
封育对宁夏盐池县草地植物组成及群落复杂性的影响
2015-03-15 01:37刘小丹张克斌林田苗刘建康
水土保持通报 2015年4期
刘小丹, 张克斌, 林田苗, 刘建康
(1.中国环境管理干部学院, 河北 秦皇岛 066102; 2.北京林业大学 水土保持学院
水土保持与荒漠化防治教育部重点实验室, 北京 100083; 3.水利部 水土保持植物开发管理中心, 北京 100038)
封育对宁夏盐池县草地植物组成及群落复杂性的影响
刘小丹1,2, 张克斌2, 林田苗3, 刘建康2
(1.中国环境管理干部学院, 河北 秦皇岛 066102; 2.北京林业大学 水土保持学院
水土保持与荒漠化防治教育部重点实验室, 北京 100083; 3.水利部 水土保持植物开发管理中心, 北京 100038)
摘要:[目的] 定量评价封育对宁夏盐池县沙化草地植物组成及群落复杂性的影响,为半干旱区草地植被恢复与管理提供理论依据。 [方法] 对3种封育类型老、新封育区和对照区在2003—2013年样线与样方结合法进行植被调查,并用群落总复杂性、无序结构复杂性、有序结构复杂性3种指数来分析其复杂性特征。 [结果] 老封育区由黑沙蒿(Artemisia ordosica)和刺沙蓬(Salsola ruthenica)群落逐渐演替为阿尔泰狗娃花(Heteropappus altaicus)群落,新封育区和对照区主要是黑沙蒿群落;新封育区和对照区的植被生物量与盖度大多年份高于老封育区;封育可增加无序与有序结构复杂性,进而增加群落总复杂性。 [结论] 长期封育降低了植被数量特征值;随封育年限的延长,群落复杂性指数约以5 a左右为波动周期,说明适宜封育年限为5 a。
关键词:封育草地; 植物组成; 群落复杂性
草地是陆地生态系统重要组成部分,由于自然与人为等因素的影响,草地退化严重。在草地恢复过程中,封育被认为是最有效的手段之一。目前在干旱半干旱区封育对草地植被恢复、演替状况等的研究仍是热点,其中通过研究植被的群落特征及物种多样性是比较常用的手段,但这些方法很难定量描述植被群落内物种相互作用,自组织及有序性方面的特征,而群落复杂性的研究可弥补多样性研究的不足,复杂性除了包括群落中各个尺度的多样性以外,还包括这些多样的组分之间的联结和作用[1]。自从Anand[2]提出基于Huffman编码的群落复杂性测度计算方法以来,国内此领域研究相对较少[1,3-10],用于对草地生态系统植物群落复杂性研究则更少。因此,本研究选取宁夏自治区盐池县典型封育草地为研究对象,基于多年植被调查数据,分析封育对草地植物组成、生物量与盖度数量特征及群落复杂性的影响,通过数据分析,探讨植物组成及群落复杂性指数的变化规律,旨在为半干旱区沙化草地的恢复与利用提供理论依据。
1材料与方法
1.1 研究区概况
盐池县位于宁夏回族自治区东部、毛乌素沙漠的南缘,坐标为北纬37°04′—38°10′,东经106°30′—107°41′。该县总面积8 661.3 km2。在地理位置上属于典型的过渡地带(自南向北从黄土高原向毛乌素沙地过渡)。县内地势南高北低,南部为黄土丘陵区,中部为鄂尔多斯缓坡丘陵区。气候类型上,属中温带大陆性气候,年均气温为8.1 ℃;年降水量仅250~350 mm。土壤类型以灰钙土为主。常见植物种类以旱生和中旱生类型为主。
1.2 样地设置与采样方法
典型封育草地选择盐池县北部荒漠草原区柳杨堡封育区,3种处理方法:老封育区为90年代初封育示范区,新封育区为2002年开始封育的区域,对照区2002年也采取了封育措施,但由于其位于新封育区与周边公路之间的区域,因而受到一些人为干扰和放牧的影响。3块样地距离相近,自然条件基本相同。调查时间为2003—2013年7—8月份植物生长高峰期,采用样线和样方结合法,由于调查区域内主要以小半灌木和多年生草本植物为主,故每块样地每隔30 m左右布设1 m×1 m样方,3块区域各布设10个样方,研究期内共调查330个样方,调查内容包括植物名称、物种数、盖度、株数、高度、生物量等。
1.3 指标计算及数据处理
1.3.1重要值计测作为度量植物种群的重要指标,综合反映了植物物种在群落中的重要程度,故综合考虑植物生物、生态学特性,同时结合研究区生境特征,选用5个植物特征值进行重要值的计算[11-12]。

(1)
式中:某种植物的某项指标相对值=该样方内该植物的某项测定值/该样方内所有植物的该项测定值之和×100%。
1.3.2复杂性指数物种相互作用(主要是竞争)是群落结构组建的主要因素,那么群落自组织或有序性就间接反应了群落内物种相互作用的大小,因此复杂性测度就可作为群落内物种相互作用总量的测度。有序结构复杂性可理解为构成一个群落结构的规则或规律的最小描述的长度,而无序结构复杂性是关于群落结构的随机部分的最小描述的长度。由于Ha(S)熵是平均码长的下限,而L(S)可以超过Ha(S)但不能小于Ha(S)。正因为满足这个关系,才可将L(S)作为总复杂性的测度,将Ha(S)作为无序结构复杂性的测度。以最小剩余码的平均码长和12阶Rényi熵的差作为有序结构复杂性△S的度量指标[1,6]。
1.3.3复杂性数据处理通过调查建立研究区草地植被数据库,计算不同封育区植物物种重要值矩阵。群落复杂性计算先由Matlab 7.0编写Huffman编码程序,把相对重要值转换为二元码,统计植物的二元码码字长度,利用公式计算复杂性指数。
2结果与分析
2.1 不同封育年限各样地植物组成变化
通过植被调查统计研究表明,老封育区在2003—2005年优势种为黑沙蒿,伴生植物有狗尾草(Setariaviridis)、赖草(Anaeurolepidiumsecalium)、刺沙蓬;2006—2007年优势种过渡到刺沙蓬,伴生种有黑沙蒿(Artemisiaordosica)、阿尔泰狗娃花(Heteropappusaltaicus),在此期间植物组成结构发生演替;2008—2012年优势种变为阿尔泰狗娃花,伴生种包括丝叶山苦荬(Ixerischinensis)、草木樨状黄芪(Astragalusmelilotoides)、刺沙蓬、赖草;到2013年优势种为刺沙蓬与赖草。总的演替趋势为以黑沙蒿和刺沙蓬为优势种的植物群落逐步演替到以阿尔泰狗娃花为优势种群落。新封育区在2003—2012年优势种为黑沙蒿,伴生植物有苦豆子(Sophoraalopecuroides)、刺沙蓬;2013年优势种为阿尔泰狗娃花与黑沙蒿;说明在新封育区黑沙蒿占据着绝对的优势地位。对照区在2003—2006年优势种为黑沙蒿,伴生种主要为苦豆子;2007—2013年优势种仍为黑沙蒿,伴生种逐渐由苦豆子变为刺沙蓬。
2.2 不同封育年限各样地生物量及植被覆盖变化
生物量、盖度指标较大程度上能反映植被生长状况。从表1可知,3块样地区域的植被生物量、盖度变化趋势基本相似。新封育区和对照区的植被生物量、盖度大多年份高于老封育区,一方面随着封育年限的延长,老封育区大量生物结皮产生,结皮抑制降雨的入渗,同时结皮与植物争夺生长资源,结皮大量存在抑制植被生长,建议对老封育区进行轻度干扰,以保持其较高的生物量与覆盖度;一方面封育区植被以多年生为主,植物根部变的发达,地下生物量增加,而地上新增生物量相应减小;一方面植被恢复的过程就是群落的建立和演替过程,核心是物种的更替,随着优势种的壮大,将占据封育区大量的资源空间,使其特化种逐渐丧失生存空间,导致植被盖度与生物量降低;另外,由于研究区地理位置较为特殊,生态环境脆弱,植被随气候的波动特征十分明显,尤其是植被生长季降水量,降水的波动变化使得植被生物量与盖度也发生波动增减。
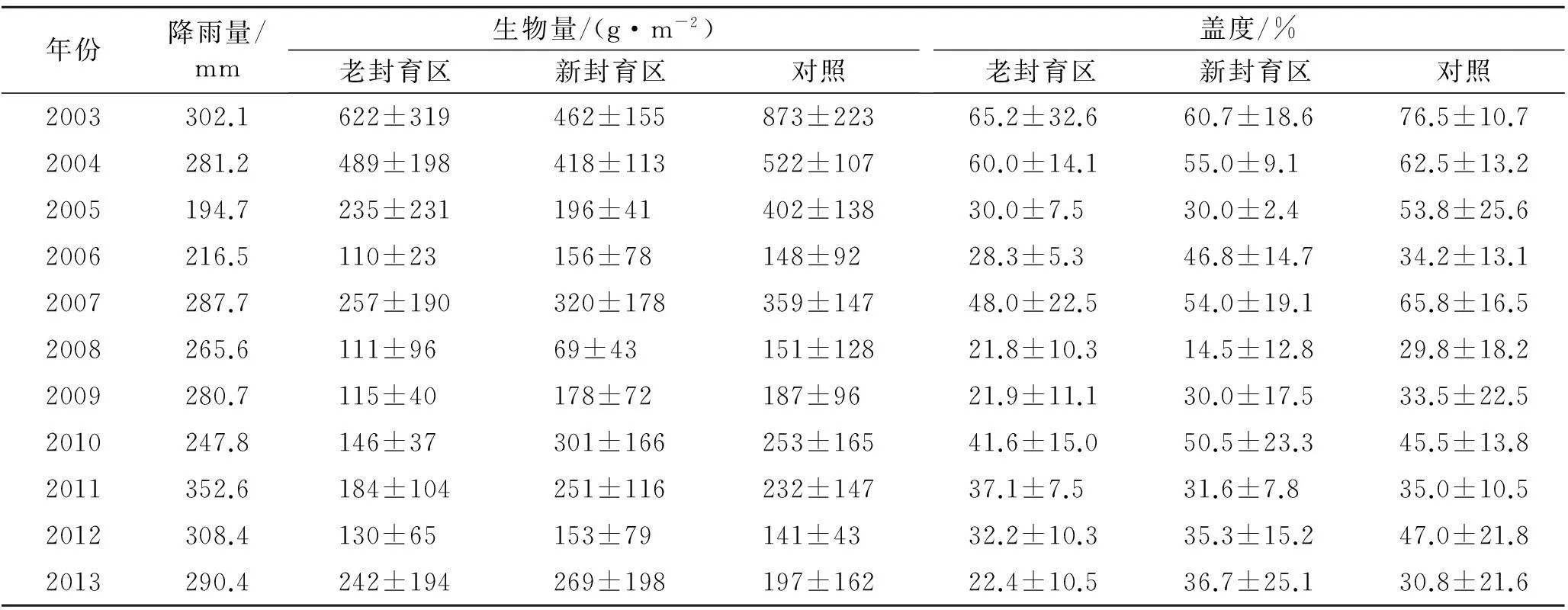
表1 研究年份降雨量、各样地生物量及植被盖度变化
注:生物量和盖度数据为平均值±标准差。
2.3 不同封育年限样地的群落复杂性变化特征
由表2可知,老封育区、新封育区总复杂性指数值大部分比对照区指数值高,这是由于采取封育措施避免人畜干扰,使得草原群落植被得以恢复,封育增加了群落总复杂性;基于无序的复杂性指数波动变化与群落总复杂性指数波动基本一致,封育有利于增加群落无序结构复杂性;老封育区有序结构复杂性指数值整体偏高,新封育区次之,对照区偏低,说明封育也提高了有序结构复杂性。另外,随着封育年限的延长,3种复杂性指数的高峰值与低峰值有规律性的波动变化,波动频率约5 a左右。
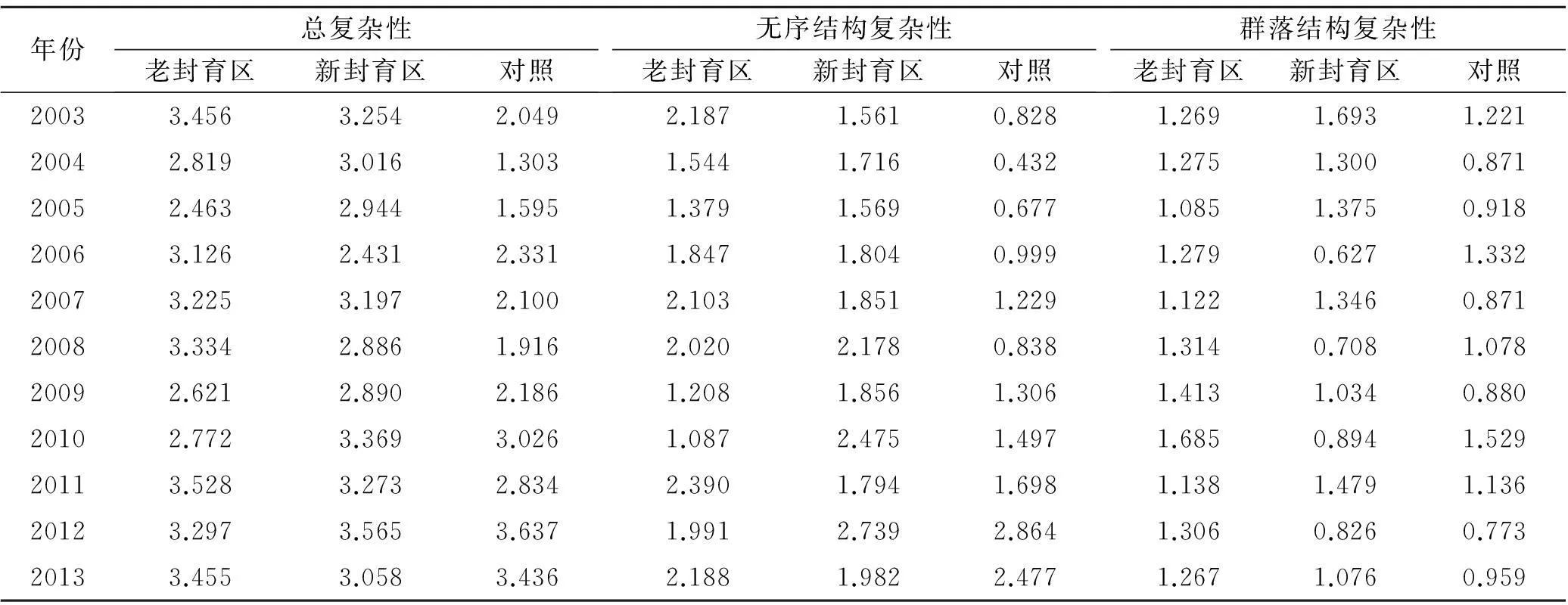
表2 研究年份各样地群落复杂性指数变化
3讨论与结论
(1) 植物组成的研究表明,老封育区总的演替趋势为以黑沙蒿和刺沙蓬为优势种的植物群落逐渐演替到以阿尔泰狗娃花为优势种群落。新封育区黑沙蒿群落占据着绝对的优势地位。对照区黑沙蒿优势种与苦豆子伴生种逐渐变为黑沙蒿优势种与刺沙蓬伴生种群落。
(2) 围栏封育是促进退化草地植被恢复的有效手段。相关的封育研究[13-14]表明,一定封育时间内,植被盖度、生物量增加,但围封时间过长不利于维持较高植被数量特征值。本研究表明,不同封育类型区的植被生物量、盖度变化趋势基本相似,由于生物结皮、种间竞争、植物特性、气候因素等多因素的影响,不同封育年限下,植被盖度、生物量发生波动变化。新封育区和对照区的植被生物量与盖度大多年份高于老封育区,调查表明老封育区分布有大量生物结皮,根据卢晓杰等[15-16]土壤结皮试验研究表明结皮盖度与植被盖度、土壤入渗深度呈极显著负相关。结皮由相互促进生长转变为相互竞争的关系,结皮抑制了植被的生长;另外,由于生物结皮的存在,阻止了水分向深层渗透,植被长势变差,植被盖度、生物量等指标值降低。
(3) 研究表明,长期封育不是合理的,在草地管理中要确定合理的封育年限。蒋德明等[13]在科尔沁沙地封育研究表明,适度的封育年限6~10 a。程积民等[17]对黄土高原半干旱区封育研究认为长期封育对草地物种更新和生态系统稳定性有负面影响,提出封育10~15 a后需进行合理利用。白于等[18]的研究表明,适宜封育时间为9 a。而宁夏典型封育草地的相关研究[19-20]表明适宜封育年限为5 a。本研究表明,随着封育年限的延长,群落复杂性波动频率约5 a左右,说明封育周期以5 a左右为宜。
(4) 目前围栏封育对生物多样性影响的报道很多,但是对群落复杂性影响的研究极少见。李镇清[6]对中国东北样带植物群落复杂性研究表明,干扰相对较少的围封草地群落具有较高的群落复杂性。本研究利用群落复杂性测度来研究封育区植被群落,表明封育确实增加无序结构复杂性与有序结构复杂性,进而增加群落总复杂性。
[参考文献]
[1]李镇清.分离生态变化中的物种相互作用组分[J].生物数学学报,2001,16(3):320-333.
[2]Anand O L. Complexity in plant communities: The notion and quantification [J]. Journal of Theoretical Biology, 1996,179(2):179-186.
[3]金森.群落结构复杂性的测度方法研究进展[J].植物生态学报,2006,30(6):1030-1039.
[4]金森.基于Huffman编码的群落结构复杂性[J].植物生态学报,2007,31(6):1154-1160.
[5]安丽娟,朱志红,王孝安,等.子午岭马栏林区主要植物群落复杂性[J].东北林业大学学报,2008,36(12):28-31.
[6]李镇清.中国东北样带(NECT)植物群落复杂性与多样性研究[J].植物学报,2000,42(9):971-978.
[7]覃林,余世孝.森林群落复杂性分析:以广东黑石顶森林为例[J].生物多样性,2004,12(3):354-360.
[8]王宇超,王得祥.佛坪自然保护区植物群落物种多样性和复杂性的海拔格局研究[J].西北植物学报,2013,33(1):169-176.
[9]尤海梅,藤原一绘.徐州低山丘陵区侧柏人工林群落的复杂性[J].南京林业大学学报:自然科学版,2011,35(6):34-38.
[10]冶民生,吴斌,关文彬,等.岷江源区植物群落复杂性研究[J].干旱区资源与环境,2009,23(8):174-179.
[11]李瑞,张克斌,刘云芳,等.西北半干旱区湿地生态系统植物群落空间分布特征研究[J].北京林业大学学报,2008,30(1):6-13.
[12]Huffman D A. A method for the construction of minimum-redundancy codes [J]. Proceedings of the Ire, 1952,40(9):1098-1101.
[13]蒋德明,苗仁辉,押田敏雄,等.封育对科尔沁沙地植被恢复和土壤特性的影响[J].生态环境学报,2013,22(1):40-46.
[14]刘凤婵,李红丽,董智,等.封育对退化草原植被恢复及土壤理化性质影响的研究进展[J].中国水土保持科学,2012,10(5):116-122.
[15]卢晓杰,张克斌,李瑞.北方农牧交错带生物结皮的主要影响因子探讨[J].水土保持研究,2007,14(6):1-4.
[16]卢晓杰,李瑞,张克斌.农牧交错带地表覆盖物对土壤入渗的影响[J].水土保持通报,2008,28(1):1-5.
[17]程积民,井赵斌,金晶炜,等.黄土高原半干旱区退化草地恢复与利用过程研究[J].中国科学:生命科学,2014,44(3):267-279.
[18]白于,苏纪帅,程积民.黄土区不同封育时期天然草地的根系生物量分布[J].草业科学,2013,30(11):1824-1830.
[19]赵菲,谢应忠,马红彬,等.封育对典型草原植物群落物种多样性及土壤有机质的影响[J].草业科学,2011,28(6):887-891.
[20]李学斌,陈林,李国旗,等.干旱半干旱地区围栏封育对甘草群落特征及其分布格局的影响[J].生态学报,2013,33(13):3995-4001.
Effects of Enclosing on Vegetation Composition and Community Complexity in Grassland in Yanchi County, Ningxia Hui Autonomous Region
LIU Xiaodan1,2, ZHANG Kebin2, LIN Tianmiao3, LIU Jiankang2
(1.EnvironmentalManagementCollegeofChina,Qinhuangdao,
Hebei066102,China; 2.KeyLaboratoryofSoilandWaterConservationandDesertificationCombatingoftheMinistryofEducation,CollegeofSoilandWaterConservation,BeijingForestryUniversity,Beijing100083,China; 3.PlantDevelopmentandManagementCenterforSoilandWaterConservation,MinistryofWaterResources,Beijing100038,China)
Abstract:[Objective] The effects of the enclosing on vegetation composition and community complexity were examined in grassland of Yanchi County, Ningxia Hui Autonomous Region, in order to provide the theoretical basis for vegetation restoration and grassland management in semi-arid area. [Methods] By quadrat method, the vegetation was investigated from 2003 to 2013 with respect to the impacts of different fencing time on vegetation composition and three kinds of complexity index(total complexity, disorder-based complexity, and structural complexity). Three processing methods of study region were adopted: old enclosure area, new enclosure area and control area. [Results] Artemisia ordosica and Salsola ruthenica communities gradually converted into Heteropappus altaicus community in the old enclosure area, and Artemisia ordosica was dominant species in new enclosure and control area. The biomass and coverage in the old enclosure area were less than those in the new enclosure area and control area; enclosure increased disorder-based complexity and structural complexity, thus which made the total community complexity increase. [Conclusion] The long-term enclosure is not conducive to vegetation recovery. The high peak and low peak values of each complexity index appeared regularly with the cycle of five-year. So, 5-year is appropriate enclosure period for grassland in the semi-arid area.
Keywords:enclosure grassland; vegetation composition; community complexity
文献标识码:A
文章编号:1000-288X(2015)04-0304-04
中图分类号:Q948.1, Q948.15
收稿日期:2014-06-17修回日期:2014-06-23
资助项目:国家自然科学基金项目“半荒漠湿地植被群落时空分布特征研究”(30771764); 国家林业局科技项目(322340)
第一作者:刘小丹(1983—),男(汉族),山西省临县人,博士,讲师,主要从事环境生态工程、生态环境监测方面的研究工作。E-mail:liuxiaodan831121@163.com。
