
秦岭中段南坡油松林生态系统碳密度
2015-03-11 06:47党坤良武朋辉朱成功
生态学报 2015年6期
沈 彪,党坤良,武朋辉,朱成功
西北农林科技大学 林学院, 杨凌 712100
秦岭中段南坡油松林生态系统碳密度
沈 彪,党坤良*,武朋辉,朱成功
西北农林科技大学 林学院, 杨凌 712100
在秦岭中段南坡油松林分布较为广泛的不同区域,采用典型取样的方法设置油松林标准地50块。通过样地调查和室内分析,对本区油松林生态系统植被层、枯落物层及土壤层有机碳密度进行了研究与估算,分析了油松林生态系统各层次的有机碳密度在不同立地因子下的分布规律。结果表明:秦岭中段南坡油松林生态系统总有机碳密度为150.12 t/hm2,其中土壤碳分库的碳密度占油松林生态系统总碳密度的56.74%,是构成油松林生态系统碳的主体组成部分。植被层碳密度为62.29 t/hm2,占油松林生态系统总碳密度的41.49%,高于我国森林生态系统植被碳密度平均值,且仍有较大的固碳潜力。枯落物层碳密度为2.66 t/hm2,占油松林生态系统总碳密度的1.77%。在植被碳分库中,乔木层碳密度是其主体构成部分,为61.22 t/hm2,占植被层碳密度的98.30%;灌木层、草本层碳密度及其所占植被层碳密度的比例分别为:0.65 t/hm2(1.04%)、0.41 t/hm2(0.66%)。碳在乔木不同器官中的分配大小顺序为:树干(55.82%)、树枝(21.25%)、树根(10.28%)、树叶(7.35%)、树皮(5.30%)。灌木层碳密度和草本层碳密度受地形因子影响不显著。随海拔的升高,乔木层碳密度呈先增后减的变化趋势,在海拔1500—1700 m处达到最大值,枯落物层碳密度、土壤层碳密度及总碳密度变化不显著;随着坡度的增大,油松林生态系统枯落物层碳密度、土壤层碳密度及总碳密度显著减小,乔木层碳密度呈先增后减的变化趋势,在坡度为26—35°范围达到最大值;下坡位土壤层碳密度高于中坡位和上坡位,而中坡位乔木层碳密度和生态系统总碳密度高于下坡位和上坡位,枯落物层碳密度受坡位影响不明显;阳坡乔木层碳密度大于阴坡,枯落物层碳密度、土壤层碳密度及总碳密度受坡向影响不明显。
碳密度;油松;生态系统;秦岭中段南坡
温室效应正在使地球以前所未有的速度变暖,目前,大气中CO2的含量比过去6.5万年中任何时期都高,比工业革命前高了35%[1]。森林是陆地生态系统的主体,同时也是陆地生态系统最大的碳库,陆地生态系统有机碳地上部分的80%和地下部分的40%存储于森林中[2]。森林通过同化作用吸收并固定大气中的CO2,抑制其浓度上升的功能对于缓解全球变暖具有不可替代的作用和地位。森林作为一个动态的碳库,其贮碳能力不仅取决于森林的面积,还取决森林的质量,即单位面积的森林碳密度[3]。秦岭既是我国的主要林区之一,也是我国中部的重要生态屏障,对秦岭林区不同森林生态系统碳储量和碳密度特征的研究和估算,不仅能够准确估算秦岭林区的碳储量,而且对我国森林碳汇研究具有重要的意义。
关于秦岭林区森林碳储量与碳密度及其分配已有学者进行过部分研究[4-8],但现有研究主要集中在火地塘林区等几个面积较小的区域内,并且对于森林生态系统碳密度在不同地形因子下的分布状况少有研究。在较大尺度上对不同地形因子下森林生态系统碳密度分布状况的研究对研究森林生态系统碳循环以及森林生态系统碳储量的准确估算具有重要意义。本研究旨在通过样地调查及生物量和含碳率的测定,较准确地估算秦岭中段南坡油松林生态系统的碳密度,同时分析秦岭南坡中段油松林生态系统不同层次的碳密度与立地因子间的关系,为研究秦岭林区乃至全国森林生态系统碳库特征提供参考。
1 材料与方法
1.1 研究区概况
秦岭中段是指秦岭位于陕西省中部的一部分。研究区位于陕南秦巴山地北部,地理位置位于32°53′—33°57′N,105°53′—111°01′E,总面积484 km2。海拔700—3071 m。年平均气温11.5 ℃,极端高温41.3 ℃,极端低温-25.1 ℃。年降水总量563—1274 mm,全年降水日数93—140 d。无霜期210 d, 植物生长期130—206 d。区内植被属山地暖温带典型落叶阔叶林带,森林主要分布于中高山区的中西段,绝大部分为次生林。由于本区森林在20世纪50—70年代经历了过度采伐和利用,因此原生植被几乎荡然无存,到20世纪90年代,森林基本恢复,形成大面积的天然次生林和人工林[9]。
油松属阳性、喜光、耐旱耐瘠薄树种,在秦岭中段南坡海拔1000—2200 m范围普遍分布。伴生树种主要为锐齿栎(Ouercusalienavar.Acuteserrata)、华山松(Pinusarmandi)、红桦(Betulaalbo-sinensis)、青榨槭(Acerdavidii)、千金榆(Carpinuscordata)等。根据陕西省2006年森林资源清查资料,在秦岭林区,油松林分布面积为34.55km2,以天然林为主,中幼龄林占油松林总面积的百分之九十以上。
1.2 研究方法
1.2.1 样地设置
本研究以秦岭中段南坡油松林分布较广、面积较大的陕西省太白林业局黄柏塬林场(20块)、西北农林科技大学火地塘试验林场(10块)、陕西省宁东林业局旬阳坝林场(10块)、陕西省宁西林业局菜子坪林场作为研究地点(10块),共布设20 m×20 m的调查样地50块。调查样地设置在每个林区不同立地条件下,林分生长状况可以代表区域内平均水平的油松纯林中。所选林分均为天然中龄林。利用GPS确定每个样地的位置,并记录每块样地海拔、坡度、坡位、坡向等立地因子,进行每木检尺,记录每株乔木的树高、胸径,调查林分郁闭度、灌木与草本盖度、主要植物种类。采集样品。各研究地区及样地概况见表1。
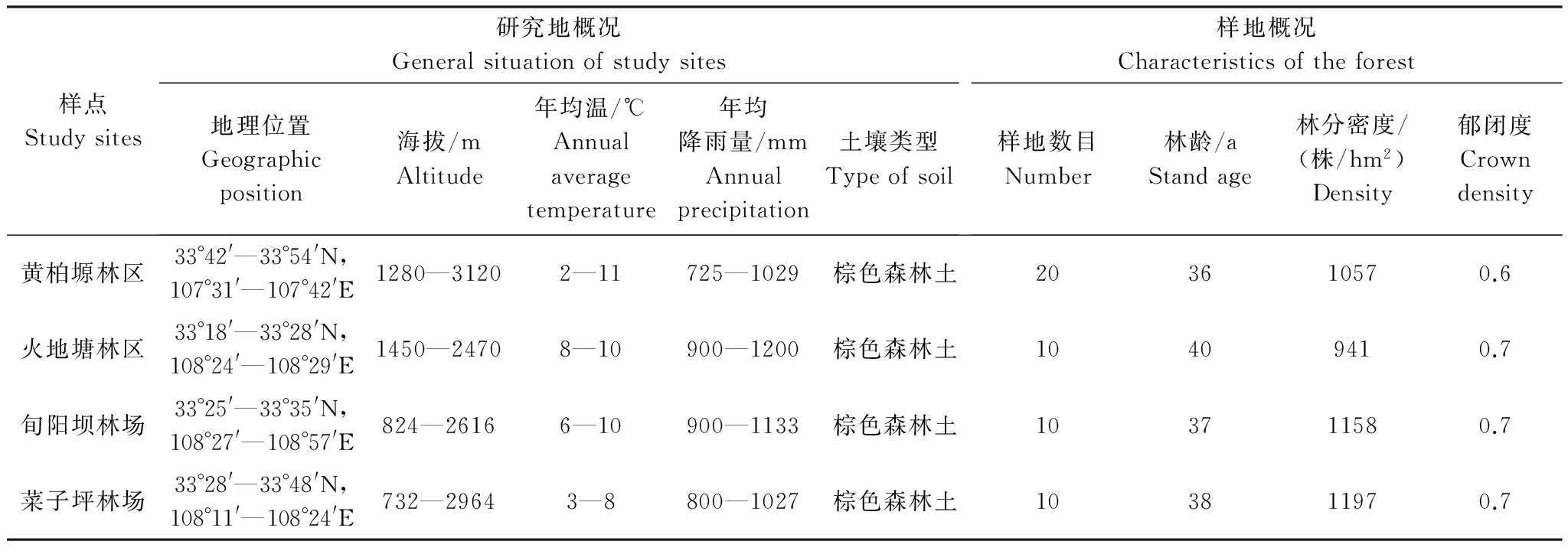
表1 研究地自然条件概况及林分状况Table 1 General situation of study sites and characteristics of the forest
1.2.2 立地因子的划分
按照《西北主要树种培育技术》[10]立地因子等级划分方法,并结合本研究野外调查实际,本文立地因子的划分如下:海拔按1300 m以下、1300 m—1500 m、1500 m—1700 m、1700 m以上划分为4个等级;坡度按平地(≤5°)、缓坡(5°—15°)、斜坡(15°—25°)、陡坡(25°—35°)、急坡(35°—45°)、险坡(>45°)划分为6个等级;坡位按下坡位(地形图上距离坡顶大于2/3 处及其以下的坡面)、中坡位(上坡位与下坡位之间的坡面部分)、上坡位(地形图上距离坡顶1/3 处及其以上的坡面)划分为3个等级;坡向分为阴坡(西北、正北、东北、正东)和阳坡(东南、正南、西南、正西)两个等级。
1.2.3 碳密度的估算
(1)乔木层生物量的估算 以2 cm为一个径阶,在每个样地进行每木检尺。在每个样地采用平均标准木法选取标准木3—5株。本研究共选取标准木184株,伐倒标准木并用收获法挖出其根,分别称量标准木的干(去皮)、皮、枝、叶、根等器官的鲜重。分器官采样并带回实验室85℃烘至恒重,测定各器官的含水率,计算标准木各器官的生物量,进而估算调查样地的乔木各器官的生物量。调查样地乔木层生物量计算公式如下:
(1)
式中,Bj为样地乔木各器官单位面积生物量(t/hm2);j=1,2,3,4,5分别代表代表枝、叶、干、皮、根;Wj标为标准木不同器官的生物量(t);n为株数;S为样地面积(0.04 hm2)。
(2)灌木层、草本层生物量及枯落物量 在每个调查样地内四角和中央分别设置2 m×2 m的灌木调查样地、1 m×1 m的草本调查样地、1 m×1 m的枯落物调查样地,将灌木、草本连根挖出,分别称量灌木根、枝干和叶鲜重,草本地上、地下部分鲜重,枯落物全部收集并称重,分别取样。所有样品带回实验室85 ℃烘至恒重,测定其含水率,进而推算样地内单位面积灌木、草本各器官生物量及枯落物量。
(3)植被与枯落物层碳密度的估算 植被与枯落物所烘干样品用粉碎机粉碎,过200目筛用德国ELEMENTAR公司LIQUIC TOCⅡ总有机碳分析仪测定各部分含碳率,各部分单位面积生物量乘以含碳率即为其碳密度。
(4)土壤碳密度的估算 在每个调查样地内四角及中央各挖取1个土壤剖面,按照土壤自然发生过程分A、B、C3个层次分别采样,记录各层次厚度(本研究中A、B、C 3个层次的平均厚度分别为11、25 cm和30 cm)。用环刀法测土壤容重,挑出其中的植物根,大于2 mm的砾石,测定其含量(体积),另取土样风干后磨碎,过200目筛,用德国ELEMENTAR公司LIQUIC TOCⅡ总有机碳分析仪测定样品含碳率,土壤各层碳密度计算公式如下:
(2)
式中,SOCD为整个土壤剖面土壤有机碳密度(t/hm2),n=3,ρi为第i层土壤平均容重(g/cm3),θi为第i层>2 mm石砾含量(%),ci为第i层土壤含碳率(g/kg),di为第i层土壤厚度(cm)。
各层土壤碳密度之和即为总土壤碳密度。每个样地中5个剖面平均值为样地土壤碳密度。
1.2.4 数据分析
数据处理与分析采用EXCEL及SPSS软件进行。通过相关分析研究生态系统各组分碳密度间的关系,运用方差分析比较不同海拔、不同坡度、不同坡位碳密度差异,差异显著性检验采用LSD法,用独立样本t检验比较不同坡向碳密度差异程度。
2 结果与分析
2.1 秦岭中段南坡油松林生态系统碳的总体分布特征
森林生态系统的碳库由土壤碳分库、植被碳分库和枯落物碳分库三部分组成。通过研究和计算,秦岭中段南坡油松林生态系统碳库总碳密度为89.81—202.35 t/hm2,平均碳密度为150.12 t/hm2。其中土壤碳分库的碳密度为39.97—117.70 t/hm2,平均碳密度为85.17 t/hm2,占油松林生态系统碳库总碳密度的56.74%;植被碳分库的碳密度为18.93—112.22 t/hm2,平均碳密度为62.29 t/hm2,占油松林生态系统碳库总碳密度的41.49%;枯落物层碳密度为0.37—6.50 t/hm2,平均碳密度为2.66 t/hm2,占油松林生态系统碳库总碳密度的1.77%(表2)。

表2 油松林生态系统各组分碳密度Table 2 Carbon density of each component in the Pinus tabulaeformis forest ecosystem
在油松林生态系统土壤碳分库中,A层土壤碳密度为19.30—74.74 t/hm2,平均碳密度为43.07 t/hm2,占土壤碳分库总碳密度的50.56%;B层碳密度为16.27—46.73 t/hm2,平均碳密度为33.31 t/hm2,占土壤碳分库总碳密度的39.11%;C层碳密度为0.51—21.56 t/hm2,平均碳密度为8.80 t/hm2,占土壤碳分库总碳密度的10.33%。
在油松林生态系统植被碳分库中,乔木层碳密度为17.89—111.48 t/hm2,平均碳密度为61.22 t/hm2,占植被碳分库碳密度的98.28%;灌木层碳密度为0.09—2.07 t/hm2,平均碳密度为0.65 t/hm2,占植被碳分库碳密度的1.04%;草本层碳密度为0.03—1.25 t/hm2,平均碳密度为0.41 t/hm2,占植被碳分库碳密度的0.66%。
2.2 乔木层碳在不同器官的分配
在森林生态系统中,不同树种构成的生态系统由于其生物学和生长特性不同,因而乔木层碳在不同器官的分配也具有明显差异。在油松林生态系统中,乔木层树干碳密度为34.18 t/hm2,占乔木层总碳密度的55.82%;枝条碳密度为13.01 t/hm2;占乔木层总碳密度的21.25%;树根碳密度为6.30 t/hm2,占乔木层总碳密度的10.28%;树叶碳密度为4.50 t/hm2,占乔木层总碳密度的7.35%;树皮碳密度为3.24 t/hm2,占乔木层总碳密度的5.30%(表3)。可见,树干是构成油松林生态系统乔木层碳密度的主要组成部分,在碳汇方面具有极为重要的作用。

表3 乔木层不同器官碳密度Table 3 Carbon density of different organs of the arbor
2.3 不同区域油松林生态系统碳密度差异分析
在秦岭中段南坡不同区域,由于各区域气候、土壤和人为活动的影响不同,油松林生态系统总碳密度也存在一定差异,但这种差异在4个林区之间并不显著(P>0.05)(表4)。乔木层碳密度大小顺序为:旬阳坝(71.83 t/hm2)>菜子坪(69.13 t/hm2)>火地塘(65.34 t/hm2)>黄柏塬(49.90 t/hm2),乔木层碳密度黄柏塬林区显著低于其他3个地区,而菜子坪、火地塘、旬阳坝3个林区之间乔木层碳密度差异不显著;灌木层碳密度以菜子坪林区最高,平均为1.09 t/hm2,并与其他3个区域灌木层碳密度存在显著差异;不同林区之间草本层、枯落物层的碳密度差异不显著;4个林区油松林的土壤碳密度非常接近,几乎不存在差异;由此可见,导致不同区域油松林生态系统碳密度差异的主要因素为乔木层碳密度,但总碳密度差异不显著,说明在秦岭中段南坡这一较大范围内进行油松林碳密度的估算可以代表该区域油松林碳密度的总体水平。

表4 各研究地油松林生态系统碳密度(t/hm2)Table 4 Carbon density of Pinus tabulaeformis forest ecosystem in each study sites
不同字母表示差异显著(α=0.05)
2.4 立地因子对油松林生态系统碳密度的影响
2.4.1 海拔对油松林生态系统碳密度的影响
通过统计分析和检验,结果表明在秦岭中段南坡油松林集中分布的海拔范围内,油松林生态系统总碳密度(F3,46=1.364,P=0.266)、土壤层碳密度(F3,46=0.1.277,P=0.293)、灌木层碳密度(F3,46=0.199,P=0.897)、草本层碳密度(F3,46=0.742,P=0.532)和枯落物层碳密度(F3,46=0.329,P=0.804)与海拔具有一定的关系,但这种关系不显著;从碳密度随海拔变化趋势来看,油松林生态系统总碳密度与土壤层碳密度随海拔升高具有增加的趋势(图1)。在不同海拔范围油松林生态系统乔木层碳密度存在极显著差异(F3,46=4.686,P=0.006),在海拔1500—1700 m范围内,油松林生态系统乔木层碳密度显著高于其它海拔范围,达到76.08 t/hm2;当海拔低于1500 m,随着海拔的升高,乔木层碳密度增加,当海拔高于1700 m,乔木层碳密度有下降趋势。
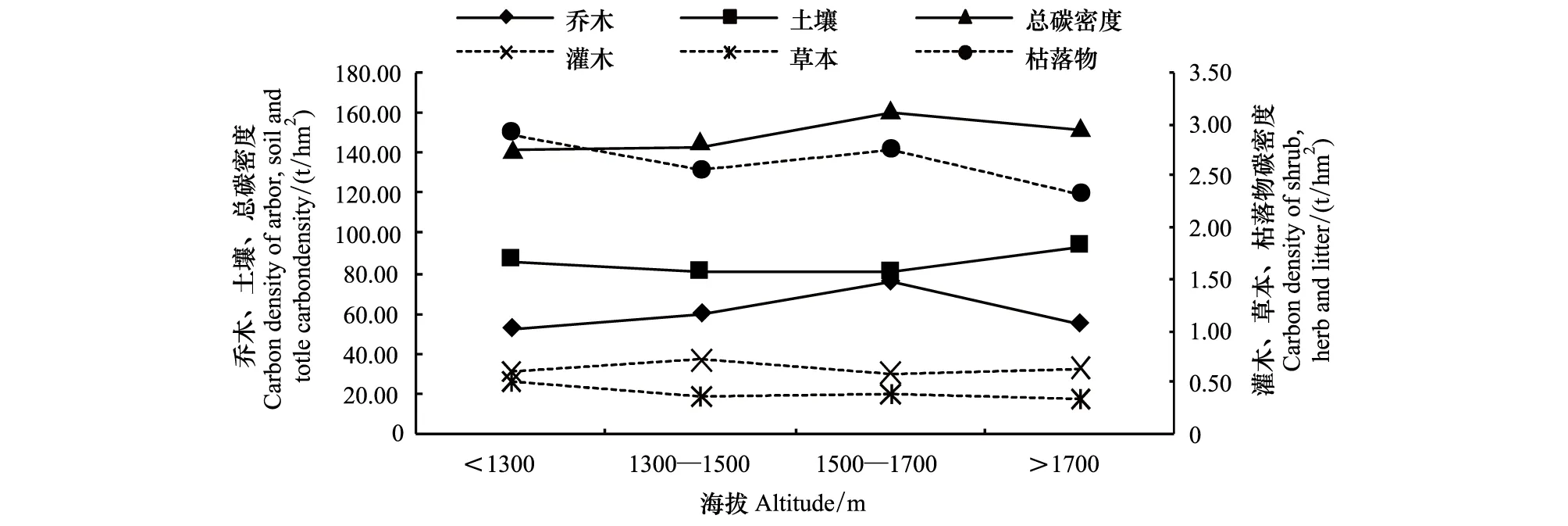
图1 不同海拔油松林生态系统碳密度(t/hm2)Fig.1 Carbon density of Pinus tabulaeformis forest ecosystem of different elevation
2.4.2 坡度对油松林生态系统碳密度的影响
坡度对秦岭中段南坡油松林生态系统的总碳密度(F5,44=8.296,P=0.000)、乔木层碳密度(F5,44=3.342,P=0.012)、枯落物层碳密度(F5,44=4.771,P=0.001)及土壤碳密度(F5,44=16.025,P=0.000)具有极为重要的影响,均呈现出显著相关关系。乔木层碳密度随坡度增大呈先增后减的趋势,斜坡与陡坡乔木层碳密度显著高于其他坡度(P<0.05),在陡坡达到最大值74.23 t/hm2,土壤层碳密度、枯落物层碳密度及总碳密度随坡度的增加而显著减小(图2)。不同坡度灌木层的碳密度(F5,44=1.127,P=0.360)和草本层碳密度(F5,44=0.615,P=0.689)差异不显著。
2.4.3 坡位对油松林生态系统碳密度的影响
通过计算,在秦岭中段南坡,油松林生态系统的总碳密度(F2,47=4.594,P=0.015)、乔木层碳密度(F2,47=5.249,P=0.009)、土壤层碳密度(F2,47=4.815,P=0.013)受坡位影响较为显著(表5)。其中,土壤碳密度下坡位最大,平均为94.62 t/hm2,与上坡位(75.34 t/hm2)差异显著,中坡位(86.42 t/hm2)介于上坡位和下坡位之间。总碳密度和乔木层碳密度都是中坡位大于上坡位和下坡位。不同坡位灌木层(F2,47=0.038,P=0.962)、草本层(F2,47=0.088,P=0.916)、枯落物层(F2,47=1.121,P=0.334)碳密度差异不显著,但枯落物层碳密度平均值大小表现为下坡位(2.98 t/hm2)>中坡位(2.78 t/hm2)>上坡位(2.23 t/hm2)趋势。
2.4.4 坡向对油松林生态系统碳密度的影响
t检验结果表明,不同坡向油松林生态系统乔木层碳密度差异显著(表6),阳坡(68.39 t/hm2)显著高于阴坡(54.61 t/hm2),这与油松具有喜光的生物学特性相吻合。灌木层、草本层、枯落物层以及土壤层碳密度阴坡大于阳坡,总碳密度阳坡大于阴坡,但差异均不显著(P>0.05),说明坡向对油松林生态系统碳密度的影响主要表现在对乔木层碳密度的影响上。

图2 不同坡度油松林生态系统碳密度Fig.2 Carbon density of Pinus tabulaeformis forest ecosystem of different gradient

表5 不同坡位油松林生态系统碳密度Table 5 Carbon density of Pinus tabulaeformis forest ecosystem of different slope position
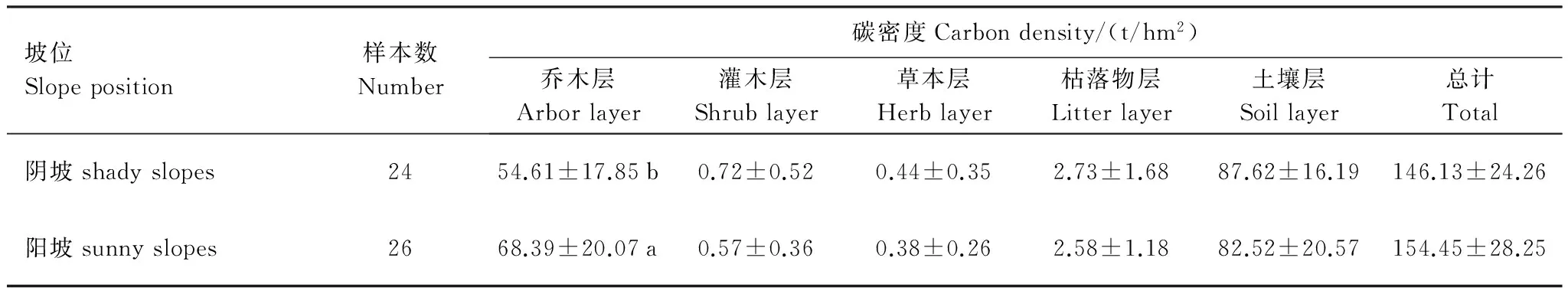
表6 不同坡向油松林生态系统碳密度Table 6 Carbon density of Pinus tabulaeformis forest ecosystem of different slope aspect
3 讨论
3.1 油松林生态系统各组分的碳密度
研究结果表明,秦岭中段南坡油松林生态系统碳密度平均为150.12 t/hm2,乔木层与土壤层为生态系统碳库的主体,占生态系统总碳密度的97%以上。其中植被层碳密度为62.29 t/hm2,与邓蕾等人通过二类森林调查资料估算的宁陕县油松林碳密度(63.90 t/hm2)[8]相近,比李海奎等[11]人估算的中国油松林平均碳密度(20.94 t/hm2)高出41.35 t/hm2,比赵敏(41.32 t/hm2)[12]、徐新良(38.56 t/hm2)[13]、方精云(41.0 t/hm2)[14]等人估算的我国森林植被平均碳密度分别高出20.97 t/hm2、23.73 t/hm2、21.29 t/hm2。可见该地区油松林乔木层碳密度与全国森林碳密度平均值相比处于相对较高水平,但仍低于世界平均值(86 t/hm2)[15]。土壤碳密度平均为85.17 t/hm2,约为植被层的1.4倍,相对于全球森林土壤碳储量约为植被碳储量的2.2倍[15]而言较低,这主要是因为秦岭林区地形以山地为主,山势陡峭,油松林多分布在土层较为瘠薄的地段所致。油松喜温凉气候,由表1可以看出本区油松主要分布区年均温在2—11℃,年降水充沛,土壤为棕色森林土,气候环境较为适合油松生长;本区平均林龄35—40a,以中幼龄林为主,因而仍有较大的碳汇潜力。
本研究结果表明秦岭中段南坡油松林乔木层碳密度在不同器官中分布不均,主要集中在树干(55.82%)、树枝(21.25%)和根系(10.28%)中。这与程堂仁[16]等在小陇山和任军辉等[17]在贺兰山的研究结论一致。树皮与树叶占乔木层碳密度的比例很小,虽在不同地区存在差异但差别不大。本研究中树干碳密度在乔木层碳密度中所占的比重(55.82%)高于小陇山(48.76%)与贺兰山(44.95%)。乔木层碳密度在不同器官中的分配与树高、胸径以及根冠比等有关。这些生长指标受立地条件以及树木年龄的影响。小陇山林区年均降水量460—900 mm,蒸发量989—1658 mm,贺兰山林区年均降水量420 mm,相对缺水的环境使得油松必须扩大根系,以获取足够的水分,根系在小陇山林区与贺兰山林区油松林碳密度所占的比重分别为17.78%和17.43%,明显高于本研究(10.28%)。由于树干不参与凋落过程,其所固定的碳可以在较长时区内储存于森林中。较高的树干碳密度有利于碳的长期固定,因此秦岭中段南坡油松林具有相对较高的固碳能力。
3.2 地形因子对油松林生态系统碳密度各组分的影响
油松林生态系统中乔木层与土壤层碳密度之和占到总碳密度的97.52%,因此,地形因子对油松林生态系统总碳密度的影响主要通过影响乔木层和土壤层碳密度来实现。
本研究结果表明,不同海拔范围的油松林生态系统总碳密度无显著差异,海拔对油松林生态系统碳密度的影响主要体现在乔木层,对其他各层的影响则是不显著的。海拔高度的变化会引起水热状况的差异。随着海拔的增加,气温逐渐降低,而根据董立民[18]等对秦岭火地塘林区的研究,年降水量随海拔呈现先增后减的规律,年降雨量最大值出现在2000 m。陕西森林编辑委员会[19]和李战刚[20]的资料表明在海拔800—1000 m,年均温为13—14 ℃,≥10℃年积温4000—4500 ℃;海拔1200—1500年均温6—10 ℃,≥10 ℃年积温1700—2500 ℃;而海拔2000 m以上年均温在6 ℃以下,≥10 ℃年积温不足1700 ℃。油松喜温凉气候,耐寒能力较强,因此油松林乔木层的碳密度在海拔1500 m以下随海拔降低而减小的可能原因为低海拔的降水量减少而温度上升,油松生长情况不如中海拔范围,这也是油松在本区域分布下界被马尾松代替的原因;而高海地区由于气温较低,年积温较少,油松的生长较之中海拔范围也较差。根据陈存根[21]等的研究油松在火地塘林区适生海拔为1560—1640 m,这与本研究油松林乔木层碳密度在海拔1500—1700 m范围内达到最大相一致。
在该地区,坡度对油松林生态系统碳密度的影响在乔木层、土壤层、枯落物层均达到显著或极显著水平。这使得该地区不同坡度的油松林生态系统总碳密度呈现极显著差异。本研究结果显示,斜坡和陡坡油松林生态系统乔木层碳密度显著高于其他坡度范围。阳昌平等[22]在太岳山对不同坡度油松生长状况的研究表明,缓坡与斜坡的树高、胸径均高于平地和陡坡,本研究的结果与之相似。坡度主要影响林冠层所接受的光照以及土壤水肥状况。事实上坡度可以增加林冠层接受光照的面积,这有利于乔木的生长和吸收固定CO2。但是在坡面上,随着坡度的增加,降雨入渗到土壤的量减少,而多以地表径流的方式流失[23]。由于重力作用,土壤水分难以保持;同时坡度的增大还带来土壤养分的流失加剧,随着坡度增大,土壤养分减少。土壤水肥状况变差使得较大坡度上油松生长较差,碳密度较低。而本研究与阳昌平等在太岳山的研究具体最适坡度的不同一个可能的原因是太岳山年降水低于秦岭,坡度因素中水分的影响较大,而秦岭南坡中段由于水分条件较好,光照的影响较太岳山显著。在本研究中,油松林生态系统土壤层、枯落物层的碳密度随坡度的增大而减小,达到极显著水平。在较大坡度的油松林中,枯落物由于重力,地表径流以及风等作用向下运动而造成损失,难以在林内积累并向土壤中输入有机质。土壤有机质因为水土流失和输入减少而显著降低。最终由于坡度对乔木、土壤以及枯落物层的显著影响,在该地区,油松林生态系统碳密度随坡度的增大而显著减少。
坡位对油松林生态系统碳密度的影响主要体现在乔木层与土壤层。本研究的结果类似于刘艳等[24]在北京百花山和侯琳等[25]在火地塘的研究,土壤碳密度在下坡位显著高于上坡位,而中坡位土壤碳密度介于上坡位和下坡位,但与两者差异均不显著。其可能的原因除了土壤颗粒以及枯落物向下运移(事实上在本研究中枯落物层的碳密度在不同坡位也表现为下坡位(2.98 t/hm2)>中坡位(2.78 t/hm2)>上坡位(2.23 t/hm2)趋势),还与上坡位的土层较薄有关。乔木层的碳密度中坡位显著高于上坡位和下坡位。土壤养分和水分的向下运移以及上坡位较浅的土层使得中坡位较上坡位更适合油松的生长。而下坡位在较优的水肥条件下其乔木层碳密度并不高,其可能原因之一为下坡位人为干扰比较强烈,严重影响了油松林碳的积累。这也是在本研究中油松林生态系统碳密度在低海拔、小坡度、下坡位地段碳密度较小的重要原因之一。
油松林乔木层碳密度在不同坡向表现为阳坡显著大于阴坡。在秦岭南坡中段水热条件较好的情况下,光照条件是影响油松生长的主要因素。油松是喜光树种,阴坡光照条件相对较差,不利于油松的生长和碳密度的积累。林下植被的生长受到水分、温度、养分、光照等多种因素的综合影响。阳坡油松生长优于阴坡,郁闭度较高,林下植被获得的光照较少,可能是导致林下灌木层、草本层碳密度阴坡大于阳坡的主要因素。阴坡地表温度较低,不利于枯落物的分解,导致枯落物层碳密度阴坡大于阳坡,但是这些差异并不显著。
4 结论
秦岭中段南坡油松林生态系统中地形因子对油松林生态系统碳密度的影响主要表现在乔木层和土壤层。海拔、坡度、坡位、坡向均对乔木层的碳密度影响显著(P<0.05),而坡度和坡位对土壤层碳密度具有显著的影响(P<0.05)。森林生态系统的碳密度在不同立地条件下的差异,最终是受光照、温度、水分等环境因子的综合影响,同时也与森林植被的特征有关。在不同地区差异较大的气候条件下或不同的森林类型下,森林生态系统各碳库在不同立地条件下的分布可能有所差异。这使得森林生态系统碳循环的研究以及利用森林调查资料进行大尺度碳储量估算时,应给与地形因子和气候条件与植被类型同等的重视,以保证研究或估算的可靠性。
[1] 水利部应对气候变化研究中心. 气候变化权威报告——IPCC报告. 中国水利, 2008, (2): 38-40.
[2] Malhi Y, Baldocchi D D, Jarvis P G. The carbon balance of tropical, temperate and boreal forests. Plant, Cell and Environment, 1999, 22(6): 715-740.
[3] 胡会峰, 刘国华. 森林管理在全球CO2减排中的作用. 应用生态学报, 2006, 17(4): 709-714.
[4] 刘华, 雷瑞德. 秦岭火地塘林区主要森林类型碳储量和碳密度估算. 中国农学通报, 2005, 21(3): 138-142.
[5] 马明, 王得祥, 刘玉民. 秦岭天然华山松林碳素空间分布规律及其动态变化. 林业资源管理, 2008, (5): 75-78.
[6] 侯琳, 雷瑞德, 王德祥, 尚廉斌, 赵辉. 秦岭火地塘林区油松群落乔木层的碳密度. 东北林业大学学报, 2009, 37(1): 23-24.
[7] 马明, 王得祥, 刘玉民. 秦岭火地塘林区天然油松林碳素空间分布规律. 西南大学学报: 自然科学版, 2009, 31(3): 114-118.
[8] 邓蕾, 上官周平. 秦岭宁陕县森林植被碳储量与碳密度特征. 西北植物学报, 2011, 31(11): 2310-2320.
[9] 康永祥, 张景群, 李登武. 秦岭中段北坡主要植被类型潜在火行为划分. 东北林业大学学报, 1999, 27(5): 20-24.
[10] 罗伟祥. 刘广全, 李家珏. 西北主要树种培育技术. 北京: 中国林业出版社, 2007: 238-245.
[11] 李海奎, 雷渊才, 曾伟生. 基于森林清查资料的中国森林植被碳储量. 林业科学, 2011, 47(7): 7-12.
[12] 赵敏, 周广胜. 中国森林生态系统的植物碳贮量及其影响因子分析. 地理科学, 2004, 24(1): 50-54.
[13] 徐新良, 曹明奎, 李克让. 中国森林生态系统植被碳储量时空动态变化研究. 地理科学进展, 2007, 26(6): 1-10.
[14] 方精云, 郭兆迪, 朴世龙, 陈安平. 1981—2000年中国陆地植被碳汇的估算. 中国科学D辑, 2007, 37(6): 804-812.
[15] Dixon R K, Solomon A M, Brown S, Houghton R A, Trexier M C, Wisniewski J. Carbon pools and flux of global forest ecosystems. Science, 1994, 263(5144): 185-190.
[16] 程堂仁, 冯菁, 马钦彦, 王玉涛, 康峰峰, 冯仲科, 张彦林, 邓向瑞. 甘肃小陇山森林植被碳库及其分配特征. 生态学报, 2008, 28(1): 33-44.
[17] 任军辉, 刘建军, 刘斌, 张宇钰, 杜承星. 宁夏贺兰山天然油松林碳储量和碳密度. 东北林业大学学报, 2011, 39(5): 108-110.
[18] 董立民, 刘淑明, 辛继红. 秦岭火地塘林场气候要素随海拔高度的变化. 水土保持通报, 1995, 15(3): 16-19.
[19] 陕西森林编辑委员会. 陕西森林. 北京: 中国林业出版社, 1986: 1-31.
[20] 李战刚, 党坤良, 李登武. 陕西天华山自然保护区综合科学考察与研究. 西安: 陕西科学技术出版社, 2005: 1-29.
[21] 陈存根, 彭鸿. 秦岭火地塘林区主要森林类型的现存量和生产力. 西北林学院学报, 1996, 11(Supp l): 92-102.
[22] 阳昌平, 林鸿兴, 骈铁虎. 太岳山油松生长与主要地形林分因子关联分析. 山西林业科技, 1988, (3): 8-13.
[23] 郝振纯, 吕美霞, 吕美朝, 王加虎. 坡度作用下土壤水分时空异质性研究. 水文, 2012, 32(2): 5-10.
[24] 刘艳, 查同刚, 付汝军. 百花山典型林分土壤有机碳储量及垂直分布特征. 西北农业学报, 2012, 21(3): 182-187.
[25] 侯琳, 雷瑞德, 王得祥, 赵辉. 秦岭火地塘天然次生油松林土壤有机碳的特征. 西北农林科技大学学报: 自然科学版, 2008, 36(8): 156-160.
Organic carbon density inPinustabulaeformisforest ecosystem on the south slope of the middle Qinling Mountains, China
SHEN Biao, DANG Kunliang*, WU Penghui, ZHU Chenggong
CollegeofForestry,NorthwestAgriculture&ForestUniversity,Yangling712100,China
Organic carbon density (OCD) in different carbon pools ofPinustabulaeformisforest ecosystem and its relationship with different geocentric factors are important for estimating forest carbon budgets, and helpful to understand forest carbon cycles.In this study, we established fifty sample plots at different site conditions using typical sampling method on the south slope of the Middle Qinling Mountains, wherePinustabulaeformisis widely distributed. Organic carbon density in different carbon pools ofPinustabulaeformisforest ecosystem and its relationship with different geocentric factors were estimated and analyzed. Our objective was to estimate OCD in different carbon pools ofPinustabulaeformisforest ecosystem and its distribution on different geocentric factors, to understand the effects of geocentric factors on forest carbon budgets. The results showed that the total OCD ofPinustabulaeformisforest ecosystem was 150.12 t/hm2in this region. The proportion of soil pool OCD was 56.74% and comprised the major part of the OCD of ecosystem. The OCD of litter pool was 2.66 t/hm2, accounting for 1.77% of the ecosystem. Carbon density of vegetation pool was 62.29 t/hm2,taking up 41.49% of the OCD of ecosystem. The OCD of vegetation layer in this region was much higher than the average vegetation pool OCD of forest ecosystem in China, and mere carbon sequestrations are expected. The tree OCD was 61.22 t/hm2, accounting 98.3% of the whole vegetation OCD. The OCD of shrub and herb were 0.65 t/hm2and 0.41 t/hm2, accounting for 1.04% and 0.66% of the vegetation pool respectively. The OCD of different compositions of tree ranked as stem>branch>roots>leaf>bark. Effects of geocentric factors on the OCD of shrub and herb were insignificant. With the increase of elevation, the OCD of tree first increased and then decreased, peaking at 1500—1700 (a.s.l.). Variations of OCD in litter, soil and the ecosystem were insignificant. With the increase of slope gradient, the OCD of litter, soil and the ecosystem significantly decreased. In comparison, the OCD of tree first increased and then decreased, reaching its maximum on the slope gradient of 26—35°. The OCD of soil was higher in the lower slope compared with that of the middle and the upper slope, while the OCD of tree and the ecosystem was larger in the middle slope. Nevertheless, effects of slope position on the OCD of litter were slight. The OCD of tree at sun-facing slope appeared higher than that of back slope. While effects of slope aspect on the OCD of litter, soil layer and ecosystem were insignificant. It was concluded that effects of geocentric factors on the OCD ofPinustabulaeformisforest ecosystem were mainly on the tree pool and soil pool on the south slope of the Middle Qinling Mountains. Effects of elevation, slope gradient, slope position, slope aspect were significant on tree pool, and effects of slope gradient, slope position were significant on soil pool. Geocentric factors should be considered in researches on forest ecosystem carbon cycles and estimation of carbon storage of forest ecosystem.
organic carbon density;Pinustabulaeformis;ecosystem; the south slope of the Qinling Mountains
林业公益性行业科研专项项目(201204502)
2013-05-09;
日期:2014-07-18
10.5846/stxb201305090996
*通讯作者Corresponding author.E-mail: Dangkl@126.com
沈彪,党坤良,武朋辉,朱成功.秦岭中段南坡油松林生态系统碳密度.生态学报,2015,35(6):1798-1806.
Shen B, Dang K L, Wu P H, Zhu C G.Organic carbon density inPinustabulaeformisforest ecosystem on the south slope of the middle Qinling Mountains, China.Acta Ecologica Sinica,2015,35(6):1798-1806.
猜你喜欢
天天爱科学(2022年4期)2022-05-23
当代陕西(2022年5期)2022-04-19
水资源开发与管理(2019年11期)2019-12-02
世界家苑(2018年9期)2018-09-18
小哥白尼(野生动物)(2018年4期)2018-08-01
现代农村科技(2018年3期)2018-01-18
山东林业科技(2017年1期)2017-06-29
河北林业科技(2016年5期)2016-11-08
河北林业科技(2016年3期)2016-10-27
河北林业科技(2015年3期)2015-12-17
