
不同生境下柔毛淫羊藿的开花物候
2015-03-14 11:31:06谌利民权秋梅黎云祥
西华师范大学学报(自然科学版) 2015年1期
杨 萍,谌利民,权秋梅,黎云祥
(1.西华师范大学西南野生动植物资源保护教育部重点实验室;2.唐家河国家级自然保护区,四川 南充 637009)
不同生境下柔毛淫羊藿的开花物候
杨 萍1,谌利民2,权秋梅1,黎云祥1
(1.西华师范大学西南野生动植物资源保护教育部重点实验室;2.唐家河国家级自然保护区,四川 南充 637009)
用野外定点观测的方法对金城山和唐家河地区的柔毛淫羊藿的开花物候特征以及其与结实率进行了比较究,结果表明:在不同生境下的种群、个体、花序与单花的花期存在差异.种群的开花振幅曲线都呈单峰曲线,金城山柔毛淫羊藿的开花比例60%-70%,开花强度分布范围集中在50%;唐家河柔毛淫羊藿的开花比例40%-45%,开花强度分布范围则为20%-50%.个体水平的开花振幅曲线均呈单峰曲线,具有较高的开花同步指数,表现出一种集中开花的特性.坐果数与物候指数的相关分析表明两个地方的座果数均与开花数和花期存在着显著的相关性.综上所述,柔毛淫羊藿的个体开花物候容易受到地理环境的影响,因此,在环境干扰的选择压力下,为了吸引更多的传粉者来访而达到成功生殖的目的,它便形成了大量集中开花的模式.
柔毛淫羊藿;开花物候;环境因子
开花物候(Flowering phenology)是描述植物生活史及其特征的一个重要表征方式,也是研究植物传粉生态学不可或缺的关键内容之一[1-3].植物本身的遗传特性是决定植物开花物候特征的一个重要方面,但因为植物在长期的生存进化中也受到周围环境因子的影响,例如海拔高度、温湿度、光照、水分等[4],所以,这些因素也成为决定植物开花物候特征的另一重要方面.量化植物个体或种群的开花物候,主要有开花数量、开花时间和开花持续时间等参数[5,6],而这些特征参数深深地影响着植物的有效传粉者、花粉流动以及交配格局等繁殖特性.通过对不同生境下柔毛淫羊藿(EpimediumpubescensMaxim)的开花物候的研究,探讨微地理环境对同种植物开花物候的影响,摸清其开花物候特征和生殖传粉特性,对于人工栽培提高生殖率具有重要意义.
柔毛淫羊藿是开发潜力巨大、应用前景广阔的中药之一[7],因此,开展生态学的研究工作已经成为野生淫羊藿属植物引种栽培的当务之急.基于以上,本实验作为一个对传粉生态学的研究,为淫羊藿的人工引种驯化、实现规模化人工栽培提供现实依据.
1 材料与方法
1.1 实验地概况
1.1.1 金城山国家森林公园
该实验样地坐落于嘉陵江流域四川段南充市高坪区、蓬安县和广安市岳池县三区县接壤地带,属夏季高温多雨、冬季低温少雨的亚热带季风湿润性气候,年均温17.6℃,最冷月(1月)温度5.14℃,最热月(7月)温度21.7℃[8],发于亚热带常绿阔叶林;雨热同期植物生长期长,无霜期在300d以上.全年降雨量1063mm左右,但分布不均匀,绝大部分降雨集中在5-10月份,暴雨多且集中,其余季节雨水较少,从11月到第二年4月份降雨量只占全年的19%[8].据实地调查,该地植被群落组成主要有:乔木层主要是马尾松(Pinusmassoniana)、川灰木(Symplocosszechumensis)和柏木(Cupressusfunebris);灌木层优势种为杜鹃花(Rhododendronsimsii),草本层植物主要有柔毛淫羊藿、巫山淫羊藿(Epimediumwushanense)、鸢尾(Iristectorom)、复叶耳蕨(Arachniodeschinensis)、细柄草(Capilipediumprviflorum)、苔草(Carexsp.)、吉祥草(Reimeckeacarnea)、卷柏(Selaginellapulvinata)等.
1.1.2 唐家河国家自然保护区
该研究样地设在广元市青川县唐家河国家级自然保护区(104°24′E-104°53′E,32°32′N-32°41′N),位于四川盆地西北缘岷山东端摩天岭南麓,地处横断山脉,北端向青藏高原过渡的高山峡谷地带,东西长大致24.5km,南北宽约18.5km,总面积在400km2左右.总的地势是西北高,东南低,海拔最高处为西北角大草坪,达3 864m,最低海拔1 430m,相对高差2 400m[9].地处亚热带季风性气候,年平均气温在12℃左右,7月平均气温19.7℃,1月平均气温-1.2℃,雨量充沛,年平均降雨量1 100mm.
唐家河自然保护区的植被可细划分为9个基本植被类型[10],经过野外实地考察得出,本区域内柔毛淫羊藿在1 450m-1 990m的海拔梯度范围内均有分布.同时在该海拔梯度上又可更加细分为常绿阔叶林、常绿落叶阔叶混交林和次生落叶阔叶林三个基本植被类型.群落Ⅰ:以油樟(Cinnamomumlongepaniculatum)为优势种的常绿阔叶林,林下郁闭度72%左右,乔木层以油樟和卵叶钓樟(Linderalimprichtii)为主,灌木层主要为刺悬钩子(Rubuspungens)和川莓(R.setchuenensis),柔毛淫羊藿为草本层优势种,伴生种丝叶苔草和禾叶土麦冬等.群落Ⅱ:以细叶青冈(Cyclobalanopsisglaucavar.gracilis)为优势种的常绿落叶阔叶混交林,林下郁闭度为83%左右,乔木层主要以细叶青冈为主,灌木层主要为川莓、野花椒(Zanthoxylumsimulans)和猫儿刺(Ilexpernyi),草本层以柔毛淫羊藿和扶桑金星蕨(Parathelypterisnipponica)共生.群落Ⅲ:以红桦为主要优势种的次生落叶阔叶林.群落乔木盖度度为58%,主要的优势种有红桦,灌木层以糙花箭竹(Fargesiascabrida)为主,草本层优势种为柔毛淫羊藿,伴生川滇苔草(Carexschneideri)和大叶冷水花(Pileamartini)等.就3个群落相比较而言,群落I的土壤含水量是三个群落里较高的,群落Ⅲ的相对光照较大、相对湿度较大、土壤腐殖质较厚.
1.2 材 料
柔毛淫羊藿(EpimediumpubescensMaxim)是多年生草本性植物,属小檗科(Berberdiaceae)淫羊藿属(Epimedium)[11].植株高在20-70cm左右,根状茎粗且短,被褐色鳞片;叶为一回三出复叶,基生或茎生,茎生叶2枚对生,小叶3枚;圆锥花序具30-100余朵花,长10-20cm,通常序轴及花梗被腺毛,有时无总梗,花直径约1cm;萼片2轮;花瓣远较内萼片短,长约2mm,囊状,淡黄色;雄蕊长约4mm,花药长约2mm;雌蕊长约4mm,花柱长约2mm;果实为蒴果长圆形,宿存花柱长喙状,花期4-5月,果期5-7月.
1.3 方 法
1.3.1 种群花期和开花振幅
在前文所述不同种群内,分别人为标记长势良好,生长期大致相同的植株个体30株,并在样株上随机标记一个花序,每天记录植株上开花的数量.观察和统计的开花参数主要有始花期、盛花期、持续时间和末花日期[12],个体的始花日期为第一朵花在花序和个体上开花的日期,种群水平以25%的个体开花时视为始花期,花序和个体开花数大于或等于50%时为盛花期,50%的个体达到开花高峰时视为种群开花高峰日期,95%的植株开花结束时视为末花期[13],花朵全部开放或枯萎时为终花期.
根据Herrera[14]的统计方法计算以下物候参数:①相对开花强度(Relative flowering intensity):植株于开花高峰日期的日产花数量与种群中植株于开花高峰日期日产单株最大花数的比值;②平均开花振幅(Meanfloweringamphtude):单位时间开花数,利用每天开花数量来计算出植株的平均开花振幅(Meanfloweringamplitude),就是单位时间的开花数(花数·株-1·d-1).用同步指数反映开花同步性高低,其方法参照文献如Mcintosh[15]和肖宜安等[16].其计算式为:
式中Si表示开花同步指数,变异范围为0-1,其中“0”表示种群个体花期无重叠性,“i”表示种群个体花期完全重叠.n表示种群中个体总数,fi表示个体i开花的持续时间(d),еj表示种群内个体i和j花期重叠的时间(d).
1.3.2 数据分析
数据均采用excel 2003整理,SPSS19.0统计分析软件进行one-way ANOVA,相关性分析进行数据统计分析.
2 结果与分析
2.1 种群花期和开花振幅
2.1.1 金城山柔毛淫羊藿与唐家河柔毛淫羊藿种群花期和开花振幅
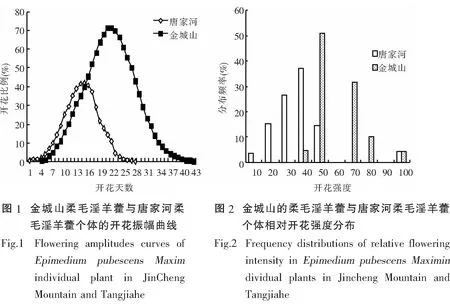
比较金城山柔毛淫羊藿与唐家河柔毛淫羊藿的开花物候进程(开花振幅曲线),二者均呈单峰曲线,表现为开始缓慢上升达到高峰后再逐渐下降.但存在一定差异,金成山柔毛淫羊藿在第23天达到峰值,开花比例能达到的较高的比例在60%-70%,而唐家河柔毛淫羊藿在第16天就能达到峰值,但开花比例仅能达到40%-45%(图1).比较开花强度,二者均表现为集中开花的模式,金城山柔毛淫羊藿开花强度约50%,唐家河柔毛淫羊藿开花强度约40%,因此金城山柔毛淫羊藿表现出的集中开花时间的特性相较唐家河柔毛淫羊藿更为突出(图2).金成山柔毛淫羊藿的个体开花的始花日期、开花高峰日期、花期持续时间、终花日期、相对开花强度和花期同步指数的变化规律参见图3,唐家河柔毛淫羊藿参见图4.
2.1.2 不同种群花期和开花振幅
柔毛淫羊藿开花周期为一年一次,因各地方的生境条件不一样,其始花期、盛花期、终花期也不完全一样.金城山柔毛淫羊藿的花期为3月上旬到4月上旬,种群、个体、花序和单花的花期分别历时30-41d、22-34d、9-18d和3-4d.唐家河柔毛淫羊藿的种群花期、个体、花序和单花的花期分别历时24-25d、18-25d、7-13d、3-4d.以上数据表明,相较而言,金城山柔毛淫羊藿始花期早,终花期也早;唐家河柔毛淫羊藿始花期和终花期都较晚.
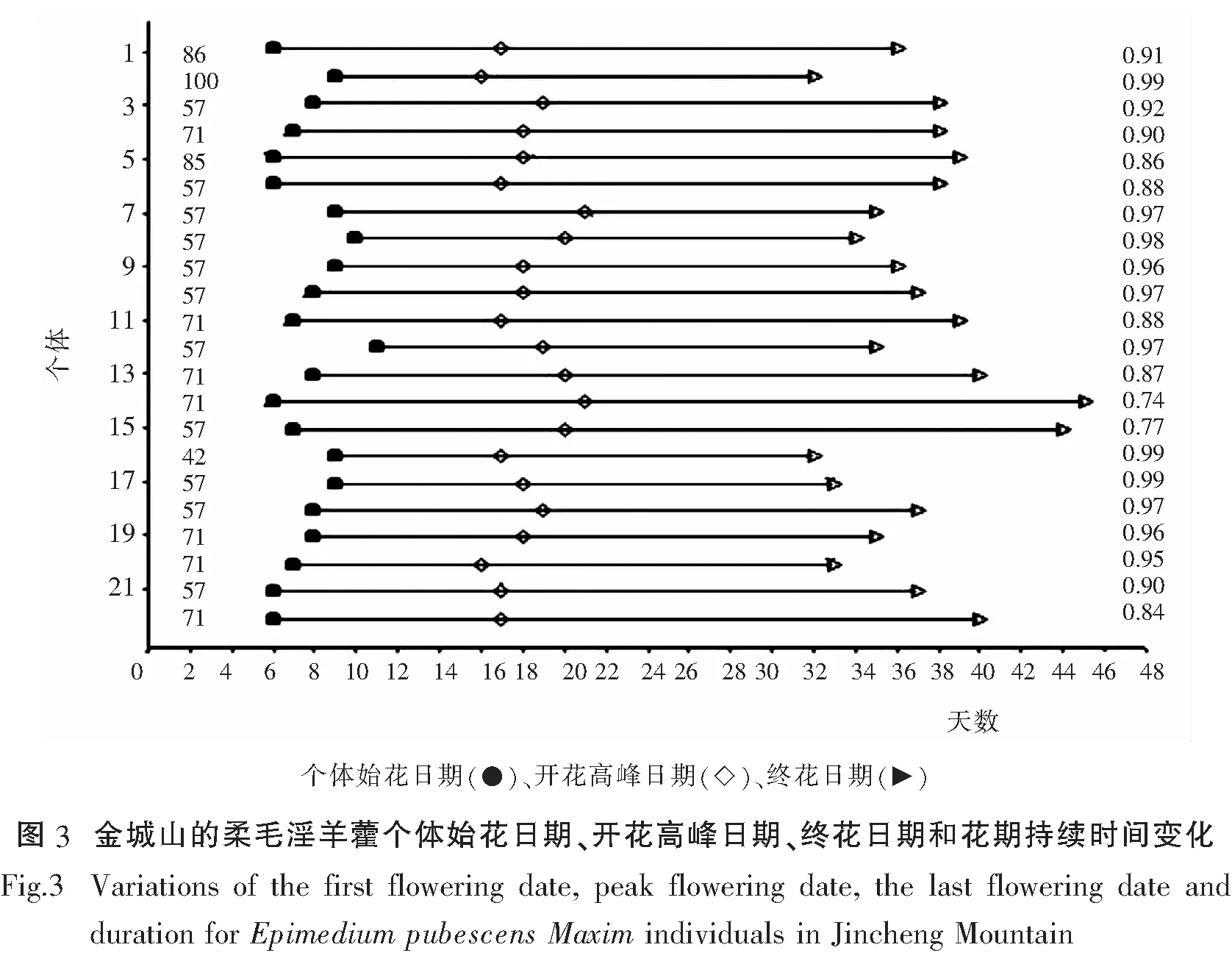
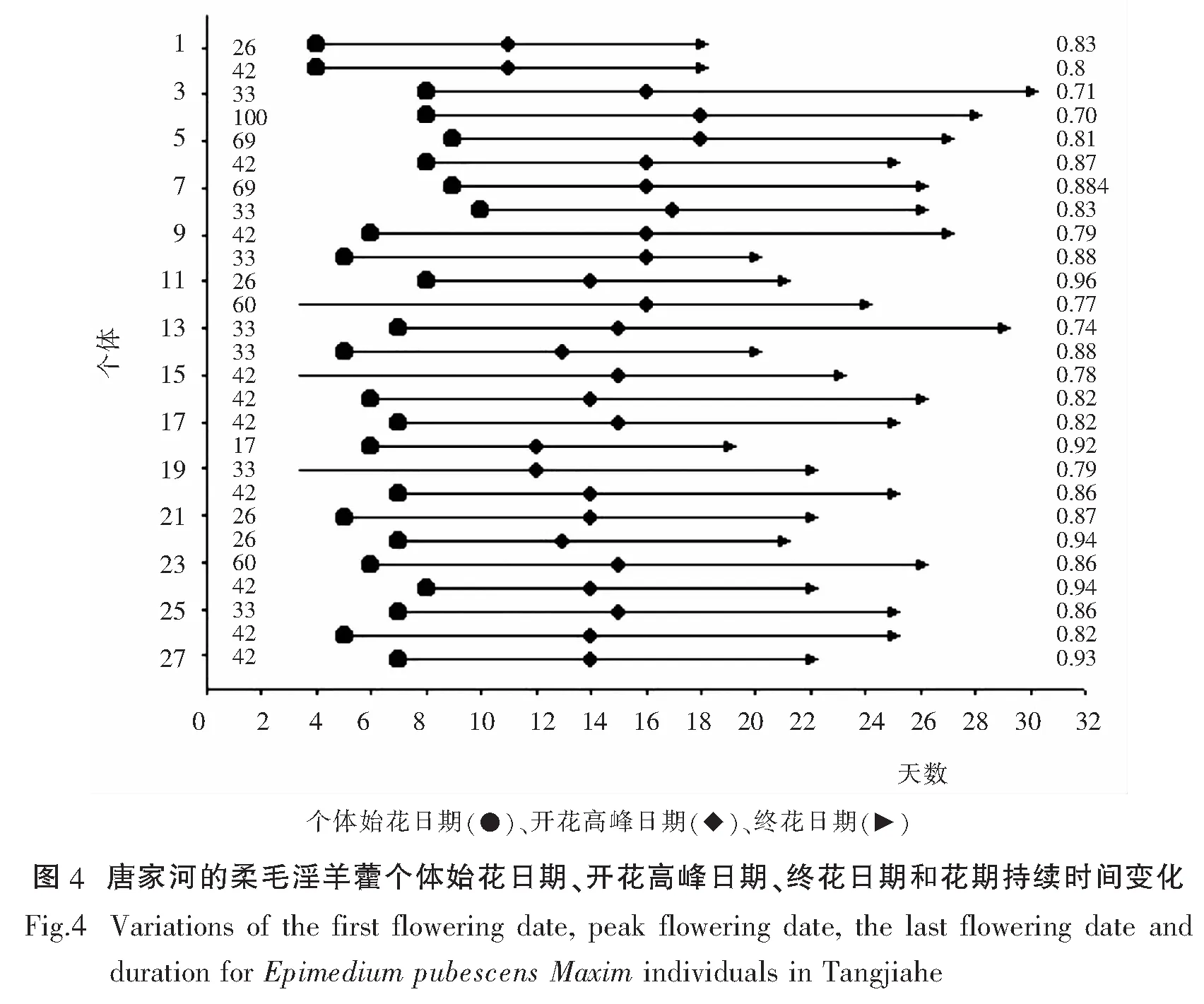
表1 金城山、唐家河两地柔毛淫羊藿花期比较

项目金城山唐家河始花期3-33-29盛花期3-154-9末花期4-24-21持续时间(单位d)3024
2.2 单花开花动态
2.2.1 金城山柔毛淫羊藿的单花开花动态
金城山柔毛淫羊藿单花开花动态,第一天是一个小小的花蕾,花蕾慢慢长大,直至基本长到一定的花长后,外萼片开始脱离,花瓣开始慢慢展开,直至完全开放.发育成熟的花朵,雄蕊和雌蕊开始行使自己的职责—授粉,授粉成功后,雄蕊便渐渐失去水分萎焉,同时伴随着花距的脱落而脱落,当花距和雄蕊都脱落后,花瓣也开始慢慢脱离花朵,直至剩下果实.
2.2.2 唐家河柔毛淫羊藿的单花开花动态
唐家河柔毛淫羊藿单花开花动态,开始同样也是一个小小的花蕾,花蕾慢慢长大,直至基本长到一定的花长后,外萼片开始脱离,花瓣开始慢慢展开,直到完全开放.发育成熟的花朵,雌雄蕊开始行使自己的职责—授粉,授粉成功后,雄蕊便渐渐失去水分萎焉,而与金城山柔毛淫羊藿不同的是,其花距的脱落要早于雄蕊的脱落,当雄蕊脱落后,花瓣开始慢慢脱离花朵,直至剩下果实.
2.3 物候指数与坐果率的相关性分析
2.3.1 金城山柔毛淫羊藿物候指数与坐果率的相关性分析
金城山中的坐果数与始花期成显著负相关,与花期和开花数呈正相关关系0.505和0.628达到极显著水平,花期与始花期呈显著负相关,花期与开花数成显著正相关0.606(P<0.01).

表2 金城山柔毛淫羊藿个体水平的始花日期,花期、开花数及坐果果数的相关分析
注:**.在0.01水平(双侧)上显著相关;*.在0.05水平(双侧)上显著相关.
2.3.2 唐家河柔毛淫羊藿物候指数与坐果率的相关性分析
唐家河的坐果数与花期和开花数呈正相关关系达0.512和0.742,达到极显著水平(P<0.01),花期与开花数呈显著正相关关系达0.610(P<0.01).

表3 唐家河柔毛淫羊藿个体水平的始花日期,花期、开花数及坐果数的相关分析
注:**.在0.01水平(双侧)上显著相关;*.在0.05水平(双侧)上显著相关.
4 讨 论
通过对不同地区(海拔不同,经纬度相异)的柔毛淫羊藿其开花物候进行研究,发现南充金城山地区的柔毛淫羊藿始花期最早,终花期也最早;唐家河地区的始花期和终花期都较晚.表明虽然个体的开花物候一般由植物的遗传因子决定[17],但柔毛淫羊藿开花物候的差异,可能是由不同生境的海拔、光照、水分等生态因子不同而引起的.
一般地认为,海拔较低、纬度较低、气温相对较高的地区,如南充金城山,其柔毛淫羊藿的始花日期和终花日期也较早;而海拔较高、均温较低的唐家河地区,始花日期和终花日期都较晚.这与Blionis等对风铃草属(Campanula)9个物种的研究发现[18],Olsson等对多年生草本植物Lythrumsalicaria不同纬度的群体研究发现[19]结果相似.
植物的开花物候特征与所处的生态环境在一定程度上影响着植物的生殖.这种影响包括物种之间,种群之间以及个体水平之间.如植株个体太小,没有足够的资源发育成成熟果实;花期不同步,雄花资源匮乏;开花过早或太迟,以及开花期间天气不稳定等因素,缺乏传粉者的有效访问等,都会对生殖有很大影响.所以,植物个体的生殖成功与否,植株间生长,种群动态等各方面都受到植株个体的开花物候的影响[20].如金城山中的坐果数与始花期成显著负相关,与花期和开花数呈正相关关系0.505和0.628达到极显著水平,花期与始花期呈显著负相关,开花数与花期成显著正相关0.606.唐家河的坐果数与花期和开花数呈正相关关系达0.512和0.742,达到极显著水平,花期与开花数呈显著正相关关系0.610.
[1] WILLSON M F.Plant Reproductive Ecology[M].New York:John Wiley & Sons,1983:80-85.
[2] WHITEHEAD D R.Wind Pollination:Some ecological and Evolutionary Perspectives[M].In:Real L.(ed.) Pollination Biology.Academic,New York,1983:97-108.
[3] OLLERTON J,DIAZA.Evidence for Stabilizing Selection Acting on Flowering Time in Arum-maculatum (Araceae):The Influence of Phylogeny on Adaptation[J].Oecologia,1999,119:340-348.
[4] RATHCKE B,LACEY E P.Phenological Patterns of terres-trial plants[J].Annual Review Ecology and Systematic,1985,16:179-214.
[5] AUGSPURGER C K.Phenology,Flowering Synchrony and Fruit Set of Six Neotropical Shrub[J].Biotropica,1983,15:257-267.
[6] PICKERING C M.Variation in Flowering Parameters Within and Among Five Species of Austrilian Alpine Ranunculus[J].Austr J Bot,1995,43:103-112.
[7] MORAN J F,BECANA M,ITURBE ORMAETXE I,et a.l Drought induces Oxidative Stress in Pea Plants[J].Planta,1994,194(3):346-352.
[8] 杨子松,黎云祥,钱宝英.金城山国家森林公园淫羊藿生态位研究[J].林业调查规划,2006,131(1):58-62V.
[9] 赵 刚.柔毛淫羊藿的开花物候与结实格局的研究[D].西华师范大学硕士学位论文,2011:10-20.
[10] 胡锦矗.四川唐家河自然保护区综合科学考察报告[M].四川:四川科技出版社,2005:15-26.
[11] 王悦云,何顺志,徐文芬.中国淫羊藿属小花类群分类系统[J].贵州科学,2009,27(4):43-49.
[12] PICKERING C M.Variation in Flowering Parameters Within and Among Five Species of Austrilian Alpine Ranunculus[J].Austr J Bot,1995,43:103-112.
[13] 高泽梅,权秋梅,肖 娟,等.金城山柔毛淫羊藿的开花物候与生殖特性[J].广西植物,2013,33(1):1-6.
[14] HERRERA J.Flowering and Fruiting Phenology in the Coastal Shrublands of Donana[J].S Spain Veg,1986,68:91-98.
[15] MCINTOSH ME.Flowering Phenology and Roductive Output in two Species of Ferocactus (Cactaceae)[J].Plant Ecol,2002,159(1):1-13.
[16] 肖宜安,何 平,李晓红.濒危植物长柄双花木开花物候与生殖特性[J].生态学报,2004,24(1):14-21.
[17] ZIMMERMAN M,GROSS RS.The relationship Between Flowering Phenology and Seed Set in an Herbaceous Perennial plant,Polemonium Foliosissimum Gray[J].Am Midl Nat,1984,111:185-191.
[18] BLIONIS GJ,HALLEY JM,VOKOU D.Flowering Phenology of Cam panula on Mt Olympos,Greece,[J].Ecography,2001,24(6):696-706.
[19] 马文宝,施 翔,张道远,等.准噶尔无叶豆的开花物候与生殖特征[J].植物生态学,2008,32(4):760-767.
[20] BRONSTEIN JL.The Plant-pollinator landscape.in:Hanssos L,Fahrig L,Merriam G.Mosaic Landscapes and Ecological Processes[M].London:Chapman & Hall Press,1995:256-288.
The flowering phenology of Epimedium pubescens Maxim in different habitats
YANG Ping1,CHEN Li-ming2,QUAN Qiu-mei1,Li Yun-xiang1
(1.China West Normal University,Key laboratory of southwest China wildlife resources conservation, 2.Tangjiahe National Nature Reserve,Nanchong 637009,China)
This paper studied flowering phenological characteristics and the relevance between flowering phenology and seed rate ofEpimediumpubescensMaximin JinCheng Mountain and TangJiaHe by using field observations.The results showed that the flowering span of the population,individual,inflorescence and single flowering are different.the population showed a single peak concentration blossom style,The flowering ratio ofEpimediumpubescensMaximin JinCheng Mountain is approximately 60%-70%,the range of flowering intensity distribution focus on 50%;The TangJiaHe’s flowering ratio is approximately 40%-45%,the range of flowering intensity distribution is 20%-50%.the flowering amplitude curves of the individual level were of single peak and higher synchrony,exhibiting a “mass-flowering” pattern.The correlation analysis among the flowering phenology index and fruit number showed that there was correlation between fruit number and flowering date and correlation to flower number in these two places.In conclusion,the flowering phenology ofEpimediumpubescensMaximwas affected by environment easily.so,it exhibits a “mass-flowering” pattern,which may be regarded as an adaptive strategy to ensure its reproductive success.
EpimediumpubescensMaxim;flowering phenology;environmental factors
1673-5072(2015)01-0017-07
2014-09-16
唐家河国家级自然保护区植物保护项目
杨 萍(1990-),女,四川德阳人,西华师范大学生命科学学院硕士研究生,主要从事生态学研究.
黎云祥(1968-),男,四川眉山人,西华师范大学生命科学学院教授,博士生导师,主要从事植物生态学和药用植物学研究.
Q945.17
A
猜你喜欢
乐山师范学院学报(2023年12期)2024-01-22 09:14:22
湖南生态科学学报(2023年3期)2023-09-19 09:16:22
科普童话·神秘大侦探(2022年12期)2023-01-31 08:52:29
能源研究与利用(2022年6期)2023-01-05 08:58:12
贵州科学(2022年5期)2022-11-25 13:26:38
小哥白尼(野生动物)(2020年5期)2020-09-24 09:24:34
中学生英语·阅读与写作(2020年8期)2020-09-24 03:10:42
兰台世界(2017年4期)2017-03-08 08:13:26
剑南文学(2016年23期)2016-11-21 11:36:15
剑南文学(2016年23期)2016-11-21 11:36:15
