
不同适应条件对细鳞鲑幼鱼游泳能力的影响
2015-03-11 06:47徐革锋尹家胜牟振波
生态学报 2015年6期
徐革锋,尹家胜,韩 英,马 波,牟振波,*
1 中国水产科学研究院黑龙江水产研究所, 哈尔滨 150070 2 东北农业大学动物科技学院, 哈尔滨 150030
不同适应条件对细鳞鲑幼鱼游泳能力的影响
徐革锋1,2,尹家胜1,韩 英2,马 波1,牟振波1,*
1 中国水产科学研究院黑龙江水产研究所, 哈尔滨 150070 2 东北农业大学动物科技学院, 哈尔滨 150030
为了研究适应条件对鲑科鱼类幼鱼续航游泳能力的影响,在(16.0±0.2)℃和8 mg/L溶解氧条件下,以续航时间作为评定游泳能力的关键指标,通过适应时间、适应流速、加速时间以及测试流速对细鳞鲑幼鱼的续航游泳时间的影响进行了研究, 试验采用4因素8水平的均匀设计方案。结果表明,最大适应流速应控制在1—1.5 BL/s为宜,相应的最佳适应时间应为1 h,在合理条件下适应流速和适应时间对续航游泳能力的影响可忽略不计;加速时间只在大流速条件下(>0.5 m/s)对续航时间有极显著影响,其他情况的影响可以忽略不计;尽管长时间适应环境更有利于鱼类身体机能的充分调整,但适应时间仍建议控制在1—2 h以内,且受试鱼类要经过试验前的适应能力筛选;测试流速建议控制在3—4 BL/s以内,以避免超过鱼类的最大游泳耐受范围。测试流速对细鳞鲑幼鱼的续航时间有极为显著的影响,其续航时间随测试流速的增加呈幂函数规律衰减。
细鳞鲑;适应时间;适应流速;加速时间;续航时间
游泳能力是关乎鱼类生存的至关重要因素,且大多鱼类必须通过不同游泳形式进行逃避敌害或灾害环境、猎食、求偶、迁徙等活动[1-3]。大多学者通常将游泳速度和游泳时间作为评价鱼类游泳能力的关键指标[4-5],并利用游泳时间将游泳能力区分为持续游泳能力和短暂游泳能力,而持续游泳又区分为恒速游泳和变速游泳[6];因此,相应延伸出了鱼类游泳速度的划分标准,持续游泳能力主要包括最大续航游泳速度[7-8]、最优巡航游泳速度[9-12]、最大游泳速度[13]、临界游泳速度[14-16]、短暂游泳能力主要为冲刺游泳速度[17-19]等。一般认为,最大续航速度与鱼类在自然水域的真实游泳能力最为贴近,也是鱼类索饵、迁徙或洄游等的主要运动形式。而续航属于非疲劳状态,且最大续航速度一般无法直接测定。但大多学者认为,通过多组次续航时间的测定,并构建数学模型可获得任意给定续航时间下的最大续航速度[8, 20-21]。而鱼类的最大续航速度和最大续航时间的测定结果受适应条件的影响极大,如果不区别对待鱼的种类特异性,将导致大多研究结果差异较大而失去可比性。因此,设计合理的适应条件不但对鱼类游泳能力的准确测定起到至关重要的作用,而且对一些拦水工程的鱼道设计具有指导意义,即在进行试验之初要设计合理的适应条件,对于鱼道的激流区设计要充分考虑鱼的适应能力,避免鱼类适应不足或是疲劳过度浪费体力,而导致生理应激或是无法克服水流速度及流态造成的水流障碍,从而试验失败或鱼道设计产生重大失误[22]。
水槽实验法是进行鱼类游泳能力及游泳行为等基础研究所采用的最常用手段[7-8, 16, 21]。这种试验方法可对众多复杂的环境因子进行有目的的调控,将最主要刺激源进行强化,易于直接观测及定量比较,试验重复性较高。大多学者都非常重视测试前的准备与适应过程,但对适应条件(水温、光照、溶解氧、初始适应流速、适应时间、加速适应等)的选择与其对测试结果的影响通常未做过深入探讨,无法确定适应条件与测试结果的相关程度,导致不同研究之间的比较缺乏足够的说服力。本研究选择了我国珍稀、名贵的细鳞鲑(Brachymystaxlenok)幼鱼进行测定试验,目前该鱼是我国“十二五”期间主推的名特优品种之一,近年来在有关细鳞鲑的健康养殖、疾病和生物学等方面的报道较多[23-25],但未见在行为生态学方面的研究报道。本文借鉴桂福坤等[21]方法,在恒定的环境因子条件下,通过调整适应流速、适应时间、加速时间这3种适应条件来研究对幼鱼阶段细鳞鲑的最大续航时间和最大续航速度的影响,旨在为今后开展鲑科鱼类的行为学、运动能量学和行为生态学等方面的试验设计,尤其在前期适应条件方面提供有价值的资料,并对拦水工程的鱼道设计提供鲑科鱼类游泳能力的基础参考数据。
1 材料与方法
1.1 试验对象
细鳞鲑(Brachymystaxlenok)来自于中国水产科学研究院黑龙江水产研究所丹东养殖基地,为5月龄幼鱼,共60尾,体长为(18.52±0.35)cm, 体重为(64.50±3.5)g。试验前, 受试鱼被暂养于控温循环水水族箱内(1.5 m × 0.5 m×0.6 m),水温为(16.0±0.2)℃,溶解氧大于8.0 mg/L,光照为不作特殊控制的自然光,暂养时间10 d。当试验鱼适应了暂养环境,继而进行为期10 d的养殖驯化,选用鲑鳟鱼专用饲料(北京,思诺)每日按鱼总体重的2%进行投喂,每天8:00和14:00各投喂1次。水族箱内配备了污水过滤、紫外杀菌、精确控温、除氨等辅助系统,污水过滤系统每1—3 d清洗1次,且每天交换20%的水体。
1.2 试验设计
本试验设计主要借鉴桂福坤等[21]方法,选择续航时间作为细鳞鲑幼鱼的游泳能力的评价指标。试验共设定3个适应因子,分别为鱼在水槽中的流速适应、时间适应以及由适应流速升至测试流速的加速适应,加上最大续航流速(测试流速)共4个因子,并且忽略各因子间的交互影响,以降低试验强度。细鳞鲑幼鱼的续航时间测定采用4因子8水平的均匀设计试验方案[26](表1),共需试验对象48尾,分为8组,每组2尾鱼,试验共设3个重复组,每组测试后不再进行重复试验。
试验对象的测试流速范围根据预试验获得的相关数据进行设定,测试流速范围为0.25—0.60 m/s,流速水平为均匀递增;适应流速以大致1.0 BL/s(0.19 m/s)作为最大值,适应流速水平为均匀递减。
适应时间范围根据预试验情况设定,范围为5—75 min,适应时间的水平增量为10 min;加速时间设定为8个水平,为1—8 min,加速时间水平增量为1 min。
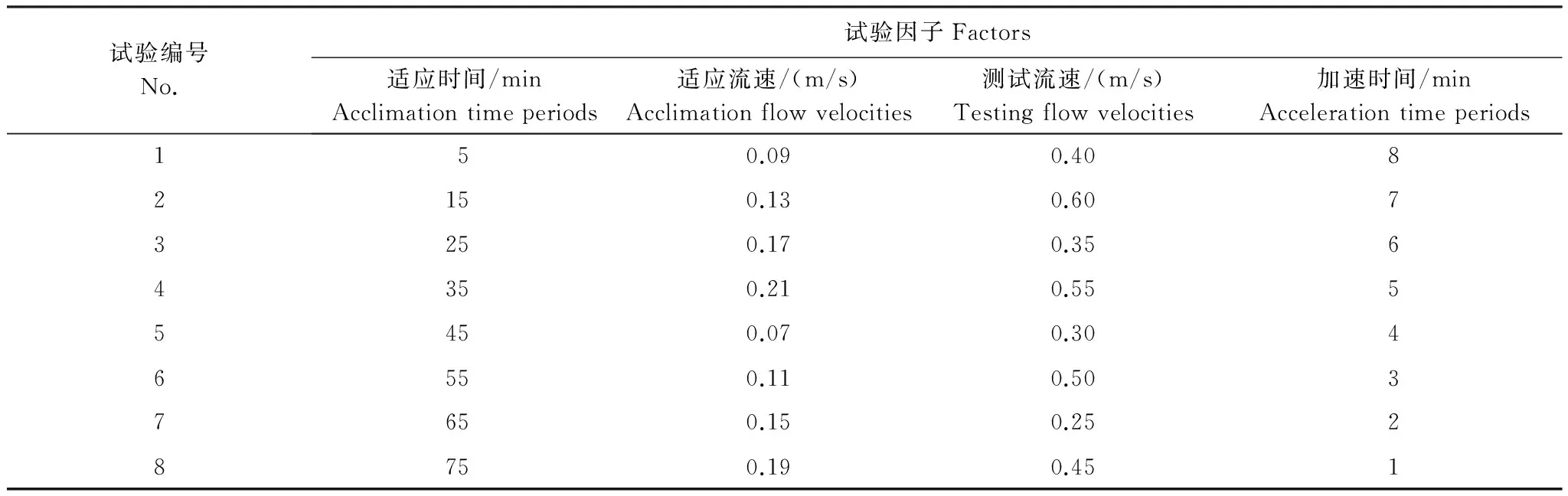
表1 细鳞鲑续航时间试验均匀设计方案Table 1 Uniform design for lenok sustained swimming time tests
1.3 试验装置与试验方法

图1 鱼类游泳代谢测定仪 Fig.1 The structure of fish swimming-MO2 determining device
目前,水槽实验中最常用的装置均来自于Brett[27]式呼吸仪的改进设计。本研究小组从丹麦购置了一套全自动数字化游泳代谢测定仪(图1),该装置为一个横截面为矩形的循环式水槽, 具有流速、溶氧、光照和温度均可控的特点,工作区的上游有一个控流装置, 下游为不锈钢遮挡网,受试鱼在工作区做趋流运动,水槽外部安装有监视设备,对鱼的游泳行为、状态进行记录。研究小组利用该装置能对各种适应条件进行量化调控,研究初始适应流速、适应时间、加速适应和测试流速对鱼类游泳的续航能力产生的影响。试验在专用鱼类游泳代谢测定仪的环形水槽中进行(丹麦进口,图1),工作区(测试区)观测窗口长46 cm,水流断面尺寸14 cm×14 cm,测试系统造流流速范围(0—1.5)m/s。测试区的上下游设有网状稳流装置,整个环形水槽被封闭于自动控温、循环过滤和充气系统之中。
试验前,给整个测试系统注入爆气24 h以上的自来水,并调控水温在(16.0± 0.2) ℃,打开增氧和循环系统,每次试验随机取2尾鱼(每个水平的测试设3个重复组,分别编号为A—F),放入试验区并让其适应1 h 以降低环境更替带来的压力,期间水流速度控制在0.06 m/s。测试开始时,按试验方案,将测试区的流速在1 min 内升至设定的适应流速,并使鱼在该流速下趋流到设定的适应时间,随后将流速在对应的加速时间内调整到测试流速。以测试方案的第1组为例: 适应环境1 h后,流速由0.06 m/s 升至适应流速0.09 m/s,并在该流速下适应5 min,然后在8 min内将流速匀速升至对应的测试流速0.40 m/s。当试验对象接触试验区末端阻隔网,放弃趋流30 s时视为试验结束[21]。
1.4 试验鱼体力消耗(E)公式应用
本研究中,由于作者对试验鱼采用不同的加速时间组合,因此,鱼的体力消耗也存在差异。借鉴桂福坤等[21]的方法,按下式计算试验鱼在加速阶段的体力消耗情况:
(1)
式中,E代表体力消耗比例,t1表示加速初始时间(t1= 0),t2表示加速结束时间,T表示t时刻对应的续航时间。由于本试验采用均匀加速,因此速度与时间的关系式为:
(2)
式中,Vt1为加速起始对应的适应流速,Vt2为t2对应的测试流速,ΔT为加速时间。续航时间与测试速度采用幂函数模型:
T=aVb
(3)
式中,a和b为进行线性相关回归分析得到的幂函数常数。将公式(2)和(3)带入公式(1),可得最终体力消耗公式:
(4)
1.5 数据处理与分析
试验计时采用时钟和数码摄像机(Sony DCR-SR200E)进行全程记录,作为计时分析和测试结束的判定依据。对于测试时间较短的计时(<600 min),通过摄像机回放确定续航时间;对测试时间较长的计时,采用时钟作为计时手段,数据采用均值作为分析依据。适应时间、流速和加速时间以及测试流速对试验鱼的续航时间与体力消耗的影响采用向后剔除回归法(Regression Backward)进行统计分析(软件为SPSS 18.0),以筛选出影响续航时间和体力消耗的最关键因子。
2 结果与分析
2.1 适应条件对细鳞鲑幼鱼续航时间的影响
在8组不同组合的测试方案下,细鳞鲑幼鱼的续航时间结果如表2所示。本试验的每个组合为3个重复,个别数据与其他数据偏差过大将被舍弃。通过表2统计发现,细鳞鲑幼鱼的最大续航时间为768 min(测试流速为0.25 m/s),最小续航时间为2.4 min(测试流速为0.60 m/s);续航时间随着测试流速的增大而减小。
利用相关函数模型分析各适应因子对细鳞鲑幼鱼续航时间的影响,各因子单独对续航时间的影响以及两者之间的拟合曲线见图2—图5。从各个拟合的趋势线可以看出,只有测试流速与续航时间具有密切的相关性,且呈显著的幂函数关系,即续航时间随测试流速的增加而迅速缩短。
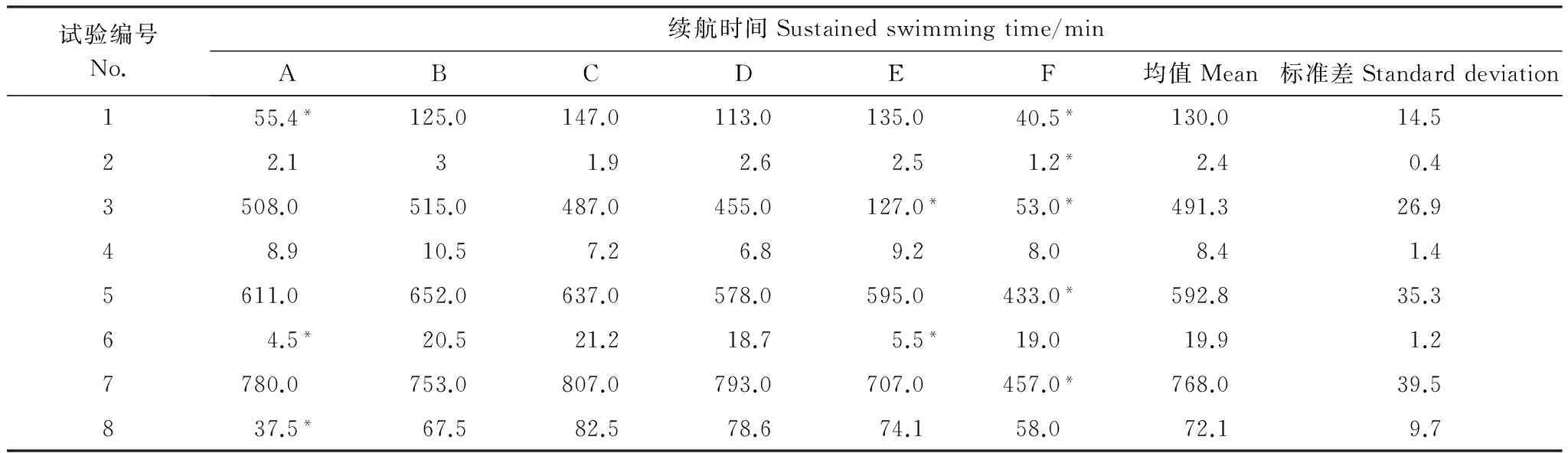
表2 细鳞鲑幼鱼续航时间测试结果Table 2 Experimental sustained swimming time of juvenile lenok
*为淘汰数据
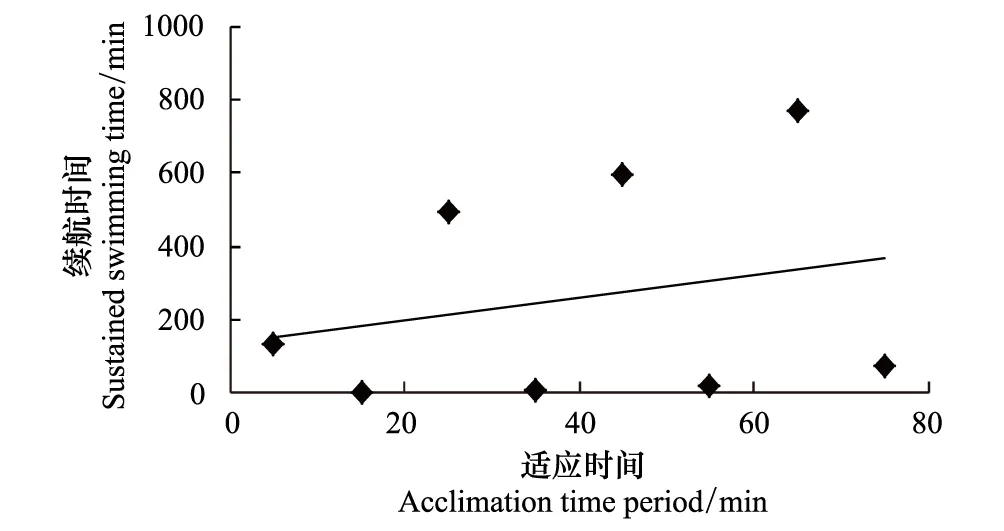
图2 适应时间与细鳞鲑幼鱼游泳续航时间的关系 Fig.2 Relationship between acclimation time period and the sustained swimming time of juvenile lenok
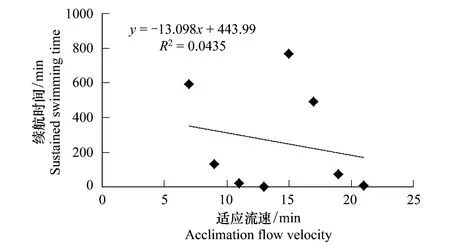
图3 适应流速与细鳞鲑幼鱼游泳续航时间的关系 Fig.3 Relationship between acclimation flow velocity and the sustained swimming time of juvenile lenok
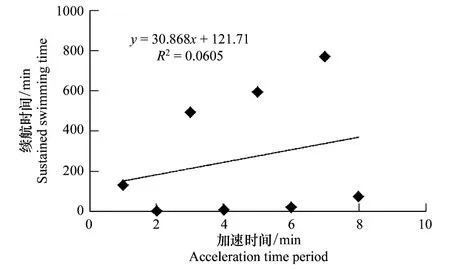
图4 加速时间与细鳞鲑幼鱼游泳续航时间的关系 Fig.4 Relationship between acceleration time period and the sustained swimming time of juvenile lenok
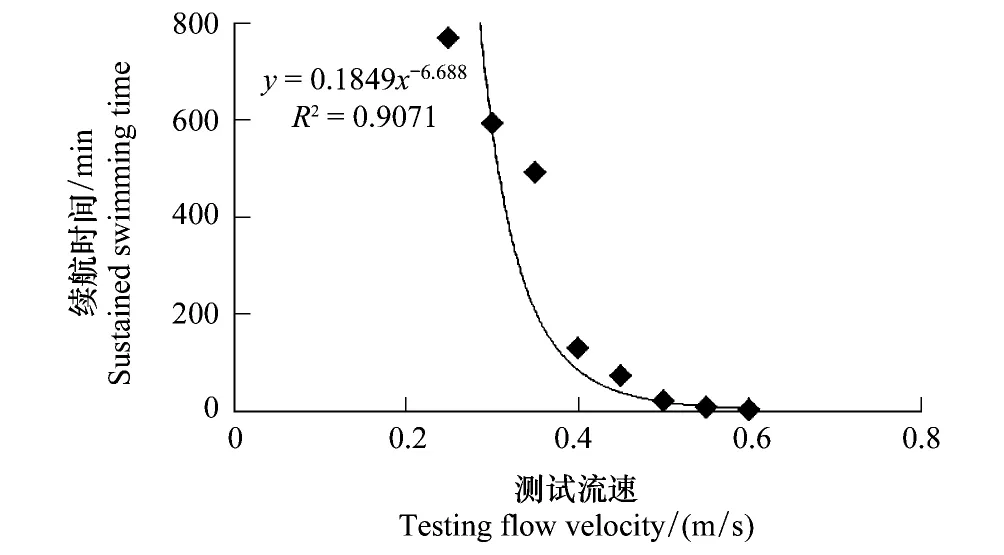
图5 测试流速与细鳞鲑幼鱼游泳续航时间的关系 Fig.5 Relationship between testing flow velocity and the sustained swimming time of juvenile lenok
2.2 适应条件对续航游泳时间的显著性分析
虽然大多适应条件与续航时间之间未呈现出某种最佳的线性关系,但并不代表其对续航时间没有影响,只是影响程度存在差异。因此,主要采用多元线性回归的向后剔除法来分析各适应因子对续航时间的影响程度以及相关性。根据图2—图5中各个模型的初步判断,只有测试流速与续航时间具有相关的幂函数关系(图5),因此根据向后剔除回归的运算要求,要将测试流速进行幂函数进行转换处理,即转换测试流速(V′)按照公式V′=V-6.688计算,其他适应条件数据不做处理,直接带入运算。之后依次将适应时间、适应流速、加速时间、V′和续航时间相应数据导入统计软件中,应用线性回归的向后剔除法进行分析,进而检验各因子对续航时间的影响程度,统计结果见表3。结果分析表明,测试流速在各阶元回归中,对细鳞鲑幼鱼续航时间的影响极为显著(P<0.01),而其他适应因子对续航时间的影响均不显著(P>0.05)。表3中测试流速的回归系数与图5中的回归系数不一致,主要是对测试流速进行转换处理而导致的,不影响最终的结果分析。
分析中测试流速按照V′=V-6.688进行换算处理
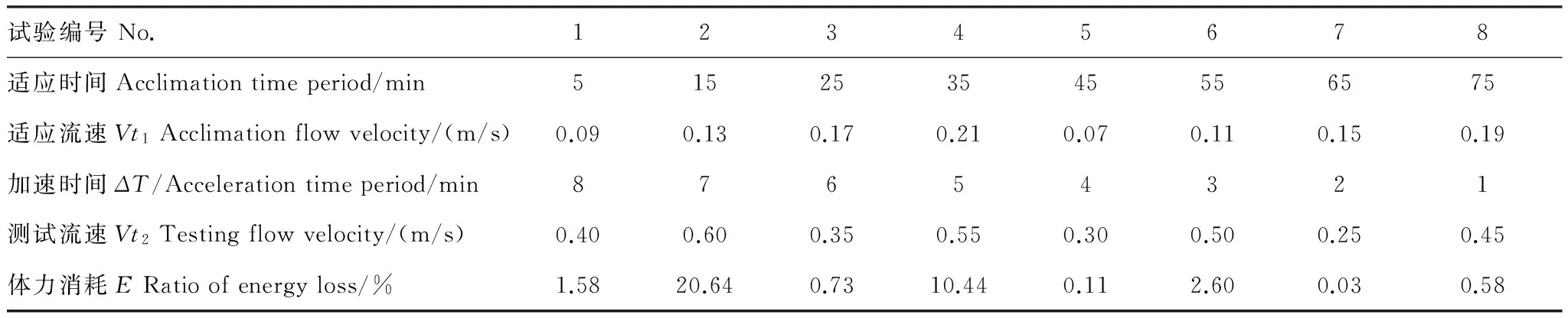
表4 细鳞鲑幼鱼加速过程中的体力消耗情况Table 4 Ratio of energy loss of juvenile lenok during acceleration process
2.3 不同适应因子对细鳞鲑幼鱼体力消耗的影响
在不断加速的过程中,各个因子对幼鱼体力消耗的影响见表4,各试验组的体力消耗情况存在差异,第2组和第4组的体力消耗均超过了10%;第1组和第6组的体力消耗超过了1%;剩余其他各组的体力消耗均小于1%,可以忽略不计。依据向后剔除回归分析的结果(表5),加速时间与测试流速是影响试验鱼体力消耗的相关的条件因子,且测试流速是直接决定试验鱼体力消耗大小的关键因子;而测试流速在0.25—0.45 m/s时,体力消耗最少。本试验的最大适应流速为0.19 m/s,利用续航游泳时间与测试流速的幂函数关系(图5) 进行换算,细鳞鲑幼鱼的续航游泳时间应为12320.63 min;而在此适应流速下的适应时间为75 min,仅占理论总续航时间的0.61%,其它适应流速条件下适应时间对应的体力消耗情况(仍以续航时间比例作为分析依据) 如表6所示。
3 讨论
3.1 适应时间和适应流速对细鳞鲑续航游泳时间的影响
通过表6可以发现,本研究与桂福坤等[21]的研究结果相一致,适应时间与适应流速对续航游泳时间的影响极其微弱(表6),体力消耗最大的情况仅为0.55%(适应流速为0.21 m/s,适应时间为35 min),该体力消耗可以忽略,因此不会对最终的续航游泳时间的测定结果产生显著影响,这与在显著性分析部分的结论相吻合。
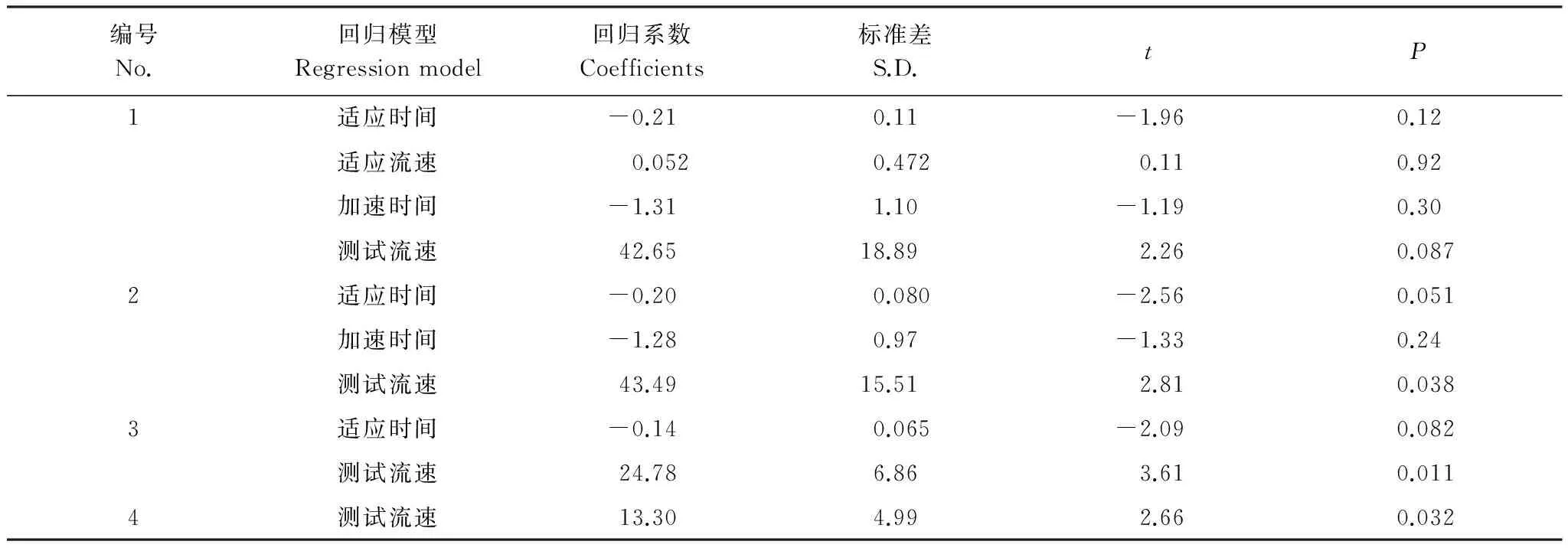
表5 加速过程中细鳞鲑幼鱼体力消耗与不同适应条件的向后线性回归分析Table 5 Backward linear regression analysis for the energy loss of lenok during acceleration process
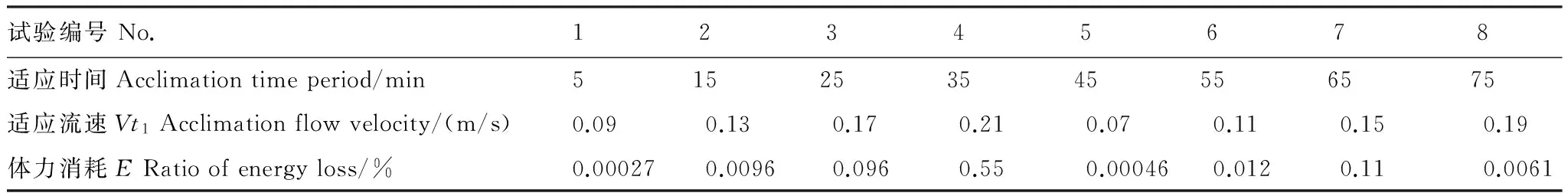
表6 细鳞鲑幼鱼适应时间与适应流速条件下相对的体力消耗情况Table 6 Ratio of energy loses of juvenile lenok during flow velocity acclimation stage
但按照上述方法,若将适应流速调整为1.5—2 BL/s适应1 h时间的情况下,细鳞鲑幼鱼的体力消耗将达到10%—70%。若仍以10%作为体力消耗比例的控制标准[21],细鳞鲑幼鱼在1 h适应时间内的最大适应流速应为1.5 BL/s(即0.30 m/s);或适应流速为1.0—1.5 BL/s条件下的最长适应时间应不超过1 h。大多学者常采用的1 h作为鱼类的适应时间[8,28],我们在进行相关游泳能力测定时,建议鲑科鱼类幼鱼的流速最好不超过1.0 BL/s,在适当流速情况下适应时间可根据试验需求确定,1—2 h均不影响最终试验结果,但根据大量试验摸索发现,适应流速的初始值应设定为微流水状态(<0.06 m/s);在试验开始前的适应环境的时间应设定在1 h为最佳,最多不超过2 h。国内外对于鲑科鱼类适应条件的研究尚处于空白,但条件因子对于试验结果的影响显著,还需进一步深入研究形成一致的试验惯例。
3.2 加速时间对细鳞鲑幼鱼续航游泳时间的影响
适应条件对鱼类游泳能力的影响具有两面性[28],足够的适应可使受试鱼的身体机能得以充分地调整,直至最佳游泳状态,进而延长了其续航时间,即积极的影响;但过度的适应也会使受试鱼消耗大量体力,导致其续航时间被缩短,即消极的影响。这两方面的影响在整个适应过程中是相伴存在的,针对不同种类鱼进行大量实验摸索,找出各适应条件对续航游泳时间影响的有效范围,是确保测定鱼类游泳能力研究结果准确和可靠的基本前提,也可为不同种类间的试验结果比较分析提供强有力的理论依据。对于加速时间而言,缓慢而匀速的过程对于试验是利好的,但过慢同样会浪费大量不必要的体力;但过快情况下鱼类将无法迅速调整自身的生理机能以适应不断变化的流速环境,进而使其始终处于紧张和胁迫状态下,导致机体浪费大量体力来做调整所用。多数研究人员都希望鱼有足够的适应时间,但又不想让实验鱼在此过程中消耗过多的体力,因此,通过大量科学合理的试验探索,筛选出合适的条件因子,进而指导鱼类游泳能力的相关研究是必要的。本研究基本获得了与桂福坤等[21]相一致的研究结果,通过不同适应因子与续航游泳时间和体力消耗的向后剔除回归相关分析发现,尽管测试流速是其最关键的条件因子,但在相应的测试流速情况下,加速时间对续航游泳时间的影响最为敏感。因为在较大测试流速条件下,过长的加速时间会导致体力消耗过大,从而影响了测试结果,如细鳞鲑幼鱼、美国红鱼以及黑鲷的测试流速分别在0.6、1.16 m/s和0.86 m/s时,加速时间均为7 min,这3条鱼在加速过程中的体力消耗分别占20.64%、19.40%和15.2%[21],而这3条鱼在其它流速条件下的体力消耗相对较小,这也证明了回归分析的正确性,即加速时间对续航时间的影响总体并不显著。
通过大量分析与比较研究发现,在鲑鱼类续航游泳能力试验中,幼鱼的最大测试流速一般应控制在3—4 BL/s,且在大流速情况下,应避免其体力消耗过大,需根据预试验设定相对较短的加速时间,但加速时间应考虑鱼的种类、规格、年龄以及测试流速等情况,视具体情况而定[21];小流速情况对其体力消耗影响不大,加速时间的选择范围也较大。根据本研究的结果,如以10%体力消耗作为控制标准,对细鳞鲑幼鱼而言,在最大测试流速下的适应时间应控制在1 h、其加速时间应控制在4 min以内。
3.3 测试流速对细鳞鲑幼鱼续航时间的影响
通过表1发现,细鳞鲑在幼鱼阶段游泳能力不是特别强,测试流速达到0.6 m/s时,续航时间仅能达到2.4 min,这符合该鱼的早期生活史主要是在北方地区江河的支流或山间溪流的栖息习性[29]。细鳞鲑经过长期进化,其主要的栖息地在溪流中,因此其游泳能力不是特别强,但冲刺能力较好。由于该鱼自身的特点,在设定测试流速时,应充分考虑最大测试流速极值,根据续航时间结果和体力消耗的情况,本研究认为,鲑科鱼幼鱼阶段的最大测试流速应设置在3—4 BL,因为通过之前体力消耗比例公式测算,如果测试流速过大,将直接影响其续航时间的测定结果。根据上述分析与大量试验摸索我们还发现,对于鲑科鱼类幼鱼的续航游泳能力的测定,首先应对试验鱼进行初步筛选,因为鱼类属于群体性动物,存在个体差异(适应能力),为了使测定的游泳能力能尽量接近群体的真实情况,所以应先通过预试验找到一个测试速度标准,即在该测试流速下能自然趋流游泳,且游泳状态正常(无挣扎与拒绝游泳情况)3—5 min,如能通过该标准测试,即视为合格,因为有些个体适应环境的能力很差,必须采用超长的适应环境时间才能进行正常游泳测定,但由于不能浪费大量宝贵时间以及体力消耗,所以我们要进行初步筛选。本研究将细鳞鲑幼鱼的最小测试流速通过预试验判定为0.25 m/s。在自然界中,鱼类的运动是一系列复杂的游泳状态的综合体,常与阶段性的持续式游泳运动、暂停及偶尔性的爆发游泳运动相互穿插发生[30-31]。以鲑科鱼类生殖洄游为例,爆发游速为其越过水流障碍到达产卵场提供了保障,持续式和耐久式游泳状态则在长距离洄游中发挥重要作用[32]。而通常鱼类在季节性洄游或迁徙时还会面临大型水坝设施,他们只能借助鱼道通行。因此,鱼道内的设计流速通常按鱼类的耐久游泳速度设计,在修建较长的鱼道时,通过鱼类的耐久游泳能力计算其最大游泳距离,以此确定休息池的距离。尤其对于鲑科鱼类的幼鱼,在索饵或迁徙过程中,对于水流障碍的影响更为明显。对于细鳞鲑幼鱼而言,如果想穿越鱼道,那么该鱼道的最大流速不能超过体长4倍,且距离不能超过80m。
[1] Stobutzki I C, Bellwood D R. An analysis of the sustained swimming abilities of pre-and post-settlement coral reef fishes. Journal of Experimental Marine Biology and Ecology, 1994, 175(2): 275-286.
[2] Drucker E G. The use of gait transition speed in comparative studies of fish locomotion. American Zoologist, 1996, 36(6): 555-566.
[3] Watkins T B. Predator-mediated selection on burst swimming performance in tadpoles of the Pacific tree frog,Pseudacrisregilla. Physiological Zoology, 1996, 69(1): 154-167.
[4] Hammer C. Fatigue and exercise tests with fish. Comparative Biochemistry and Physiology Part A: Physiology, 1995, 112(1): 1-20.
[5] Plaut I. Critical swimming speed: its ecological relevance. Comparative Biochemistry and Physiology Part A: Physiology, 2001, 131(1): 41-50.
[6] 王萍, 桂福坤, 吴常文. 鱼类游泳速度分类方法的探讨. 中国水产科学, 2010, 17(5): 1137-1146.
[7] Fisher R, Bellwood D R. The influence of swimming speed on sustained swimming performance of late-stage reef fish larvae. Marine Biology, 2002, 140(4): 801-807.
[8] Fisher R, Wilson S K. Maximum sustainable swimming speeds of late-stage larvae of nine species of reef fishes. Journal of Experimental Marine Biology and Ecology, 2004, 312(1): 171-186.
[9] Ware D M. Bioenergetics of pelagic fish: theoretical change in swimming speed and ration with body size. Journal of the Fisheries Research Board of Canada, 1978, 35(2): 220-228.
[10] Trump C L, Leggett W C. Optimum swimming speeds in fish: the problems of currents. Canadian Journal of Fisheries and Aquatic Sciences, 1980, 37(7): 1086-1092.
[11] Hammer C. Effects of endurance swimming on the growth of 0-and l-age group of whiting,Merlungiusmerlangus, Gadidae. Archive of Fishery and Marine Research, 1994, 42(2): 105-122.
[12] Fisher R, Bellwood D R, Job S D. Development of swimming abilities in reef fish larvae. Marine Ecology Progress Series, 2000, 202: 163-173.
[13] Rodríguez T T, Agudo J P, Mosquera L P, Mosquera L P, González E P. Evaluating vertical-slot fishway designs in terms of fish swimming capabilities. Ecological Engineering, 2006, 27(1): 37-48.
[14] Jain K E, Hamilton J C, Farrell A P. Use of a ramp velocity test to measure critical swimming speed in rainbow trout (Onchorhynchusmykiss). Comparative Biochemistry and Physiology Part A: Physiology, 1997, 117(4): 441-444.
[15] Farrell A P. Comparisons of swimming performance in rainbow trout using constant acceleration and critical swimming speed tests. Journal of Fish Biology, 2008, 72(3): 693-710.
[16] 田凯, 曹振东, 付世建. 速度增量及持续时间对瓦氏黄颡鱼幼鱼临界游泳速度的影响. 生态学杂志, 2010, 29(3): 534-538.
[17] Foster J J. The influence ofsh behaviour on trawl design with special reference to mathematical interpretations of observations on the swimming speeds ofsh and results of C.F. experiments. Proceedings of the FAO Conference on Fish Behaviour in Relation to Fishing Techniques and Tactics, Bergen, 1969:19-27
[18] Beamish F W H. Swimming capacity//Hoar W S, Randall D J. Fish Physiology. vol. 7. New York: Academic Press, 1978: 101-187.
[19] Wardle1 C S, He P. Burst swimming speeds of mackerel,ScomberscombrusL. Journal of Fish Biology, 1988, 32(3): 471-478.
[20] Brett J R. Swimming performance of sockeye salmon (Oncorhynchusnerka) in relation to fatigue time and temperature. Journal of the Fisheries Research Board of Canada, 1967, 24(8): 1731-1741.
[21] 桂福坤, 王萍, 吴常文. 适应条件对鱼类续航游泳能力的影响. 水产学报, 2010, 34(8): 1227-1235.
[22] 郑金秀, 韩德举, 胡望斌, 王翔, 张晓敏. 与鱼道设计相关的鱼类游泳行为研究. 水生态学杂志, 2010, 3(5): 104-110.
[23] 刘洋, 徐革锋, 陈玉春, 李永发, 牟振波. 细鳞鱼气单胞菌的分离、鉴定及药敏试验. 大连海洋大学学报, 2011, 26(3): 277-280.
[24] 徐革锋, 刘洋, 李永发, 牟振波. 不同投喂率对细鳞鲑(Brachymystaxlenok)幼鱼生长及体成分的影响. 海洋与湖沼, 2013, 44(2): 433-437.
[25] 马波, 尹家胜, 李景鹏. 黑龙江流域两种细鳞鲑的形态学比较及其分类地位初探. 动物分类学报, 2005, 30(2): 257-260.
[26] Fang K T. Uniform Designs, Encyclopedia of Statistics. 2nd ed. New York: Wiley, 2004.
[27] Brett J R. The respiratory metabolism and swimming performance of young sockeye salmon. Journal of the Fisheries Research Board of Canada, 1964, 21(5): 1183-1226.
[28] Zeng L Q, Cao Z D, Fu S J, Peng J L, Wang Y X. Effect of temperature on swimming performance in juvenile southern catfish (Silurusmeridionalis). Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2009, 153(2): 125-130.
[29] 解玉浩. 东北地区淡水鱼类. 沈阳: 辽宁科学技术出版社, 2007: 317-317.
[30] Castro-Santos T. Optimal swim speeds for traversing velocity barriers: an analysis of volitional high-speed swimming behavior of migratory fishes. Journal of Experimental Biology, 2005, 208(3): 421-432.
[31] Kieffer J D. Perspective-Exercise in fish: 50+years and going strong. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 2010, 156(2): 163-168.
[32] Powers P, Orsborn J. New concepts in fish ladder design: analysis of barriers to upstream fish migration, volume IV of IV. Investigation of the physical and biological conditions affecting fish passage success at culverts and waterfalls. Project No. 198201400(BPA Report DOE/BP-36523-1). Portland, OR: Bonneville Power Administration, 1985.
Effects of different acclimation conditions on sustainable swimming performance of juvenile lenok,Brachymystaxlenok(Pallas)
XU Gefeng1,2, YIN Jiasheng1, HAN Ying2, MA Bo1, MOU Zhenbo1,*
1HeilongjiangRiverFisheriesResearchInstitute,ChineseAcademyofFisherySciences,Harbin150070,China2CollegeofAnimalScienceandTechnology,NortheastAgriculturalUniversity,Harbin150030,China
Swimming performance is considered a critical factor in determining the survival of many fish species under natural conditions. Flume experimentation, which is visible, measurable, and repeatable for the adjustment and control of environmental factors, is a basic and effective method for the study of swimming performance and behavior in fish. However, many current studies of fish behavior have focused solely on the process of adaptation, while other influencing factors such as temperature, light, dissolved oxygen, velocity, and time have been generally ignored. In addition, maximal sustained swimming time and maximal sustained swimming speed are accepted as key indicators for determining swimming performance in fish. Swimming performance is assessed by determining the maximum sustained swimming performance at a given time and constant speed, and maximal sustained swimming speed is used as the base-line reference when fish become fatigued. The lenok,Brachymystaxlenok(Pallas), is one of the rare and valuable salmonid species found in the northeast of China. In recent years, the biology, ecology, reproduction success, disease prevention, and culture techniques of this species have been extensively studied and reported. However, to the best of our knowledge, there have been very few studies on the behavioral ecology of this species. In this report, the effects of acclimation time, acclimation flow velocity, acceleration time, and tested flow velocity on the sustained swimming time of juvenile lenok were investigated at a water temperature of 16.0 ± 0.2 ℃ and dissolved oxygen level of 8 mg/L. The observed results served as reference data on the ethology, kinematics, energetics, and behavioral ecology of this species. Uniform experimental design was used with four factors and eight levels being applied to all tests. The results revealed that the maximal acclimation flow velocity (AFVmax) should be limited to 1.0—1.5 (body lengths) BL/s, with a corresponding optimum acclimation time (AT) of 1 h. Under reasonable conditions (AFVmax≤ 1.0—1.5 BL/s,AT≤ 1 h), the influence of acclimation flow velocity and acclimation time on sustainable swimming performance was negligible. Acceleration time significantly affected sustained swimming time under stronger flow velocities (> 0.5 m/s) (P< 0.05). Although a longer time period may be favorable in allowing fish to adjust to a new environment, an acclimation time of 1—2 h is recommended; screening for a suitable acclimation time should be performed before trials. It is recommended that the tested flow velocity be limited to 3—4 BL/s to avoid the maximum swimming tolerance range. Sustained swimming time was significantly affected by the tested flow velocities: sustained swimming time decreased significantly with an increase in flow velocity (P< 0.05). The energy loss in fish was also affected by tested flow velocity, acceleration time, and acclimation time. Under the maximal tested flow velocity (> 0.5 m/s), the maximal acclimation time and the acceleration time should be limited to 60 and 4 min, respectively. In conclusion, this study provides a scientific baseline for the behavioral ecology, kinematic theory, and energetics ofB.lenok, which will prove valuable for the design and construction of fish-pass facilities.
Brachymystaxlenok(Pallas); acclimation time period; acclimation flow velocity; acceleration time period; sustained swimming time
中央级公益性科研院所基本科研业务费专项(HSY201312); 国家“十二五”农村领域科技计划(2012BAD26B05)
2013-05-24;
日期:2014-04-25
10.5846/stxb201305241165
*通讯作者Corresponding author.E-mail: mouzhenbo@163.com
徐革锋,尹家胜,韩英,马波,牟振波.不同适应条件对细鳞鲑幼鱼游泳能力的影响.生态学报,2015,35(6):1938-1946.
Xu G F, Yin J S, Han Y, Ma B, Mou Z B.Effects of different acclimation conditions on sustainable swimming performance of juvenile lenok,Brachymystaxlenok(Pallas).Acta Ecologica Sinica,2015,35(6):1938-1946.
猜你喜欢
水产养殖(2022年1期)2022-03-03
水产学杂志(2021年5期)2021-11-29
江苏农业科学(2017年23期)2018-01-29
动物营养学报(2015年10期)2015-12-01
动物营养学报(2015年10期)2015-12-01
应用海洋学学报(2014年1期)2014-11-22
河北渔业(2014年5期)2014-07-08
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28
智慧与创想(2013年6期)2013-08-27
校园足球(2010年3期)2010-08-15
