
菹草生物量控制对群落中沉水植物生长及水质的影响
2015-03-12 07:41徐伟伟胡维平邓建才朱金格李钦钦张洪梅
生态环境学报 2015年7期
徐伟伟,胡维平,邓建才,朱金格,李钦钦,张洪梅
1. 中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,江苏 南京 210008;2. 中国科学院大学,北京 100049
菹草生物量控制对群落中沉水植物生长及水质的影响
徐伟伟1, 2,胡维平1*,邓建才1,朱金格1,李钦钦1, 2,张洪梅1
1. 中国科学院南京地理与湖泊研究所湖泊与环境国家重点实验室,江苏 南京 210008;2. 中国科学院大学,北京 100049
摘要:沉生植物对维护和提高湖泊生态系统的生物多样性和稳定性,控制湖泊富营养化具有重要生态价值。对于可在水面形成致密冠层的沉水植物,生物量过大时可能会对水体功能产生不利的影响。实验通过对菹草(Potamogeton crispus)+金鱼藻(Ceratophyllum demersum)群落中菹草在春季暴发时采取不同菹草收割强度(100%、80%、60%、40%、20%和0%的收割)进行生物量控制。每种收割处理均在底泥厚度为15 cm,水深为110 cm独立实验池中(4 m长×2 m宽×2.5 m深)进行。实验研究生物量调控对冬季种菹草与春季种金鱼藻交替及水质的影响,探讨群落中菹草的最佳生物量控制强度,为草型湖泊进行沉水植物生态管理提供依据。结果表明:在40%、20%和0%(未收割)菹草收割强度处理下菹草容易产生断枝并且整株上浮,更高强度的收割减小了植株受到的浮力,促进了群落中菹草和金鱼藻的生长,并延长了菹草的生命周期,推迟了菹草的上浮时间。菹草遮光效应在一定程度上促进了金鱼藻的生长,中等偏高强度的菹草收割使得菹草和金鱼藻均处在较高的生物量水平,维持了群落的稳定性。在菹草生长期对其进行60%~80%收割可维持水体营养盐在较低的水平,并能实现冬季种沉水植物菹草与春季种沉水植物金鱼藻在群落中有效交替,可缓解存留菹草衰亡给水质带来的不利影响,强化沉水植物群落的水质净化功能。
关键词:生物量调控;收割强度;群落;金鱼藻;盖度
引用格式:徐伟伟,胡维平,邓建才,朱金格,李钦钦,张洪梅. 菹草生物量控制对群落中沉水植物生长及水质的影响[J]. 生态环境学报, 2015, 24(7): 1222-1227.
XU Weiwei, HU Weiping, DENG Jiancai, ZHU Jinge, LI Qinqin, ZHANG Hongmei. Influence of Harvesting Potamogeton Crispus in A Submerged Plant Community on the Growth of Submerged Aquatic Plants and Their Effects on Water Quality [J]. Ecology and Environmental Sciences, 2015, 24(7): 1222-1227.
水生植物在生长期一方面可通过分泌他感物质直接抑制藻类生长繁殖,另一方面可净化水质、减少沉积物的悬浮(Carpenter et al.,1986;Engelhardt et al.,2001;王立志等,2012)。水生植被的恢复、重建和改造已成为浅水湖泊的生态调控和内源污染负荷控制的重要手段(李文朝,1996;吴振斌等,2003;胡开明等2012)。在一些浅水草型湖泊中,由于湖泊富营养化程度加重,湖泊水生植物群落结构发生变化,部分水域沉水植物生物量过大(邱东茹等,1997),并在水面附近形成致密的冠层,产生较为严重的后果,例如阻碍水体流动、影响湖泊景观休闲功能、衰亡引起水质恶化、加速湖泊沼泽化等,在一定时期不仅没有发挥其吸收营养盐净化水质的作用,反而成为氮、磷的内污染源(李文朝,1996;潘慧云等,2008)。
合理地收割沉水植物既有利于植物的恢复和生长,增加生物多样性,又可提高群落持续地净化水质的稳定性(谷孝鸿等,2002),还能转移出植物中的营养物质,达到湖泊氮、磷收支平衡或减轻湖泊污染负荷目的(Gumbricht,1993;尚士友等,2003),也能避免沉水植物过量生长而带来的负面作用。对生物量主要集中在水体上层或表面的沉水植物(范国兰等,2005),适当收割可以缓解生物量过度集中于水体表层的趋势(左进城等,2009;倪乐意,1999)。不过已有的研究大多局限于单种水生高等植物的生物量过量时的植被改造,对沉水植物镶嵌群落人工调控关注不够(王国祥等,1999)。另外,刚刚恢复的沉水植物群落对外界干扰极为敏感,过度收割可能会导致沉水植物重建工作失败(任文君等,2011),低强度的干扰又可能会导致生物多样性的降低,达不到预期效果。因此,合理的收割强度应该是水生生态系统管理关注的关键问题。
菹草(Potamogeton crispus)是眼子菜科多年生沉水草本植物,分布广泛,是长江中下游湖泊的优势种。该植物具有易成活、生长快和繁殖能力强等特点,常与金鱼藻(Ceratophyllum demersum L.)等水生植物伴生,在冬春季节成为沉水植被的优势种(陈正勇等,2011;李伟和程玉,2000)。菹草通常作为富营养化湖泊生态修复的常用植物,一旦成活,具有暴发性,其较高的盖度和密度、巨大的生物量会阻碍其他物种的引入、定居与扩展。若管理不善,甚至危及已有物种(张圣照等,1999)。当菹草密度较高并且水温升高时,由于自身发棵密集及上部茎叶覆盖,下部根茎透光、通气性较差,易出现烂根现象,继而腐烂断开,再加上植株受到的浮力作用,使整株漂浮。随着天气变化,水体产生对流,漂浮的水草也跟着上下移动,最终不但会失去净化作用,其残体还会在短时间内降解,并恶化水质(戴莽等,1999;王海珍等,2002)。
本文通过在温室内对菹草+金鱼藻群落采取不同菹草收割强度进行生物量控制实验,探讨不同收割强度对群落的生产力、群落特征及冬季种与春季种交替时水质的影响;力图从群落特征变化角度阐明沉水植物群落对不同干扰强度的响应机制,以探讨群落中最佳的生物量控制强度,为进一步开发利用水生高等植物最优配置在富营养化水体中应用技术提供参考,并为草型浅水湖泊进行沉水植物生态管理提供科学依据。
1 材料与方法
1.1实验植物主要特征
菹草较为耐寒,主要生长在静水池沼中或缓流的河水中,早春生长较快,能够在水面大面积弯曲生长,形成遮阳,影响底层水体环境。菹草生长的适温范围为10~20 ℃,在冬季生长缓慢,在水深0.5~2.5 m生长良好,夏季温度升高植物体死亡,石芽掉落湖底并在秋季萌发。
金鱼藻为金鱼藻科金鱼藻属,花期6~7月,果期8~10月,生长于池塘、湖泊和沟渠的静水中。当秋季光照渐短,气温下降时,侧枝顶端停止生长,叶密集成叶簇,角质增厚,成为一种特殊的营养繁殖体,休眠顶芽,顶芽很易脱落,沉于泥中休眠越冬,在第二年春天萌发为新株。另外,在生长期中,折断的植株可随时发育成新株。金鱼藻种子具坚硬的外壳,有较长的休眠期,在春季萌发。
1.2实验系统环境
实验在中国科学院南京地理与湖泊所野外观测站东山站温室内6个独立实验池中进行,实验池长×宽×深均为4 m×2 m×2.5 m,实验池中底泥厚度为15 cm,水深为110 cm,透明度为110 cm。实验池内金鱼藻种群已经延续4年,在夏季金鱼藻盖度均达到100%。于2011年11月在各实验池内均匀播撒菹草石芽,播种密度为50颗·m-2,此时金鱼藻已经衰亡,依靠顶芽越冬。2012年4月12日实验开始时3日内日均水温为(22.5±0.4) ℃,各水池内水生植物菹草(P. crispus)和金鱼藻(C. demersum)分布均匀,菹草已经生长至水面,其水面盖度均达到100%,平均生物量为(1023.9±90.8) g·m-2,而金鱼藻生长在水体底层,平均生物量为(83.6±28.5) g·m-2。各实验池水体均来自东太湖,试验开始时总氮(TN)为(1.37±0.20) mg·L-1,总磷(TP)为(0.017±0.004) mg·L-1。
1.3实验方案
实验自2012年4月12日开始,共进行69 d。于2012年4月12日对菹草+金鱼藻群落中菹草在春季暴发时采取不同收割强度进行菹草生物量控制实验,菹草收割强度设100%(全部收割)、80%、60%、40%、20%和0%(未收割)的生物量收获,收割时以菹草盖度为度量,间隔均匀地依各强度对菹草整株采用手工绞拔方式进行生物量收获,并清除因收割断浮的植物残体。由于此时的金鱼藻冠层较低,拔除菹草对金鱼藻基本无物理损伤。
1.4分析项目与方法
实验开始后,连续跟踪监测菹草和金鱼藻生长和衰亡过程。于2012年5月5日、5月25日和6 月20日,进行各处理的菹草和金鱼藻的鲜生物量、盖度以及植株上浮现象(植株基部断裂后浮在水面)调查。在植物样品采集前,先在各实验池采集500 mL的水样。各次生物量调查取样在水池中随机选择3个25 cm×25 cm小样方,拨出样方内的所有植株(剔除与样方边界相交的两条边线上的植物),分别称取不同水生植物的鲜重。水样采用标准方法进行TN和TP含量的测定(国家环保总局,2002)。实验池水温使用YSI 6600V2型多参数水质监测仪测定。
1.5数据处理
不同时间段生物量增长率计算公式为:Gn=(Ln–Ln-1)/Ln-1/(Tn-Tn-1),n=1, 2, 3
其中G1、G2和G3分别为2012年4月12日至5月5日、5月5日至5月25日和5月25日至6 月20日植物生物量的增长率,Ln是第n次调查日时植物的生物量,l0是为2012年4月12日收割处理后的生物量,Tn是从实验开始至第n次调查日时实验进行的天数。
数据分析采用SPSS 11.5软件,选择单因素方差分析(One-Way ANOVA)进行差异显著性检验。
2 结果与分析
2.1不同收割强度对两种沉水植物生物量和菹草上浮的影响
2.1.1收割强度对菹草生物量和上浮的影响
实验结果表明,4月中旬菹草经100%收割处理后,菹草失去了再生长的能力(表1)。而在菹草的生长期,适量的收割对菹草的生长是有利的,但是由于水温的快速升高,在2012年5月25日之前平均水温已经超过26 ℃,菹草进入了明显的衰亡期。此后,除全部收割外,高强度收割延缓了菹草的生物量减少的趋势,在6月20日,80%和60%菹草收割处理条件下,其生物量分别是未收割处理条件下的16.8和10.0倍,而20%菹草收割处理与未收割处理相比菹草生物量无显著差异(P=0.475)。2012 年4月12日至2012年5月5日菹草生物量增长率随着收割强度的降低而减小,80%的收割处理菹草增长率达到0.189 d-1;第2阶段差异不显著,除全部收割无菹草存活的处理外,增长率介与-0.019 d-1至-0.013 d-1;第3阶段呈现出随着初始收割强度的增加植物衰亡减慢的趋势(图1)。自5月5日后,菹草在80%收割处理中衰亡相对较慢且较为稳定,而其他处理衰亡速率加快。虽然菹草的衰亡速率主要是受到温度上升的影响,但是在实验初期收割给菹草提供了再生长的空间,菹草产生大量新生分枝导致植株保持生长趋势,延迟了菹草的衰亡时间。
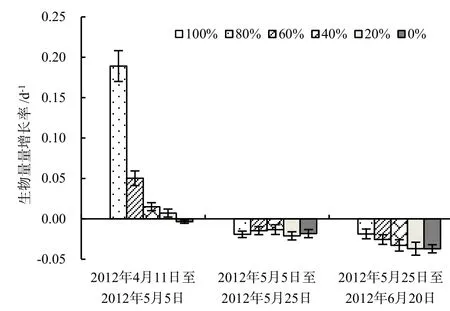
图1 菹草生物量增长率变化(n=3)Fig. 1 Changes of increasing rate of biomass of P. crispus (n=3)
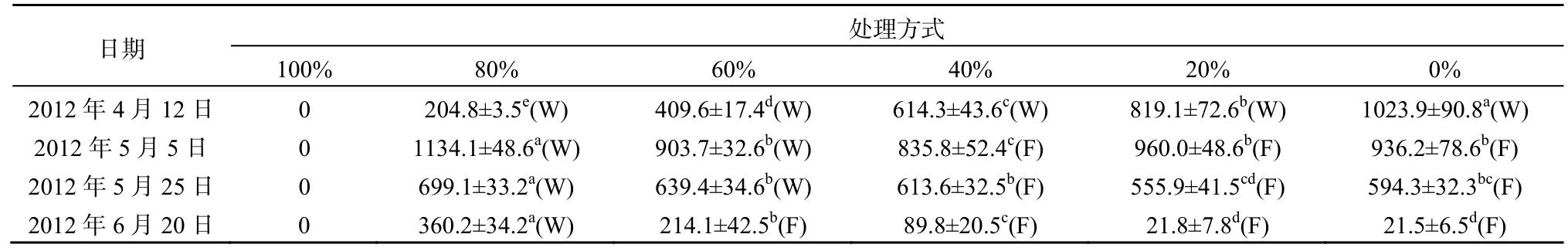
表1 不同收割强度下菹草生物量变化及上浮*Table 1 Biomass and buoyancy of P. crispus under different harvesting intensities g·m-2
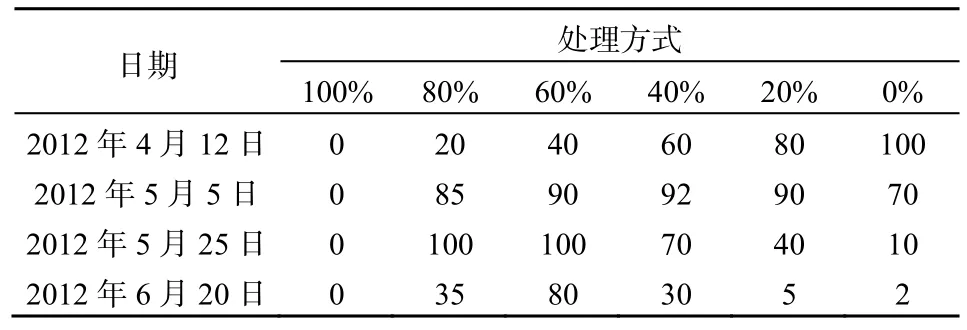
表2 不同收割强度下菹草的盖度Table 2 Coverage of P. crispus under different harvesting intensities %
沉水植物同陆生植物一样,根部固定在基质中,保持自身相对固定的状态。但是不同于陆生植物,沉水植物不能依靠自身的茎组织刚性保持相对趋向水面立地状态。在静止的水体中,沉水植物所受力主要有浮力、重力及植物根系的固着力,其中固着力和重力为向下的作用力,而浮力为向上的作用力,这些力的共同作用维持其在水体直立或植株上部漂浮于水面的状态。根据阿基米德定律,沉水植物受到的浮力即植物排开水的体积与水的比重乘积,植物受到的重力等于植物的重量。菹草成熟无性系的生物量分配模式,从根、茎、叶生物量依次增加,呈现与马来眼子菜、川蔓藻等相反的头重脚轻的结构(沈佳等,2009),所以在一定的条件下容易上浮。收割菹草无性系中的部分植物减小了植株受到的浮力。在5月25日,只有80%和60%菹草收割处理下菹草未发生上浮现象,即便此时菹草的盖度也达到了85%以上,说明收割后新生长的菹草不容易断枝上浮。至6月20日,仅有80%菹草收割处理未发生上浮现象(表1)。菹草生长、衰亡和上浮使得其盖度发生相应的变化(表2),5月5日至5月25日80%和60%菹草收割处理下虽未发生大面积上浮现象,但其生物量的减少表明菹草亦在衰亡。综合菹草的盖度呈增长的趋势,可推断菹草该段时间内菹草仅在冠层表现出增长迹象,种群从面状的分布向丛状分布的方向发展。6月20日,60%菹草收割处理下菹草盖度大于80%菹草收割处理,因其生物量远低于后者,菹草上浮可能是导致盖度较大的原因。综合结果表明较高强度收割延长了菹草的生命周期,推迟了菹草的上浮时间,改变了菹草种群生物量在水体中分布格局。
2.1.2收割强度对金鱼藻生物量的影响
各次调查中金鱼藻生物量最大值均未出现在100%菹草收割的处理中,而是出现在中高强度的收割处理中(表3)。金鱼藻增长率最大值出现在2012 年4月12日至2012年5月5日60%收割处理中,达到0.107 d-1(图2)。水体较高的温度和光照强度可能导致金鱼藻植株的死亡率升高,在水体较浅、透明度较高的条件下,菹草对水体适当遮光可能有利于促进金鱼藻的生长,这种现象在适量的漂浮植物可以促进沉水植物生长的研究中有过类似报道(Janes et al.,1996)。在5月5日(实验第23天),除未收割处理的金鱼藻的生物量仅63.0 g·m-2外,其他处理中金鱼藻的生物量均超过200 g·m-2,说明在菹草+金鱼藻群落中,金鱼藻处于生长初期时即便低强度地收割菹草也会使得金鱼藻生物量在短期内迅速增加。结合表1可以看出在实验后期各处理中当菹草上浮时,金鱼藻的生物量下降,过量的上浮菹草对金鱼藻生长是不利的。在2012年5月5日至2012年5月25日,金鱼藻增长率在40%处理中最高,达到0.034 d-1,而此时段该处理中菹草生物量与20%收割处理相当,但是20%处理中金鱼藻增长率-0.009 d-1,可见在金鱼藻生长初期其生物量的增加对40%的收割处理比20%的收割处理的响应更为敏感。100%与80%菹草收割处理中金鱼藻生物量在整个实验过程中均呈升高趋势,不过后者生物量增长率更大。实验结束时,80%菹草收割处理中金鱼藻的生物量分别是0%和100%的8.8和1.3倍。在未收割处理中,随着时间推移菹草盖度也在不断下降,而金鱼藻的生物量也呈下降趋势,主要原因可能跟金鱼藻所处的生长期有关,在早春对菹草进行刈割可更为有效促进金鱼藻的恢复。
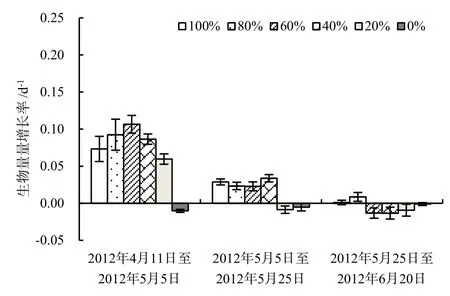
图2 金鱼藻生物量增长率变化(n=3)Fig. 2 Changes of increasing rate of biomass of C. demersum (n=3)
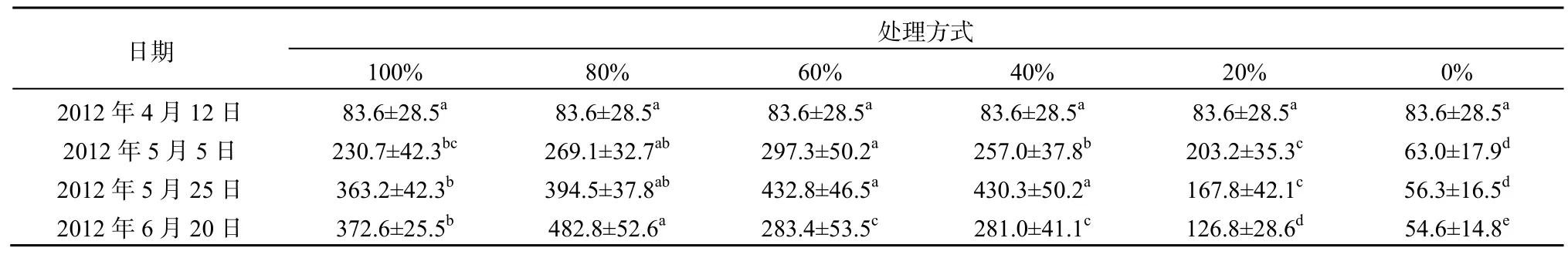
表3 不同收割强度下金鱼藻生物量变化Table 3 Biomass of C. demersum under different harvesting intensities g·m-2
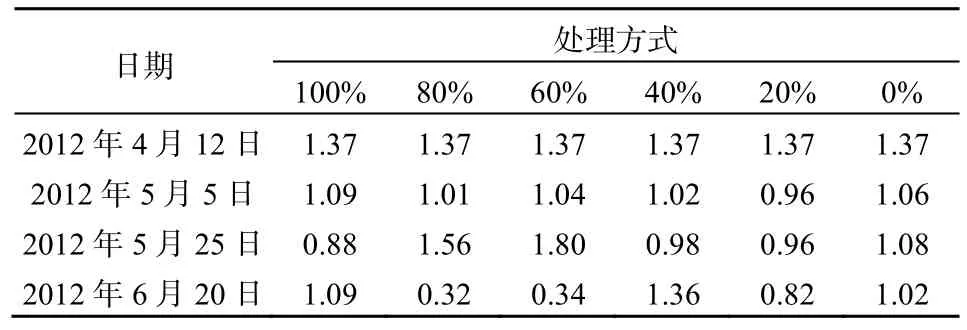
表4 不同收割强度下TN的变化Table 4 TN change under different harvesting intensities mg·L-1
菹草不仅生长速率较快,植株能够伸至水面弯曲漂浮生长,占据较大的水面,而且种群在生长和发育时间上亦早于金鱼藻,使得金鱼藻在种间竞争上处于劣势。群落上层物种遮蔽性去除和竞争力的削弱,给群落下层的物种更多机会接受光照,使之光合作用增强,对群落整体生物量起到补偿作用从而使得群落总的生物量增大,并且保持一定的生物量。在高强度收割条件下,菹草依然能产生石芽(吴晓东等,2012),因此不会影响菹草种群延续。实验结果表明通过对菹草生物量进行中、高强度的控制,可以延长2种沉水植物共存的时间,可最大限度地保证水体整年均有一定量的水生植物。
2.2收割强度对水质的影响
2012年5月25日的水质结果显示各处理间TN接近,说明菹草收割后短时间内对水体TN未产生显著影响(表4)。此后,100%和0%的菹草收割处理中TN基本稳定在1.0 mg·L-1,这2种处理基本上可以代表2种单一物种的水质净化效果,但是这种效果在后期不如沉水植物群落的TN净化效果。在菹草最迟上浮的80%和60%收割处理条件下,TN 在5月25日高于其他处理,此时菹草的盖度又达到了100%,影响了水质的净化效果,但是在6月20日低于其他处理,其原因可能是前期收割促进了金鱼藻的恢复并保有一定的生物量,部分菹草植物腐烂释放的营养盐被金鱼藻利用,后期菹草生物量和盖度稳定在较低水平促进了水质的改善。推广应用时可考虑收割后菹草盖度接近100%时对菹草再进行一次适量的收割处理。80%和60%收割处理条件下后期因菹草与金鱼藻的联合净化作用,使TN维持在较低的水平。40%收割处理下TN在后期达到1.36 mg·L-1的原因应该是上浮的菹草开始大量腐烂,加上因上浮菹草的遮阴作用导致金鱼藻的生物量大幅减少,最终TN达到较高值。菹草收割对TP的影响与对TN的影响基本一致(表5),在5月25日后,TP的大幅增加可能与水温升高引起菹草衰亡加速和底泥释放有关。6月20日TP的最大值出现在0%的菹草收割处理中,而最小值出现在80%处理中。mg·L-1
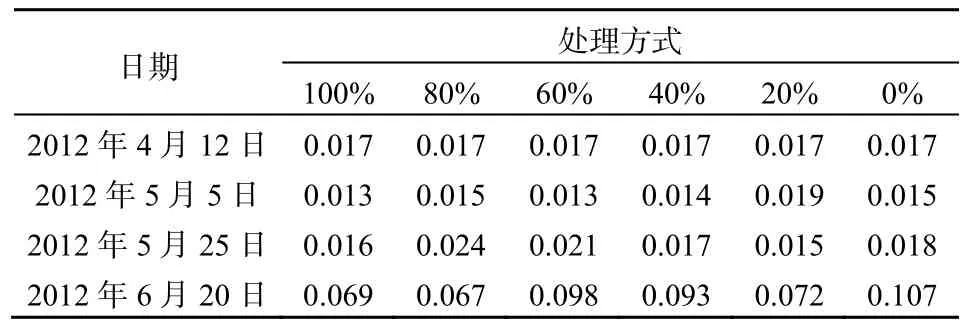
表5 不同收割强度下TP的变化Table 5 TP change different under harvesting intensities
收割对水质的影响结果表明,在生长期高强度的菹草收割可维持水体营养盐在较低的水平。有研究表明随着菹草进入衰亡期,单一菹草种群区营养盐浓度开始升高,而在菹草群落区由于其他沉水植物的作用,对水体营养盐的去除能力显著强于单一菹草种群区(王文林等,2006)。收割后菹草和金鱼藻生物量的同步增长可在春末夏初时有效缓解剩余菹草死亡给水质带来的不利影响,并可强化沉水植物群落的水质净化功能,使水质保持相对稳定。因此,对沉水植物群落在植物生长期进行选择性收割和生态管理十分必要。
3 结论
(1)在40%、20%和0%(未收割)菹草收割强度处理下,菹草容易产生断枝并且整株上浮,更高强度的物理去除减小了植株受到的浮力,促进了群落中菹草和金鱼藻的生长,并延长了菹草的生命周期,推迟了菹草的上浮时间。
(2)金鱼藻生物量最大值均未出现在100%收割菹草的处理中,菹草对水体适当遮光可有促进金鱼藻的生长,高强度的菹草收割均促进了菹草和金鱼藻的生长,在早春通过刈割菹草降低其盖度可更为有效促进金鱼藻的恢复,维持群落的稳定性。
(3)在生长期对菹草进行60%~80%收割处理可维持水体营养盐在较低的水平,并能实现冬季种沉水植物菹草与春季种沉水植物金鱼藻在群落中有效交替,可缓解剩余菹草衰亡给水质带来的不利影响,强化沉水植物群落的水质净化功能。
参考文献:
CARPENTER S R, LODGE D M. 1986. Effects of submersed macrophytes on ecosystem processes [J]. Aquatic Botany, 26: 341-370
ENGELHARDT K A, RITCHIE M E. 2001. Effects of macrophyte species richness on wetland ecosystem functioning and services [J]. Nature, 411(6838): 687-689.
GUMBRICHT T. 1993. Nutrient removal processes in freshwater submersed macrophyte systems [J]. Ecological Engineering, 2(1): 21-30.
JANES R A, EATON J W, HARDWICK K. 1996. The effects of floating mats of Azolla filiculoides Lam. and Lemna minuta Kunth on the growth of submerged macrophytes [J]. Hydrobiologia, 340: 1-3.
陈正勇, 王国祥, 吴晓东, 等. 2011. 不同水深条件下菹草(Potamogeton crispus)的适应对策[J]. 湖泊科学, 23(6): 942-948.
戴莽, 倪乐意, 谢平, 等. 1999. 利用大型围隔研究沉水植被对水体富营养化的影响[J]. 水生生物学报, 23(2): 97-101.
范国兰, 李伟. 2005. 穗花狐尾藻(Myriophyllum spicatum)在不同程度富营养化水体中的营养积累特点及营养分配对策[J]. 武汉植物学研究, 23(3): 267-271.
谷孝鸿, 陈开宁, 胡耀辉. 2002. 东太湖伊乐藻的营养繁殖及对渔业污水的净化[J]. 上海环境科学, 21(2): 43-45, 65.
国家环保总局《水和废水监测分析方法》编委会. 2002.水和废水监测分析方法(第4版)[M]. 北京: 中国环境科学出版社: 216-219.
胡开明, 逄勇, 余辉, 等. 2012. 太湖水环境功能区调整方案[J]. 河海大学学报(自然科学版), 40(5): 503-508.
李伟, 程玉. 2000. 洪湖主要沉水植物群落的定量分析Ⅲ. 金鱼藻+菹草+穗花狐尾藻群落[J]. 水生生物学报, 24(1): 30-35.
李文朝, 陈开宁, 吴庆龙, 等. 2001. 东太湖水生植物生物质腐烂分解实验[J]. 湖泊科学, 13(4): 331-336.
李文朝. 1996. 浅型富营养湖泊的生态恢复——五里湖水生植被重建实验[J]. 湖泊科学, 8(增刊): 1-10.
倪乐意. 1999. 切除顶枝对加拿大伊乐藻生长的影响[J]. 水生生物学报, 23(4): 297-303.
潘慧云, 徐小花, 高士祥. 2008. 沉水植物衰亡过程中营养盐的释放过程及规律[J]. 环境科学研究, 21(1): 64-68.
邱东茹, 吴振斌. 1997. 富营养化浅水湖泊沉水水生植被的衰退有恢复[J]. 湖泊科学, 9(1): 82-88.
任文君, 田在锋, 宁国辉, 等. 2011. 4种沉水植物对白洋淀富营养化水体净化效果的研究[J]. 生态环境学报, 20(2): 345-352.
尚士友, 杜健民, 李旭英, 等. 2003. 草型富营养化湖泊生态恢复工程技术的研究——内蒙古乌梁素海生态恢复工程实验研究[J]. 生态学杂志, 22(6): 57-62.
沈佳, 石福臣, 许文. 2009. 菹草无性系季节生长速度和生物量分配特征[J]. 水生生物学报, 33(2): 246-253.
王国祥, 濮培民, 张圣照, 等. 1999. 冬季水生高等植物对富营养化湖水的净化作用[J]. 中国环境科学, 9(2): 106-109.
王海珍, 刘永定, 肖邦定, 等. 2002. 围隔中鲢和菹草控藻效果及其生态学意义[J]. 水生生物学报, 28(2): 141-146.
王立志, 王国祥. 2012. 苦草密度对扰动引起各形态磷释放的影响[J].生态环境学报, 21(6): 1096-1101.
王文林, 王国祥, 李强, 等. 2006. 菹草伊乐藻群落对富营养化水体水质的净化效果[J]. 南京师大学报(自然科学版), 29(4): 111-116.
吴晓东, 王国祥, 魏宏农, 等. 2012. 不同收割强度对菹草(Potamogeton crispus)生长和光合荧光特性的影响[J]. 湖泊科学, 24(5): 732-738.
吴振斌, 丘东茹, 贺锋, 等. 2003. 沉水植物重建对富营养水体氮磷营养水平的影响[J]. 应用生态学报, 14(8): 1351-1353.
张圣照, 王国祥, 濮培民, 等. 1999. 东太湖水生植被及其沼泽化趋势[J].植物资源与环境, 8(2): 1-6.
左进城, 苗凤萍, 王爱云, 等. 2009. 收割对穗花狐尾藻生长的影响[J].生态学杂志, 28(4): 643-647.
Influence of Harvesting Potamogeton Crispus in A Submerged Plant Community on the Growth of Submerged Aquatic Plants and Their Effects on Water Quality
XU Weiwei1, 2, HU Weiping1*, DENG Jiancai1, ZHU Jinge1, LI Qinqin1, 2, ZHANG Hongmei1
1. State Key Laboratory of Lake Science and Environment, Nanjing Institute of Geography and Limnology, Chinese Academy of Sciences, Nanjing 210008, China; 2. University School of the Chinese Academy of Sciences, Beijing 100049, China
Abstract:Submerged macrophytes are extremely important because they serve multiple ecological functions. The submerged macrophytes, called canopy formers, are notorious for forming dense mats of “topped-out” vegetation that can displace native plants, impede navigation, and reduce the aesthetic enjoyment of affected aquatic systems because the biomass disproportionately concentrates near the water surface. An experiment of biomass regulation was conducted in the rapid growth stage of submerged macrophytes under different harvest intensities (100%, 80%, 60%, 40%, 20% and 0% harvesting Potamogeton crispus ) in P. crispus-Ceratophyllum demersum community. Each harvest intensity was manipulated in a separate concrete water pond (4 m long, 2 m wide and 2.5 m deep) independently, with sediment thickness of 15 cm, and water depth of 110 cm. The effects of biomass regulation on the alternation of winter-spring species, and the water quality by harvest was studied. The objectives of this study were to discuss the best harvest intensity in the community, and provide a scientific basis for ecosystem management of submerged plants in the submerged plants dominated lakes. The results showed that P. crispus were floated on the water under harvest intensity of 40%, 20% and 0% (no harvesting), and higher harvest intensity decreased the buoyancy of P. crispus, extended the life cycle of P. crispus and postponed the floating date. The proper shading on water bodies by P. crispus promoted the growth of C. demersum, and high harvest intensity promoted the growth of both P. crispus and C. demersum to maintain the stability of the community. The nutrient of water can be maintained at a low level under harvest intensity of 60% to 80% in the growth phase of P. crispus which can achieve efficacious alternation of winter-species P. crispus and spring-species C. demersum in the community. Harvesting alleviated adverse effects of the P. crispus decline to water quality, and consolidated the water purification function of submerged plant community.
Key words:biomass regulation; harvest intensity; communities; Ceratophyllum demersum; coverage
收稿日期:2014-12-23
*通讯作者:胡维平(1965年生),男,研究员,主要从事湖泊生态工程、生态模型研究。E-mail: wphu@niglas.ac.cn
作者简介:徐伟伟(1986年生),男,博士研究生,主要从事湖泊生态研究。E-mail: 409000498@qq.com
基金项目:国家自然科学基金项目(NSFC41230853);国家水体污染控制与治理科技重大专项项目(2014ZX07101-011);中国科学院重点部署项目(KZZD-EW-10)
中图分类号:X173
文献标志码:A
文章编号:1674-5906(2015)07-1222-06
DOI:10.16258/j.cnki.1674-5906.2015.07.022
猜你喜欢
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
黑龙江工程学院学报(2020年5期)2020-10-21
水土保持研究(2019年6期)2019-10-19
生态毒理学报(2018年6期)2018-03-16
黑龙江水产(2017年2期)2017-03-02
草业科学(2015年9期)2015-12-11
创新作文(1-2年级)(2015年10期)2015-10-21
海南热带海洋学院学报(2015年5期)2015-03-14
江苏农业科学(2014年3期)2014-07-16
