) in the East China Sea are unable to recover because of long-term trawling. Fishing induces evolutionary effects on the fish’s biological characteristics. Evidence of these changes includes small size at age, a shift to earlier age structure, and early maturation. Natural and artificial selection usually affect the fish’s life history. Selection can induce different chances of reproduction, and individual fish can give a different genetic contribution to the next generation. In this study, analysis of time-dependent probability of significance and test of sensitivity were used to explore the effects of fish exploitation rate, mesh size, and heritability with long-term trawling. Results showed that fishing parameters were important drivers to exploited fish population. However, genetic traits altered by fishing were slow, and the changes in biological characteristics were weaker than those caused by fishing selection. Exploitation rate and mesh size exhibited similar evolutionary trend tendency under long-term fishing. The time-dependent probability of significance trend showed a gradual growth and tended to be stable. Therefore, the direction of fishing-induced evolution and successful management of fish species require considerable attention to contribute to sustainable fisheries in China.
trawling pressure; fish biological characteristics; fishery management
1 Introduction
In recent years, fishing-induced evolution of fish phenotypic traits has received considerable attention (Dieckmann and Heino, 2007; Fenberg and Roy, 2007; Kuparinen and Merila, 2007; Hutchings and Fraser, 2008; Dunlop et al., 2009; Enberg et al., 2009, 2012; Jorgensen et al., 2009; Laugen et al., 2014; Liang et al., 2014). A fish’s life history is usually affected by natural and artificial selection. Selection can induce different probabilities of reproduction, and individual fish can give a different genetic contribution to the next generation (Palkovacs et al., 2012). Genetic differences cause changes in fish biological characteristics, such as size at age, age structure, and growth rate (Hauser et al., 2002; Law, 2007; Liang et al., 2008; Sun et al., 2013). The time scale of natural selection is very long, but that of artificial selection, such as fish breeding and fishing, is shorter. At present, the fisheries management emphasizes on capturing big fish and saving small ones. Many fisheries policies have been made, such as small-size fishing gear, mini-mum ratio of maturation, and minimum capture size or weight (Salas and Gaertner, 2004; Rijnsdorp et al., 2012). These policies are related to changes of biological characteristics, individual miniaturization, and early maturation. If fish minimization cannot be achieved, fisheries resources only increase in biomass rather than recover biological characteristics. Recent studies have shown that fishing gear can affect the evolution of fish population (Jorgensen et al., 2007; Sun et al., 2013, 2015). The causes of fish miniaturization and its life history have been investigated (Vainikka and Hyvarinen, 2012). These investigations aimed to develop a theoretical explication of the cause of the changes in biological characteristics of fish populations. A number of experimental evidences support the idea that fishing may cause evolutionary changes in harvested populations (Conover and Baumann, 2009; Pauli and Heino, 2014).
Industrial fishing is identified as a cause of life-history changes in many harvested stocks mainly because of high fish mortality and damage in stock structure. Trawling has its special selectivity; it is usually used in commercial fishing in China. In this research, the test of time-dependent probability of significance (TDPS) and sensitivity were used to explore the effects, such as exploitationrate of fish, mesh size, and heritability, of long-term trawling. Trawling model can simulate the changes of traits by long-term fishing and halt trawling. The simulation results of hairtail in the East China Sea have confirmed that body length at a certain age becomes shorter and younger individuals are dominant after fishing. In commercial fishing, exploitation rate and mesh size are the important measure of impact parameter. In this study, the test of TDPS and sensitivity of exploited stock aim specifically to assess the relative importance of each factor. The trawl fishing model can simulate fishing pressure to induce the changes of fish mean body length (Sun et al., 2015). The impacts of exploitation rate, mesh size, and heritability with long-term trawling and without fishing are crucial. The impact of fishing pressure is not a single linear relation; other influences are also considered. Therefore, accurate methods of quantitative and qualitative analyses are required. These methods can confirm whether fishing pressure affects fish stock and determine the significance of the impact. Exploitation rate, mesh size, and heritability are important parameters that can induce the changes of traits in exploited fish stock. The genetic traits altered by fishing are slow; the changes in biological characteristics are weaker than those caused by fishing selection. These results are consistent with the fisheries status in China (Ren et al., 2001; Shen and Heino, 2014). Therefore, the direction of fishing-induced evolution warrants considerable attention in China and successful fish stock management can contribute to sustainable fisheries.
2 Materials and Methods
2.1 Trawling Model and Sample
The trawling model adopted has been described in previous publications and employed for a simulation of trawl selectivity-induced evolution effects (Sun et al., 2013 and 2015). In this study, fish stock samples were used in the simulation, and individual body length was employed as research object. Information of the initial fish stock was based on the 1960 hairtail’s data in the East China Sea with an initial distribution shown in Fig.1. The biological characteristics are established from the previous fish stock with the maximum age of 5 years old. Individual body length at each fishing year and each age data can be obtained from the simulation model conducted between 1960 and 2000 in the East China Sea (Sun et al., 2015). This model is a nonlinear statistical analysis with a random error term for individual body length of the next generation. For the purpose of studying the effects of fishing on group characteristics, a series of experiments was carried out (Sun et al., 2015), such as tests for different exploitation rates (E), heritability for the first generation (h2), and mesh size (Ms). The sample used was the mean body length of the group instead of individual body length to test the effects on a group scale rather than an individual scale. For the effects of E, three contrast tests were conducted with different E set as 20%, 40%, and 60%, and each contrast test had three parallel tests. The same approach was used for h2and Ms, with h2set as 0.3, 0.5, and 0.7 and Ms set as 4.4 cm, 5.4 cm, and 6.4 cm.
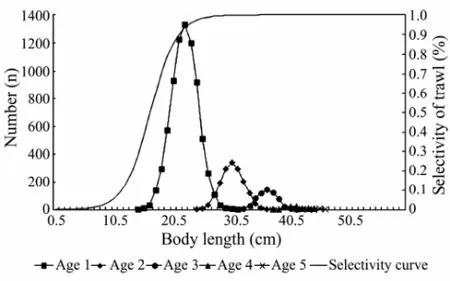
Fig.1 Distribution of body length of the initial hairtail (Trichiurus lepturus) population and selectivity curve of trawl.
2.2 Data Analysis
Sufficient evidence showed the effects of E, h2, and Ms on the fish stock with fishing pressure represented with these parameters. Through fishing years, the fish stock data were different from the initial condition. However, whether the influence of the parameters is significant, the sensitivity of these parameters through fishing years must be studied. Therefore, TDPS testing method was adopted to study the significance of the parameters through fishing years. Sensitivity analysis was also performed.
2.3 TDPS Testing Method
Fig.2 Mean TDPS of each age with different exploited rate by 40 years fishing (TDPS is time-dependent probability of significance, the value of TDPS is from 0 to 1. 0 represents no impact of sample in fish population, and 1 represents important impact).
The idea of TDPS came from the conception of oneway ANOVA. The traditional one-way ANOVA method tests whether the influence of the only parameter for the system is significant. For a long-term simulation, the state of the system not only includes the parameters E, h2, or Ms but also the function of simulation time (t). An index is necessary to test the significance of parameters (E, h2,or Ms) for the time-dependent random variable, which is the mean body length of ages. In a one-way ANOVA, a series of contrast tests is conducted for only one parameter with different values. If the influence of this parameter is significant, then the distributions of contrast tests are different, and the probability determines whether the distributions are the same or not. Thus, this probability can be used to measure significance; it is also the unit measure called TDPS in this study. However, for a time-dependent simulation, a special treatment is needed because parameter t always exists. This special treatment is shown in Fig.2. For a fixed time t0, only one parameter with different values exists. At any fixed time, the progress of one-way ANOVA can be used to obtain the TDPS value (Tian, 2013; Andrade et al., 2015).
2.4 Sensitivity Testing Method
The TDPS testing method explore whether fishing pressure affects body length at certain ages and whether each factor has different degrees of impact. Fish population may have different sensitivities for different parameters. The TDPS values can be very high, but the sensitivity can be low. Given this reason, the sensitivity testing method was used to examine the sensitivity of exploitation rate, heritability for the first generation, and mesh size.
Non-dimensional sensitivity analysis (Li et al., 2005) is expressed as follows:
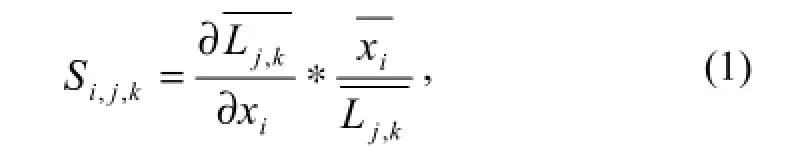
where S is sensitivity,Lis the fish mean body length, and x is the research variable. The span of i, j, k respresents the kinds of parameters, different ages of fish, and fishing time stepsand k is from 1 to 40.
3 Result
Comparing the outcome of this analysis with the ones from previous studies, the results and operation of the fishing model are ideal. The model can achieve accurate simulation by real trawling pressure and induce the changes of mean body length by long-term trawling with different fishing parameters. With the increasing exploitation rate and heritabality, and decreasing mesh size, a more important impact to fish mean body length is evident in previous results (Sun et al., 2013, 2015). The impact cannot recover to the initial condition once trawling stops. Therefore, the fishing parameters and biological characteristics are important influences on the fish stock. The impact of these fishing parameters are not a simple linear progress because the fish stock adapts to the living enviroment. Qualitative and quantitative analyses should be conducted to research into the impact of different factors. The results of these analyses can prove whether the parameters induce sensitivity changes. The impacts of E, Ms, and h2by trawling are significant to each age (see Figs.2–7).
Fig.3 The changes of TDPS with different exploited rate by 40 years fishing.

Fig.4 Mean TDPS of each age with different mesh size by 40 years fishing.
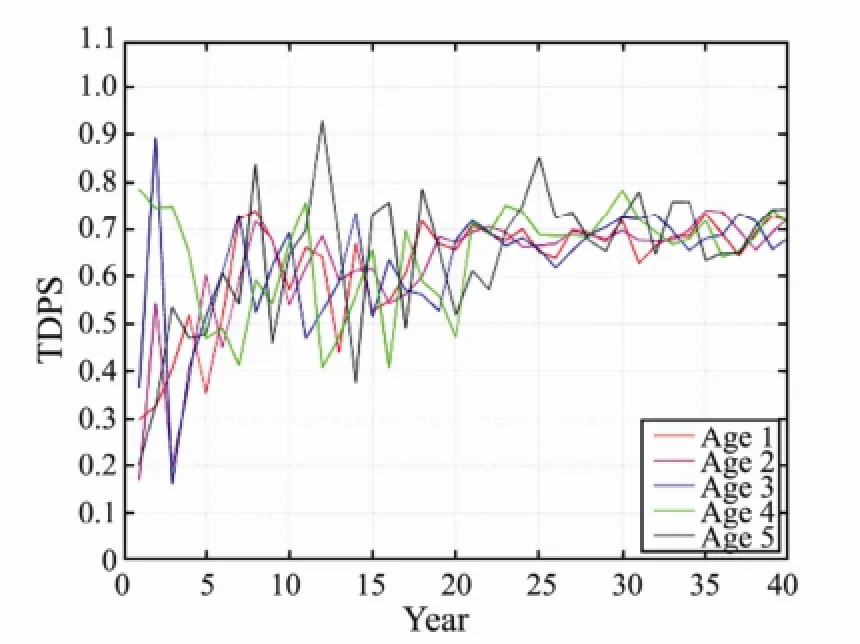
Fig.5 The changes of TDPS with different mesh size by 40 years fishing.
3.1 Results of TDPS Testing
As shown in Figs.2 to 7, the impact of E was very significant to fish individuals at ages 1 to 5, and the TDPS value was always 1 (Figs.2–3). Meanwhile, the impact of the Ms and h2was significant to fish individuals at ages 1 to 5, but the impact to younger ones was more significant by fishing. The significance of Ms and heritability tended to be stable under long-term fishing. Particularly, the impact of Ms showed a 5-year cycle in fishing. The tendency, however, was not more significantthan that of heritability (Figs.4–7). This change is consistent with the fish life history, which is a 5-year cycle.

Fig.6 Mean TDPS of each age with different heritability by 40 years fishing.
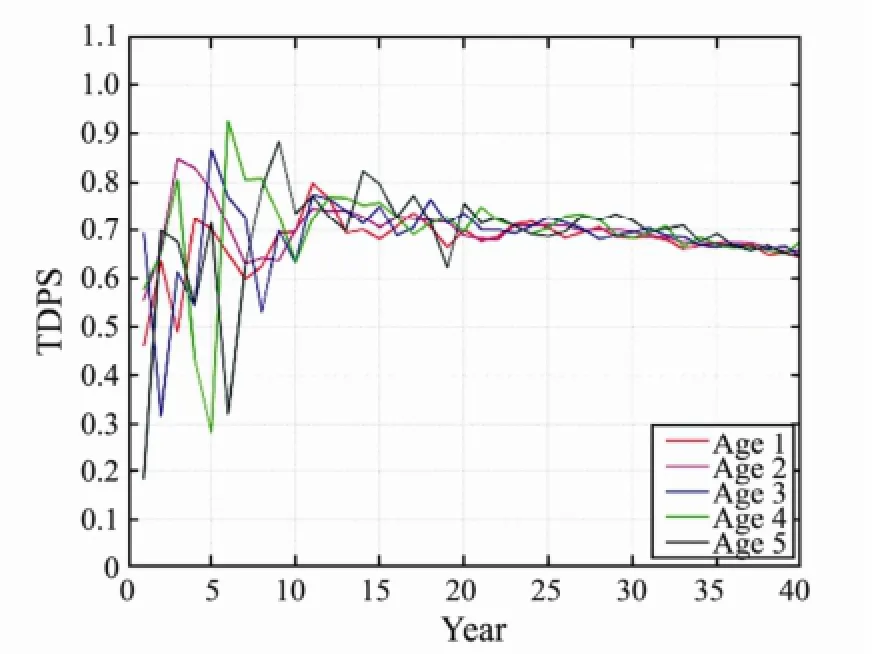
Fig.7 The changes of TDPS with different heritability by 40 years fishing.
3.2 Results of Sensitivity Testing
The test results of TDPS of E, Ms, and h2were evident with fishing pressure, and these factors have a very important impact to fish stocks. However, the sensitivity can directly measure the speed of system changes, and the sensitivity analyses of E, Ms, and h2were used to explore the degree of impact of fishing parameters on the changes. The sensitivity values of E and Ms showed the same tendency as those of fishing pressure; the values were gradually increasing and having a stead trend (Figs.8, 9). The sensitivity of each age group exhibited the same tendency. The tendency of the sensitivity of h2was not initially regular, but showed an increasing tendency until it became stable (Fig.10).
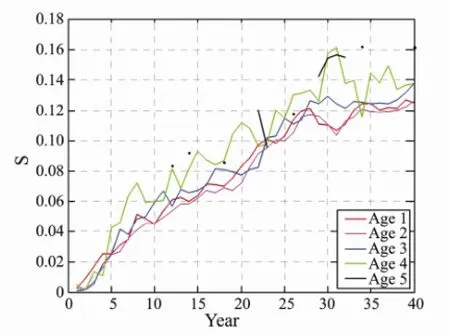
Fig.8 The sensitivity of each age with different expolited rate by 40 years fishing (S is sensitivity).
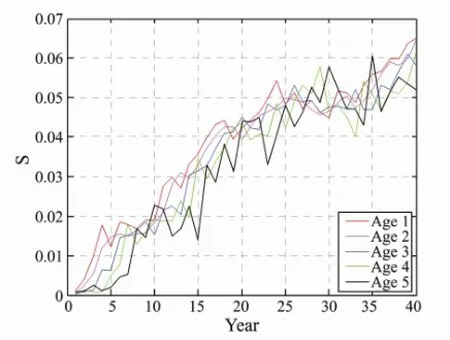
Fig.9 The sensitivity of each age with different mesh size by 40 years fishing.
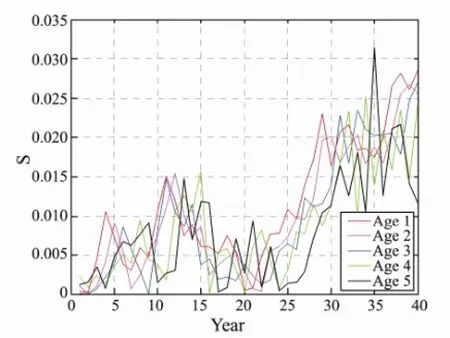
Fig.10 The sensitivity of each age with different heritability by 40 years fishing.
4 Discussion
The trawling model can determine the impact of exploitation rate and mesh size, and the sensitivity tendency was similar to long-term fishing. The changes of sensitivity showed a gradual growth until they become stable. However, the change of heritability sensitivity was different from the exploitation rate and mesh size. The results of this study provide a detailed analysis of fishing parameter, which show influences on biological characteristics by long-term fishing pressure. Although this research method is based on conventional ANOVA, the application is not similar. The analysis used the probability of test to indicate the significance of test variables. Exploitation rate, mesh size, and heritability are important factors that influence the exploited fishing stock based on the results. The changes of TDPS can determine the internal rule and verify which parameter is the most important. A range of statistical methods is available for quantifying changes in environmental impact over time (Parket al., 2011). In this study, the results indicate that fishing and biological factors are important. Some of the parameters can be controlled in fish-captured progress, and many fisheries management methods have policies to control the captured size, total biomass, sex ratio, and so on. However, if the fish stock is exploited in this manner, how can this exploitation change fisheries resources. Many studies indicate that the evolutionary response can be reversed by changing the fishing capacity and size selectivity (Mollet et al., 2007). Size selectivity can induce evolutionary response that cannot be recovered (Law, 2007; Jorgensen et al., 2009). All such academic work agrees that the biological characteristics of exploited fish stock cannot be recovered with the present fishing policy. Size selectivity is not a good method to exploit fish stocks. An example is the Norwegian spring-spawning herring. Rebuilding its stock took 30 years after its collapse in the 1960s (Toresen and Ostvedt, 2000).
At present, a series of measures to improve management of marine resources in China is available, including the fishing license system, the minimum mesh size regulation, the ‘Double Control’ system, summer fishing moratoria, fishing catch limit regimes, and so on. Under long-term fishing pressure, which measure is good for sustainable fisheries? The results of this study indicate that the exploitation rate and mesh size are important factors to fish stock. Using hairtail in the East China Sea as an example, the data of this fish stock show miniaturization under long-term trawling (Lin et al., 2006; Sun et al., 2015). Thus, minimum mesh size is important to fish stocks; the size depends on the fishing gear as well as fishing area based on Article 30 of the Fisheries Law of the People’s Republic of China (2004). Meanwhile, heritability is also an important influence on fish stock. Sustainable fishing pressure can influence fish stock in short time period, but the impact of heritability is a long-term factor. Heritability can induce the evolution of fish biological characteristics for a long time, but recovery is impossible once fishing pressure stops or fishing factors change. Therefore, whether sustainable minimum mesh size in fishing is good for exploited fish stock or not needs further research. The long-term effect of each factor’s trend can be observed in our study. Previous studies can only obtain the average magnitude of impact from fishing pressure, but it cannot verify the degree of influence of the impact. The present study obtained the trend of the changes as well as the change rule. The changes showed a fluctuation cycle in 5-year fishing, which is consistent with the fish life history that is a 5-year cycle (Figs.8–10). The sensitivity of fishing parameters affected the fish stock; it initially showed an increasing significance in 30-year trawling, and then the trend became stable. This trend is the same as that in real fisheries, minimum mesh size, and fishing catch limit policy. These management policies can initially affect the fish stock significantly, and then the trend becomes stable. Thus, the impact can affect the exploited fish stock in a short time. Whether the effect is good or not and whether it can contribute to sustainable fisheries need further study. The results also indicate that the genetic variation cannot cause regular variation trends by 40-year trawling. Therefore, the impact of genetic variation is an internal mechanism, and long-term changes should be considered.
Acknowledgements
We acknowledge the financial support from the National Natural Science Foundation of China (No. 31402 350) and Special Fund for Agro-scientific Research in the Public Interest (No. 201203018).
Andrade, L. S., Frameschi, I. F., Costa, R. C., Castilho, A. L., and Fransozo, A., 2015. The assemblage composition and structure of swimming crabs (Portunoidea) in continental shelf waters of southeastern Brazil. Continental Shelf Research, 94: 8-16.
Conover, D. O., and Baumann, H., 2009. The role of experiments in understanding fishery-induced evolution. Evolutionary Applications, 2: 276-290.
Dieckmann, U., and Heino, M., 2007. Probabilistic maturation reaction norms: Their history, strengths, and limitations. Marine Ecology Progress Series, 335: 253-269.
Dunlop, E. S., Heino, M., and Dieckmann, U., 2009. Eco-genetic modeling of contemporary life-history evolution. Journal of Applied Ecology, 19: 1815-1834.
Enberg, K., Jorgensen, C., Dunlop, E. S., Heino, M., and Dieckmann, U., 2009. Implications of fisheries-induced evolution for stock rebuilding and recovery. Evolutionary Applications, 2: 394-414.
Enberg, K., Jorgensen, C., Dunlop, E. S., Varpe, O., and Boukal, D. S., 2012. Fishing-induced evolution of growth: Concepts, mechanisms and the empirical evidence. Marine Ecology, 33: 1-25.
Fenberg, P. B., and Roy, K., 2007. Ecological and evolutionary consequences of size-selective harvesting: How much do we know? Molecular Ecology, 17: 209-220.
Hauser, L., Adcock, G. J., Smith, P. J., Ramirez, J. H. B., and Carvalho, G. R., 2002. Loss of microsatellite diversity and low effective population size in an overexploited population of New Zealand snapper (Pagrus auratus). Proceedings of the National Academy of Sciences of the United States of America, 99: 11742-11747.
Hutchings, J. A., and Fraser, D. J., 2008.The nature of fisheries and farming-induced evolution. Molecular Ecology, 17: 294-313.
Jorgensen, C., Enberg, K., and Dunlop, E. S., 2007. Managing evolving fish stocks. Science, 318: 1247-1248.
Jorgensen, C., Ernande, B., and Fiksen, O., 2009. Size-selective fishing gear and life history evolution in the Northeast Arctic cod. Evolutionary Applications, 2: 356-370.
Kuparinen, A., and Merila, J., 2007. Detecting and managing fisheries-induced evolution. Trends in Ecology and Evolution, 22: 652-659.
Laugen, A., Engelhard, G. H., Whitlock, R., Arlinghaus, R., Dankel, D. J., Dunlop, E. S., Eikeset, A. M., Enberg, K., Jorgensen, C., Matsumura, S., Nussle, S., Urbach, D., Baulier, L., Boukal, D. S., Ernande, B., Johnston, F. D., Mollet, F., Pardoe, H., Therkildsen, N. O., Uusi-Heikkila, S., Vainikka, A., Heino, M., Rijnsdorp, A. D., and Dieckmann, U., 2014. Evolutionary impact assessment: Accounting for evolutionaryconsequences of fishing in an ecosystem approach to fisheries management. Fish and Fisheries, 15: 65-96.
Law, R., 2007. Fisheries-induced evolution: Present status and future directions. Marine Ecology Progress Series, 335: 271-277.
Li, S. Y., Zhang, X. W., and Xing, L. N., 2005. The methods of dynamic selecting measures based on sensitivity analysis. Computer Simulation, 22 (3): 120-136 (in Chinese with English abstract).
Liang, Z. L., Sun, P., Tang, Y., Huang, L., Lu, Q., Liu, C., and Liu, Q., 2008. Numeric simulation on impact of trawl mesh selectivity on biological characters of fish population. Oceanologia et Limnologia Sinica, 39: 487-493 (in Chinese with English abstract).
Liang, Z. L., Sun, P., Yan, W., Huang, L. Y., and Tang, Y. L. 2014. Significant effects of fishing gear selectivity on fish life history. Journal of Ocean University of China, 13 (3): 467-471.
Lin, L., Zheng, Y., Cheng, J., Liu, Y., and Ling, J., 2006. A preliminary study on fishery biology of main commercial fishes surveyed from the bottom trawl fisheries in the East China. Marine Science, 30: 21-25 (In Chinese with English abstract).
Mollet, F. M., Kraak, S. M. B., and Rijinsdorp, A. D., 2007. Fisheries-induced evolutionary changes in maturation reaction norms in North Sea sole Solea solea. Marine Ecology-Progress Series, 351: 189-199.
Palkovacs, E. P., Kinnison, M. T., Correa, C., Dalton, C. M., and Hendry, A. P., 2012. Fates beyond traits: Ecological consequences of human-induced trait change. Evolutionary Applications, 5: 237-254.
Park, S., Chu, P. C., and Lee, J. H., 2011. Interannual-tointerdecadal variability of the Yellow Sea Cold Water Mass in 1967-2008: Characteristics and seasonal forcings. Journal of Marine Systems, 87: 177-193.
Pauli, D., and Heino, M., 2014. What can selection experiments teach us about fisheries-induced evolution? Biological Journal of the Linnean Society, 111: 485-503.
Ren, Y., Gao, T., Liu, Q., and Xue, Y., 2001. Study on the population structure and reproduction of Pseudosciaena polyactis in southern yellow sea. Transactions of Oceanology and Limnology, 1: 41-46 (in Chinese with English abstract).
Rijnsdorp, A., Overzee, H., and Poos, J., 2012. Ecological and economic trade-offs in the management of mixed fisheries: A case study of spawning closures in flatfish fisheries. Marine Ecology Progress Series, 447: 179-194.
Salas, S., and Gaertner, D., 2004. The behavioural dynamics of fishers: management implications. Fish and Fisheries, 5: 153-167.
Shen, G. M., and Heino, M., 2014. An overview of marine fisheries management in China. Marine Policy, 44: 265-272.
Sun, P., Liang, Z. L., Huang, L. Y., Tang, Y. L., and He, X., 2013. Relationship between trawl selectivity and fish body size in a simulated population. Chinese Journal of Oceanology and Limnology, 31 (2): 327-333.
Sun, P., Liang, Z. H., Yu, Y., Tang, Y. L., Zhao, F. F., and Huang, H. L., 2015. Trawl fishing induced evolutionary effects on age structure and size at age of hairtail (Trichiurus lepturus) in East China Sea, China. Journal of Applied Ichthyology, DOI: 10.1111/jai.12774.
Tian, B., 2013. Single factor analysis variance and its application. Yinshan Academic Journal, 27 (2): 24-27 (in Chinese with English abstract).
Toresen, R., and Ostvedt, O. J., 2000. Variation in abundance of Norwegian spring-spawning herring (Clupea harengus, Clupeidae) throughout the 20th century and the influence of climatic fluctuationsz. Fish and Fisheries, 1: 231-256.
Vainikka, A., and Hyvarinen, P., 2012. Ecologically and evolutionary sustainable fishing of the pikeperch Sander lucioperca: Lake Oulujarvi as an example. Fisheries Research, 113: 8-20.
(Edited by Ji Dechun)
(Received February 5, 2015; revised May 6, 2015; accepted May 14, 2015)
J. Ocean Univ. China (Oceanic and Coastal Sea Research)
DOI 10.1007/s11802-015-2885-5
ISSN 1672-5182, 2015 14 (5): 835-840
http://www.ouc.edu.cn/xbywb/
E-mail:xbywb@ouc.edu.cn
* Corresponding author. E-mail: sunbird1103@sina.com
 Journal of Ocean University of China2015年5期
Journal of Ocean University of China2015年5期
- Journal of Ocean University of China的其它文章
- Seasonal Dynamics of Turbidity Maximum in the Muthupet Estuary, India
- Decadal Variability of Global Ocean Significant Wave Height
- An Observational and Modeling Study of Extratropical Transition of Hurricane Sandy in 2012
- The Nonlinear Bifurcation and Chaos of Coupled Heave and Pitch Motions of a Truss Spar Platform
- Beach Morphology and Coastline Evolution in the Southern Bohai Strait
- Comparison of Meiofaunal Abundance in Two Mangrove Wetlands in Tong’an Bay, Xiamen, China