
胡杨枝芽生长特征及其展叶物候特征
2015-03-10 12:19:18郑亚琼李志军
生态学报 2015年4期
郑亚琼,冯 梅,李志军,*
1 新疆生产建设兵团塔里木盆地生物资源保护利用重点实验室,阿拉尔 843300 2 塔里木大学植物科学学院,阿拉尔 843300
胡杨枝芽生长特征及其展叶物候特征
郑亚琼1,2,冯 梅1,2,李志军1,2,*
1 新疆生产建设兵团塔里木盆地生物资源保护利用重点实验室,阿拉尔 843300 2 塔里木大学植物科学学院,阿拉尔 843300
以5个不同发育阶段的胡杨(PopuluseuphraticaOliv.)个体为研究对象,观测记录了枝芽展叶物候、枝芽生长特征和叶形变化的空间分布规律。结果表明:不同发育阶段的胡杨个体以及同一个体树冠的不同层次,其枝芽生长及其展叶物候期表现出不同的时空特征。随着树龄的增加和树冠层次的增高(由基向顶),当年新生枝条长度、枝条叶片数和叶形指数逐渐减小,但叶面积和叶片干重逐渐增大。5个不同发育阶段胡杨个体均表现出展叶物候始于树冠顶层,依次向下结束于树冠基部;展叶物候期共性表现在枝芽萌动期均在4月上旬,起始展叶期集中在4月中旬,展叶终期则在5月上旬到下旬;树龄较大的个体其枝芽萌动期、起始展叶期、展叶终期较树龄较小的个体早;其枝芽萌动期到展叶终期的时间进程较树龄较小的个体短;不同发育阶段的个体枝芽萌动期出现的时间较为离散,起始展叶期和展叶终期出现的时间较为集中。相关分析表明,出叶周期与枝条长度、枝条叶片数量和叶形指数呈极显著正相关,与叶面积和叶片干重呈显著负相关。
胡杨; 枝芽萌动和生长; 展叶物候; 叶形指数; 叶面积; 叶干重
芽是尚未发育的枝、花或花序,是枝、花或花序的原始体。依照芽发育成的器官来划分,芽可以分为枝芽(叶芽)、花芽和混合芽。关于枝芽的研究主要集中在同一树种或同一树种不同品种枝芽形态特征、生长特性和发育进程[1- 4]等方面,以期完善该树种发育生物学的基础理论研究,也为树木良种选育和栽培技术研究提供理论依据。
植物物候是指植物受生物因子和非生物因子如气候、水文、土壤等影响而出现的以年为周期的自然现象,它包括植物的发芽、展叶、开花、叶变色、落叶等现象,是植物长期适应每年季节性变化的环境而形成的生长发育节律[5]。各种植物的物候期均按一定的先后次序出现,前一个物候期来临的迟早与继后出现的物候现象的早迟有密切关系,并且各年各物候期的出现都有其规律性[6]。不同植物的物候期在同样的环境条件下反应各异[7- 8],同种植物在相同环境中物候期相同,这都是由其自身的生物学特性(即遗传特性)所决定的[9]。然而,同一种植物不同的品系,如杨属植物不同的杨树品种、无性系,即使在相同环境中各物候期也会存在很大差异[10- 14]。关于气候变化对植物物候的影响,大量的研究工作集中在开花物候方面。近几十年来,研究者对植物展叶时间的研究兴趣日益增加[15- 16],植物的展叶时间与气温紧密相关,而对于另一些植物而言, 光周期也很重要[17]。
胡杨(P.euphratica)是隶属于杨柳科(Salicaceae)胡杨亚属的木本植物,是最古老、最原始的荒漠河岸林树种,主要分布在中亚、西亚和地中海地区[18]。全世界最大面积的天然胡杨林集中分布在哈萨克斯坦和中国,中国91.1%的胡杨分布在新疆,其中的89.1%集中分布于南疆塔里木盆地[19]。胡杨被观察到具有异形叶性,叶形多变化表现在幼苗、幼树和成年树下部萌生条上叶呈线状披针形,随着树体的生长,成年树上不仅存在披针形叶,而且出现了卵状菱形、锯齿卵圆形或肾形叶,形成了上层树冠叶片卵圆形、树冠下部叶片仍为披针形的独特外观,因此胡杨又被称为异叶杨或变叶杨[20]。有研究进一步表明,不同发育阶段的胡杨个体上出现不同形状的异形叶源于树冠不同垂直空间枝芽的生长;在同一个生长季内,总是树冠最顶端的阔卵形叶先展开,下层的卵形、披针形和条形叶逐渐依次展开[21],表明树冠不同垂直空间枝芽的生长特征、叶形变化及展叶物候有所不同,相互间可能存在一定的内在联系。目前,关于胡杨物候特征研究仅见开花物候[20,22- 24]方面的相关报道,但尚未涉及不同发育阶段个体、以及同一个体树冠不同垂直空间枝芽生长特征及其展叶物候的研究。本文选择不同发育阶段的胡杨个体为研究对象,基于不同发育阶段胡杨个体的叶形变化,研究不同发育阶段胡杨个体以及同一个体树冠不同垂直空间枝芽生长特征、展叶物候间的差异,阐明枝芽生长特征、展叶物候与叶形变化间的关系,为进一步揭示叶形变化在胡杨生活史策略中的意义奠定基础。
1 研究方法
1.1 研究区概况
研究区位于塔里木盆地西北缘(81°17′56.52″ E,40°32′36.90″ N)新疆阿拉尔市塔里木大学校园外的人工胡杨林。该区气候炎热干燥,终年干旱少雨,年降雨量仅约50 mm,潜在蒸发量约1900 mm,年均气温10.8 ℃,年均日照时数为2900 h,是典型的温带荒漠气候。研究区人工胡杨林面积是1.72×105m2,株行距为1.20 m×4.20 m。人工林内胡杨平均胸径最小的为2.44 cm,最大的为17.4 cm,平均树龄最小的为4.13a,最大的为11.33a,平均树高6.41 m,不同树龄的个体总体上覆盖了异形叶变化的各种要类型。人工林每年春、冬灌水各1次,夏季灌水2—3 次,锈病防治1次。
1.2 观测样株和观测枝条的选定方法
有研究表明,幼龄林胡杨胸径与树龄之间呈现显著正相关,相关系数均达到0.96以上[25]。据此,本研究以5个不同发育阶段的胡杨个体(以A、B、C、D、E分别代表5个不同发育阶段的个体类型)为研究对象,每一个发育阶段的个体各选取3棵做为重复观测样株。5个不同发育阶段胡杨个体胸径、树高及树冠上自然呈现的异形叶种类情况见图1。
A型个体 仅有条形叶1种叶形,平均树龄为3a,平均胸径为2.3 cm,平均树高为4.13 m。
B型个体 具有条形叶和披针形叶2种叶形,平均树龄为5a,平均胸径为4.0 cm,平均树高为4.59 m。
C型个体 具有条形叶、披针形叶和卵形叶3种叶形,平均树龄为7a,平均胸径为5.1 cm,平均树高为5.11 m。
D型个体 具有条形叶、披针形叶、卵形叶和阔卵形叶4种叶形,平均树龄为8a,平均胸径为8.2 cm,平均树高为7.19 m。
E型个体 具有披针形叶、卵形叶和阔卵形叶3种叶形,平均树龄为10a,平均胸径为10.6 cm,平均树高为8.45 m。

图1 不同发育阶段胡杨个体异形叶的类型及空间分布格局模式图Fig.1 Populus euphratica Oliv. heteromorphic leaf ontogenetic process type and pattern of spatial distribution pattern1: 条形叶、2: 披针形叶、3: 卵形叶、4: 阔卵形叶; A—E代表个体形
在选定样株的基础上,以全站仪测量每棵样株树冠最高点及树冠最低点的高度,二者相减后得出树冠的垂直冠幅,将冠幅划分为5等分(5个层次),从下往上依次为1到5层,在每一层次的中央位置,按东、南、西、北方位选取4个定期观测枝条(作为每个层次上的重复),挂牌标记。
1.3 展叶物候观测方法
参照《中国物候观测方法》[26]和木本植物观测标准[27]逐日或隔日进行观测,观测时借助6 m铝梯到达树冠中部,结合目测与望远镜进行观测,记录不同发育阶段胡杨个体枝芽萌动期(芽的鳞片开始分离,露出枝芽)、起始展叶期(新枝基部第一片叶由卷曲到展平)、展叶终期(观测枝条上所有叶片完全展平),统计叶数量持续时间(即出叶周期,是指植物从爆芽开始到叶数量达到稳定的这段时间)[28]。
1.4 枝芽生长特征调查
从爆芽开始到新生茎叶面积、叶数量达到稳定的这段时间,将定期观测的枝条采下,调查测定5个不同发育阶段个体树冠各层次枝芽萌动后生长形成的新生茎长度、茎上叶片数,并摘取茎上所有叶片,以MRS- 9600TFU型号扫描仪获取叶片宽度、长度和叶面积参数。
1.5 数据处理
将所得数据进行整理后,用dps软件进行数据相关分析。统计处理前,先将表中的日期型数据变换成数字型数据。按减去常数项的办法[29]转换数据,计算出变动系数[30]。数据转换计算公式为:
X″ij=Xij- minjXij
式中,X″ij为转换后第i行第j列的数据,Xij为转换前i行第j列的数据,minjXij为转换前第j列的最小值。
2 结果与分析
2.1 不同发育阶段胡杨个体枝芽生长特征
2.1.1 不同发育阶段胡杨个体枝条长度的变化规律
图2显示,5个不同发育阶段的胡杨个体表现出随着树龄的增加其枝条长度逐渐变短;方差分析结果进一步表明,A型(仅有条形叶个体)和B型(条形叶与披针形叶共存个体)发育阶段个体间枝条长度没有差异,但A型、B型均与C型(具有条形叶、披针形叶和卵形叶3种叶形个体)、D型(具有条形叶、披针形叶、卵形叶和阔卵形叶4种叶形个体)、E型(具有披针形叶、卵形叶和阔卵形叶3种叶形个体)发育阶段的个体枝条长度有显著差异,C型、D型、E型3个发育阶段个体间则呈现C型、E型枝条长度有差异显著,D型表现为过渡状态。对同一发育阶段个体树冠不同层次的枝条长度进行比较发现(图2),随着冠层的增高枝条长度逐渐变短;方差分析结果显示,树龄较小的个体(A型、B型)不同层次之间枝条长度差异显著,树龄较大的个体(C、D)不同层次之间枝条长度只有第1层和第5层间有显著差异,中间层次之间差异不明显,特别是E型个体不同层次间的枝条长度没有差异。说明枝条长度变化最大的阶段在胡杨个体发育的早期阶段及其树冠的不同层次上;随着树龄的增加,枝条长度在个体的整体水平上趋于变短的同时,在树冠的不同层次上则趋于稳定。

图2 不同发育阶段胡杨个体和同一个体树冠不同层次枝条长度的变化规律Fig.2 Variation in branch length of different development stages and tree levels
2.1.2 不同发育阶段胡杨个体枝条叶片数量的变化规律
从图3可以看出,5个不同发育阶段个体叶片数/枝有所不同,表现出随着树龄增加个体叶片数/枝逐渐减少的规律;方差分析表明,B型与C型、D型与E型个体间叶片数/枝无差异,但B型与C型、D型与E型同A型间均差异显著。同一个体不同冠层之间的叶片数/枝有所不同,随着冠层的增高叶片数/枝呈现出减少的规律,但方差分析结果显示,A型个体不同冠层之间的叶片数/枝差异不显著,B、C、D、E型个体均表现出树冠第1层和第5层间差异显著。
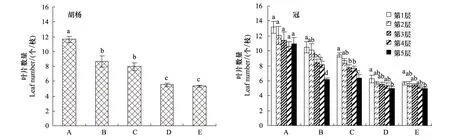
图3 不同发育阶段胡杨个体和同一个体树冠不同层次枝条叶片数量的变化规律Fig.3 Variation in leaf number of different development stages and tree levels

图4 不同发育阶段胡杨个体和同一个体树冠不同层次叶形指数的变化规律Fig.4 Variation in leaf shape index of different development stages and tree levels
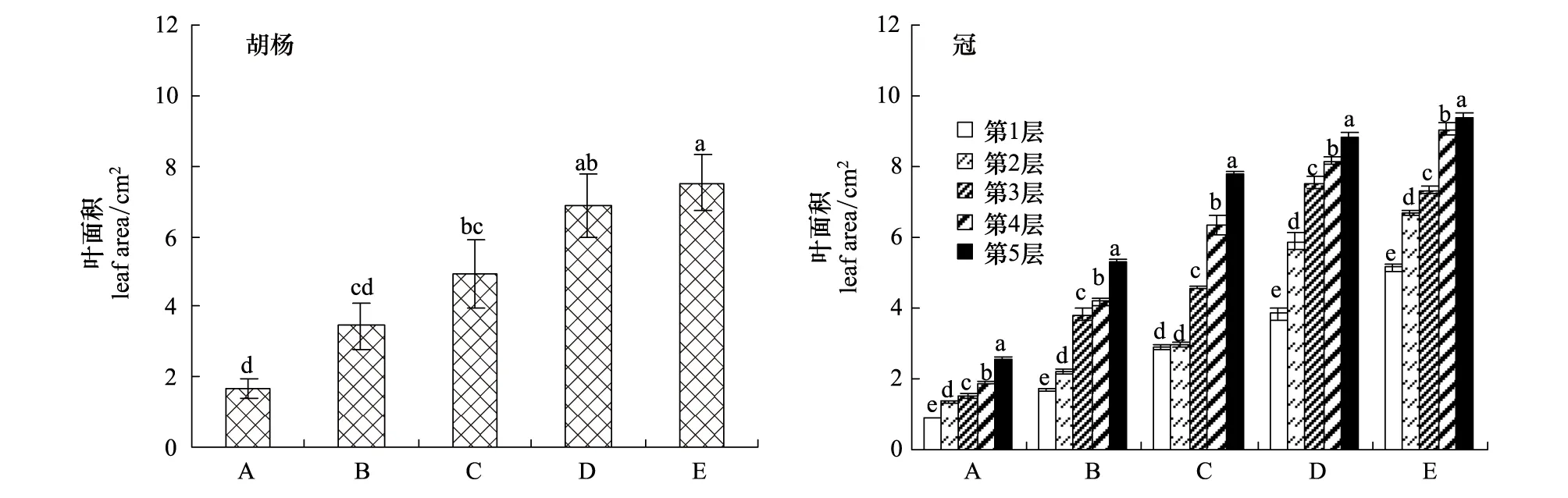
图5 不同发育阶段胡杨个体和同一个体树冠不同层次叶面积的变化规律Fig.5 Variation in leaf area of different development stages and tree levels
2.1.3 不同发育阶段胡杨个体叶形指数的变化规律
叶形指数(叶片长宽比)反映的是植物叶片形状的变化。图4表明,不同发育阶段的个体叶片叶形指数随着树龄的增加逐渐减小;方差分析结果显示,A、B型个体间差异不显著,到C型阶段差异显著,而C、D、E型个体之间差异不显著。同一个体树冠不同层次叶片叶形指数呈现出由树冠基部向顶部的方向逐渐减小的规律,A、B、C、D、E型个体均表现出树冠第1层、第2层、第3层间差异显著,第1层和第5层间差异显著,第4层和第5层间叶形指数无差异(图4)。
2.1.4 不同发育阶段胡杨个体枝条叶面积的变化规律
从图5可以看出,枝条叶面积随着树龄的增加而增加,不同发育阶段个体间枝条叶面积差异显著。同一个体树冠不同层次枝条叶面积也有不同,并呈现出由树冠基部向顶部的方向逐渐增大的规律;A、B、C、D、E型个体均表现出同一个体树冠不同冠层之间的枝条叶面积差异显著(图5)。
2.1.5 不同发育阶段胡杨个体枝条叶片干重的变化规律
从图6可以看出,枝条叶片干重随着树龄的增加而增加,不同发育阶段个体间枝条叶片干重差异显著。同一个体树冠不同层次枝条叶片干重差异不显著,但呈现出由树冠基部向顶部的方向逐渐增大的规律(图6)。
从图2—图6可以看出,叶片数量和叶形指数随着枝条长度的增长逐渐增加,但随着枝条长度的增长叶面积和叶片干重逐渐变小,表明胡杨个体上枝条长度越长,枝条上叶片数量越多,其叶面积和叶片干重越小,叶形指数越大。
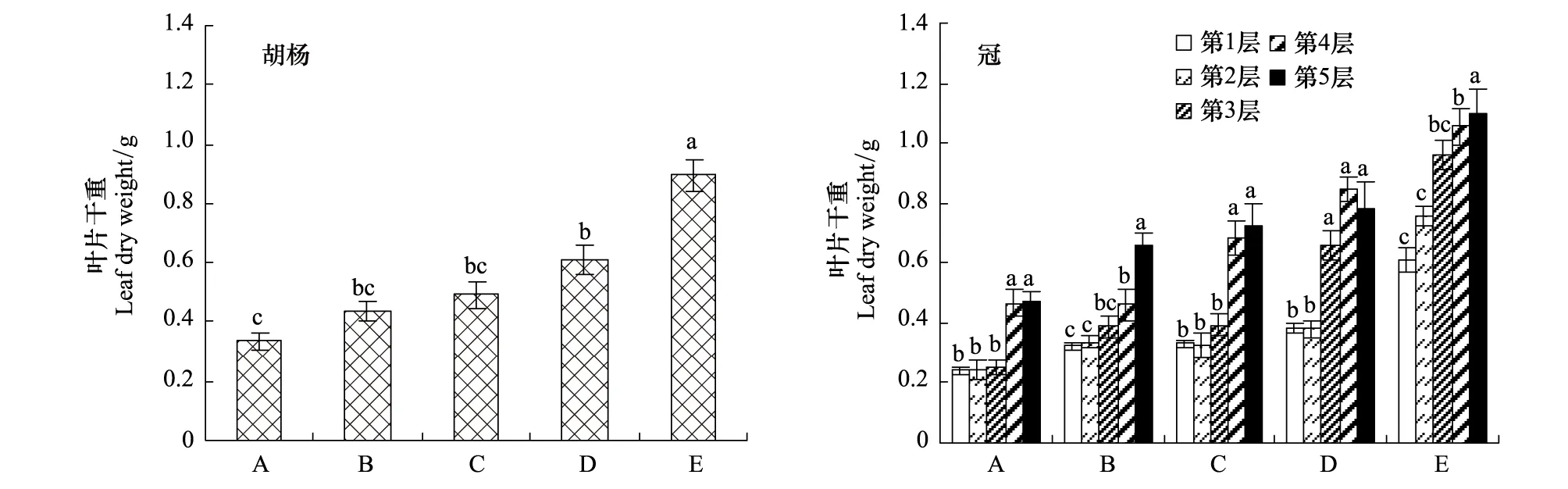
图6 不同发育阶段胡杨个体和同一个体树冠不同层次叶片干重的变化规律Fig.6 Variation in leaf dry weight of different development stages and tree levels柱状图上方字母不同表示两者间差异在0.05水平上达到显著,误差棒代表一个标准偏差
2.2 不同发育阶段胡杨个体展叶物候的时空特征
2.2.1 不同发育阶段胡杨个体展叶物候的空间格局
表1—表3显示,5个不同发育阶段胡杨个体均表现出展叶物候始于树冠顶层枝芽的萌动、展叶,依次向下,结束于树冠下层基部枝芽的萌动、展叶,表明胡杨个体展叶物候期具有空间秩序的特征。
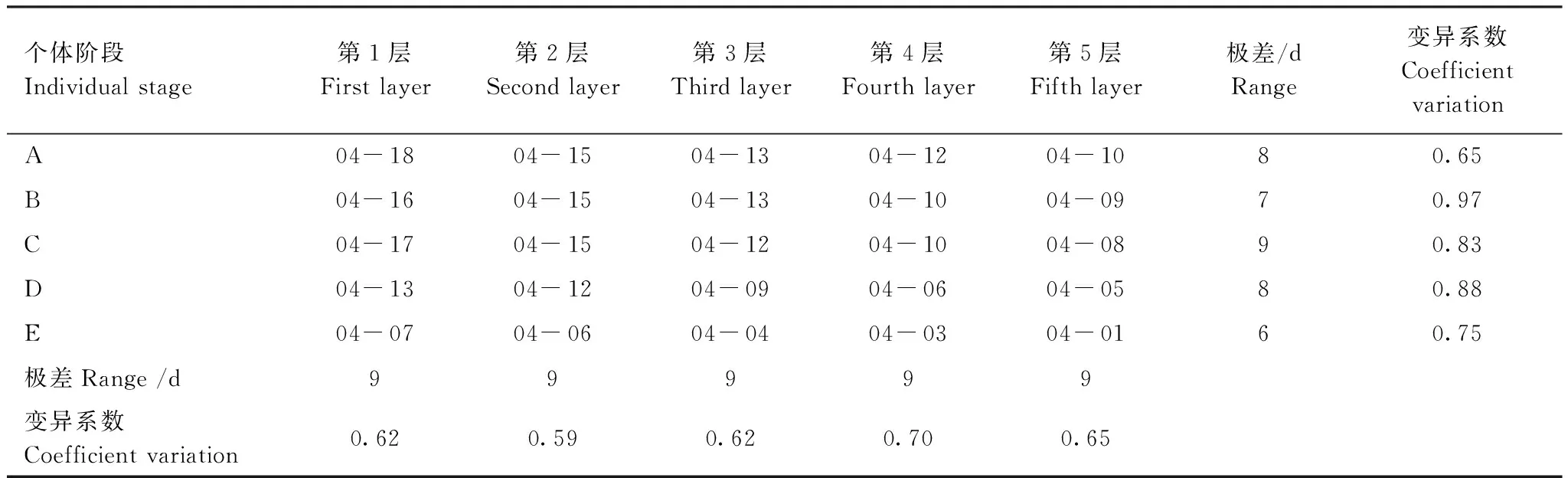
表1 不同发育阶段胡杨个体树冠不同层次枝芽萌动期物候Table 1 The leaf phenology in germinating branch bud of different development stages Populus euphratica Oliv different layers
A型(仅有条形叶个体);B型(条形叶与披针形叶共存个体);C型(具有条形叶、披针形叶和卵形叶3种叶形个体);D型(具有条形叶、披针形叶、卵形叶和阔卵形叶4种叶形个体);E型(具有披针形叶、卵形叶和阔卵形叶3种叶形个体)
2.2.2 不同发育阶段胡杨个体物展叶物候的时间特征
从表1—表3可看出,5个不同发育阶段的胡杨个体及同一个体树冠不同层次枝芽萌动、起始展叶及展叶结束的时间均有差异。5个不同发育阶段胡杨植株展叶物候的共性表现在枝芽萌动期均在4月上旬,起始展叶期集中在4月中下旬,展叶终期则在5月上旬到中下旬;差异表现为树龄较大的植株个体其枝芽萌动期、起始展叶期、展叶终期较树龄较小的个体早(E>D>C>B>A);其枝芽萌动期到展叶终期的时间进程较树龄较小的个体短(E﹤D﹤C﹤B﹤A),依次为29、33、35、38、42d。

表2 不同发育阶段胡杨个体树冠不同层次枝芽起始展叶物候Table 2 The leaf initiation period branch bud phenology different development stages of P. euphratica different layers

表3 不同发育阶段胡杨个体树冠不同层次枝芽展叶终期物候Table 3 The leaf eventually period branch bud phenology different development stages of P. euphratica different layers
变异系数能够反映不同发育阶段个体、以及同一个体树冠不同层次某一物候期的离散程度。从表2—表4可见,不同发育阶段个体的变异系数以及同一发育阶段个体不同冠层的变异系数均有差异,变异系数越大,说明此物侯期在此阶段出现的时间越离散。在同一立地条件下,不同发育阶段的胡杨个体,影响其枝芽从萌动到枝条上叶片数不再增加这一生长期长短的因素主要是枝芽萌动期、枝芽起始展叶、枝芽展叶终期的差异。
2.2.3 不同发育阶段胡杨个体出叶周期
胡杨枝芽活动过程是一个从枝芽萌动开始到枝条长度、叶面积和叶数量不断增加直至恒定的过程。从图7可以看出,不同发育阶段的个体植株比较,树龄小的个体出叶周期较长,特别是幼龄阶段的个体完全展叶需要较长的时间,随着树龄的增加个体完全展叶需要的时间逐渐减少,方差分析结果显示,不同发育阶段胡杨的出叶周期差异显著;同一发育阶段的个体不同层次比较(图7),表现为出叶周期随树冠层次的增加呈递减趋势,方差分析结果显示同一发育阶段的个体不同冠层的出叶周期差异不显著。同时发现,叶片的展叶结束时间与展叶起始时间成正比,展叶起始越早,结束就越早。
2.4 出叶周期与枝条长度、叶片数量、叶形指数、叶面积和叶片干重的相关性分析
表4显示,出叶周期与枝条长度、叶片数量和叶形指数呈极显著正相关,但与叶面积和叶片干重呈极显著负相关,说明枝条长度、叶片数量和叶形指数越大其出叶周期越长;叶面积和叶片干重越大其出叶周期越短,表明枝条长度、叶片数量、叶形指数、叶面积和叶片干重均影响不同发育阶段胡杨个体出叶周期的长短。
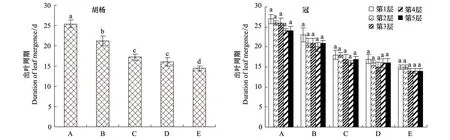
图7 不同发育阶段胡杨个体平均出叶周期及出叶周期随个体树冠层次的变化规律Fig.7 Individual average leaf emergence of different development stages and the change rule in different tree levels

表4 出叶周期与枝条长度、叶片数量、叶形指数、叶面积和叶片干重的相关性Table 4 The correlation between leaf emergence, length of branches, leaf umber, leaf shape index, leaf area and leaf dry weight
*P<0.05,**P<0.01
3 结论与讨论
3.1 胡杨不同生长发育阶段枝芽的生长特征
G. C. S. Negi[31]对生长在喜马拉雅地区600 m和2200 m海拔之间的10个常绿和15个落叶树种进行新梢生长的比较研究,指出常绿树种新梢上的叶子比落叶树种生长的快,落叶树种与常绿树种每枝上叶面积比值是587.7 cm2∶246.7 cm2,枝条延伸生长是19.3 cm∶11.2 cm,这两个群体的差异是与枝条生长特性相关的。陈波等[32]对栲树(Castanopsisfargesii)不同生长发育阶段枝系特征的研究结果表明,栲树在不同发育阶段的总体分枝率和逐步分枝率有显著变化,幼苗和幼树阶段的分枝率较低,而成株阶段的分枝率较高;幼树阶段的枝条长度、枝倾角和叶倾角明显大于幼苗和成株阶段,表现为明显的高生长对策;叶片配置在不同枝系上有较大差异,叶片主要集中于植冠内一级枝和二级枝上;叶片的大小从幼苗、幼树到成株阶段逐渐增大。研究结果显示,在胡杨的枝条系统中,叶分布在当年新生茎上,是枝芽生长的结果。当年新生枝条长度、枝条叶片数、叶形指数、叶面积和叶片干重在不同生长期阶段的个体以及同一个体树冠不同层次上有较大差异,表明枝芽生长特性具有随个体生长发育的不同阶段而变化的特征。随着树龄的增加,当年新生枝条长度、枝条叶片数、叶形指数在个体的整体水平上逐渐减少/小,在树冠由基向顶的层次上也逐渐减少/小,最终趋于稳定;而叶面积和叶片干重则随着树龄的增加而逐渐增加。特别是,随着树龄增加叶片叶形指数减小,不同生长发育阶段的胡杨个体、以及同一个体树冠的不同层次呈现出条形、披针形、卵形、阔卵形叶规律性出现的异形叶性。这一研究结果不同于前人叶性状在冠层不同高度的差异与树木大小有关而与树木年龄无关[33- 34]的研究结论。胡杨这一独特的属性,使其在枝芽生长过程中表现出一定的规律性和与其它物种的差异性。
3.2 胡杨不同生长发育阶段展叶物候及展叶格局
朱旭斌等[28]对南京地区落叶栋林主要木本植物的展叶动态进行了研究,发现叶片大的物种展叶晚,叶片越大,展叶速率越大,但展叶持续时间与叶的大小无关。而Marquis等[35]研究发现从草本层、林下层到林冠层,叶的展叶期逐渐延长,即大叶的物种具有较长的展叶期。对同一立地条件下不同发育阶段胡杨个体枝芽展叶物候的研究表明,胡杨个体展叶物候期具有空间秩序,5个不同发育阶段胡杨个体均表现出展叶物候始于树冠顶层枝芽(树冠顶层枝条叶面积最大)的萌动、展叶,依次向下结束于树冠下层枝芽萌动、展叶的特征;树龄较大(个体平均叶面积较大)的胡杨个体其枝芽萌动期、起始展叶期、展叶结束期较树龄较小(个体平均叶面积较小)的胡杨个体早(E﹥D﹥C﹥B﹥A);其枝芽萌动期到展叶结束期的时间进程较树龄较小(个体平均叶面积较小)的胡杨个体短(E﹤D﹤C﹤B﹤A)。对于胡杨这一物种,由于其枝芽生长特性、叶性状具有随个体生长发育的不同阶段而变化的特征,使其展叶物候、展叶格局也表现出随个体生长发育阶段和空间部位的不同而变化,总体上表现为具有大叶的个体、大叶所在的空间部位枝芽展叶物候较早,具有较短的展叶期。
从物种水平来看,长期的进化和对环境的适应使物种的展叶过程和环境变化相协调,实现了物种对资源利用的最大化,形成了物种特有的展叶格局[36]。在同一立地条件下胡杨的5个发育阶段个体,均表现出展叶物候始于树冠上层枝芽的萌动、展叶,依次向下,结束于树冠下层枝芽萌动、展叶,独特的展叶格局可能是胡杨自身遗传因子作用下在长期的进化过程中形成的独特属性。前期观察表明,越是长/宽比(叶形指数)值小的叶片在植株上出现得越早,即总是树冠最顶端的阔卵形叶先展开,下层的卵形、披针形和条形叶逐渐依次展开[21]。研究证明,不论处在哪个发育阶段,叶形指数最小的叶片(叶面积最大)总是处于树冠的顶层,即也是最先展叶的。叶面积大而且最先占据着树冠的最高空间,非常有利于实现胡杨对光、热资源最大化利用。调查发现,在树冠上有阔卵形叶(是胡杨异形叶中叶面积最大的阔卵形叶开始在树冠顶部出现的部位才伴随着有花芽出现,然后随着个体发育依次向下占据着树冠的绝大部分,花芽的出现表现出与阔卵形叶发生的相同规律,预示着与展叶有相同的开花格局。胡杨是先花后叶植物,胡杨这种由树冠顶部先向基部的展叶格局,至少在进入生殖生长阶段是以便能够尽早尽快为开花结实提供光合产物。
3.3 胡杨不同生长发育阶段出叶周期与枝芽生长特性的关系
胡杨的枝芽生长过程是一个从枝芽开始萌动、枝条生长到其上叶片数不再增加为止的过程,从时间上即为出叶周期。研究分析认为,胡杨枝芽展叶物候与个体发育阶段有关系,但出叶周期更多的是取决于不同发育阶段同一个体树冠不同层次枝芽生长特性,因为胡杨的幼叶蕴藏在上年生枝条顶端的枝芽内。因此,出叶周期必然会随着枝条长度、叶片数量、叶形指数和叶面积变化而变化,而且它们之间表现出极显著的相关性。Sonia Mediavilla和Alfonso Escudero[37]研究了常绿树种和落叶树种幼树和成年树出叶周期的类型和叶片寿命,目的是分析和解释叶性状与树龄的相关性,发现幼树叶片持续时间比成年树低,认为出叶周期和叶寿命在两个生长阶段的差异与单位面积上的叶片质量的资源可用性有关。胡杨个体生长发育过程出叶周期表现出与上述相似的规律,即树龄小的植株个体出叶周期较长,特别是幼龄阶段的植株个体完全展叶需要较长的时间,随着树龄的增加植株个体完全展叶需要的时间逐渐减少。
[1] 侯玉珏, 张晓云, 赵彩平, 韩明玉, 刘小媛, 邢利博. 矮化型、短枝型和柱型苹果苗枝芽特性和叶片特征比较. 西北农业学报, 2012, 21(7): 134- 137.
[2] 彭邵锋, 陈永忠, 马力, 王湘南, 王瑞, 陈隆升, 陆佳. 油茶良种枝梢生长特性研究. 林业科技开发, 2011, 25(2): 24- 28.
[3] 吴昌陆, 陈卫元, 杜庆平. 蜡梅枝芽特性的研究. 园艺学报, 1999, 26(1): 37- 42.
[4] 王丛红, 王秀华. 水曲柳枝芽特性的研究. 吉林林业科技, 2007, 36(5): 7- 9.
[5] 陆佩玲, 于强, 贺庆棠. 植物物候对气候变化的响应. 生态学报, 2006, 26(3): 923- 929.
[6] 王连喜, 陈怀亮, 李琪, 余卫东. 植物物候与气候研究进展. 生态学报, 2010, 30(2): 447- 454.
[7] 白洁, 葛全胜, 戴君虎. 贵阳木本植物物候对气候变化的响应. 地理研究, 2009, 28(6): 1607- 1614.
[8] 常兆丰, 韩福贵, 仲生年. 甘肃民勤荒漠区18种乔木物候与气温变化的关系. 植物生态学报, 2009, 33(2): 311- 319.
[9] 韩小梅, 申双和. 物候模型研究进展. 生态学杂志, 2008, 27(1): 89- 95.
[10] 李守勇, 孙明高, 李学宏, 祝玉. 11个黑杨无性系物候期变异分析. 西北林学院学报, 2003, 18(3): 40- 42.
[11] 杨成生, 王芳, 张亚军, 姜成英. 10个杨树品种的物候期研究. 甘肃林业科技, 2006, 30(2): 29- 31, 48- 48.
[12] 祁如英, 严进瑞, 王启兰. 青海小叶杨物候变化及其对气候变化的响应. 中国农业气象, 2006, 27(1): 41- 45.
[13] 杨丽桃, 侯琼. 内蒙古东部地区小叶杨物候变化与气象条件的关系. 气象与环境学报, 2008, 24(6): 39- 44.
[14] 王德新, 张晏, 段安安, 何承忠. 滇杨优树无性系物候期观测. 西南林学院学报, 2009, 29(6): 21- 23, 27- 27.
[15] Cleland E E, Chuine I, Menzel A, Mooney H A, Schwartz M D. Shifting plant phenology in response to global change. Trends in Ecology and Evolution, 2007, 22(7): 357- 365.
[16] Forrest J, Miller-Rushing A J. Toward a synthetic understanding of the role of phenology in ecology and evolution. Philosophical Transactions of the Royal Society B-Biological Sciences, 2010, 365(1555): 3101- 3112.
[17] Körner C, Basler D. Phenology under global warming. Science, 2010, 327(5972): 1461- 1462.
[18] 王世绩, 陈炳浩, 李护群. 胡杨林. 北京: 中国环境科学出版社, 1995: 3- 18.
[19] 王世绩. 全球胡杨林的现状及保护和恢复对策. 世界林业研究, 1996, 9(6): 37- 43.
[20] 魏庆莒. 胡杨. 中国林业出版社, 1990: 24- 28.
[21] 黄文娟, 李志军, 杨赵平, 白冠章. 胡杨异形叶结构型性状及其相互关系. 生态学报, 2010, 30(17): 4636- 4642.
[22] 李志军, 刘建平, 于军, 周正立. 胡杨、灰叶胡杨生物生态学特性调查. 西北植物学报, 2003, 23(7): 1292- 1296.
[23] 周正立, 李志军, 龚卫江, 高山. 胡杨、灰叶胡杨开花生物学特性研究. 武汉植物学研究, 2005, 23(2): 163- 168.
[24] 买尔燕古丽·阿不都热合曼, 艾里西尔·库尔班, 阿迪力·阿不来提, 热合木都拉·阿迪拉, 段含明, 阿不都米吉提·阿布利克木, 玉米提·哈力克. 塔里木河下游胡杨物候特征观测. 干旱区研究, 2008, 25(4): 525- 530.
[25] 顾亚亚, 张世卿, 李先勇, 李志军. 濒危物种胡杨胸径与树龄关系研究. 塔里木大学学报, 2013, 25(2): 11- 21.
[26] 宛敏渭, 刘秀珍. 中国物候观测方法. 北京: 科学出版社, 1979.
[27] 夏林喜, 牛永波, 李爱萍, 武书龙, 王喜莲. 浅谈木本植物物候观测要求及各物候期观测标准. 山西气象, 2006, (2): 47- 48.
[28] 朱旭斌, 刘娅梅, 孙书存. 南京地区落叶栎林主要木本植物的展叶动态研究. 植物生态学报, 2005, 29(1): 125- 136.
[29] 孙德祥, 何尚仁. 多元分析方法及其应用. 银川: 宁夏人民出版社, 1989: 82- 191.
[30] 北京林学院. 数理统计. 北京: 中国林业出版社, 1980: 98- 104.
[31] Negi G C S. Leaf and bud demography and shoot growth in evergreen and deciduous trees of central Himalaya, India. Trees, 2006, 20(4): 416- 429.
[32] 陈波, 达良俊. 栲树不同生长发育阶段的枝系特征分析. 武汉植物学研究, 2003, 21(3): 226- 231.
[33] Mencuccini M, Martínez-Vilalta J, Vanderklein D, Hamid H A. Size-mediated ageing reduces vigour in trees. Ecology Letters, 2005, 8(11): 1183- 1190.
[34] Matsuzaki J, Norisada M, Kodaira J, Suzuki M, Tange T. Shoots grafted into the upper crowns of tall Japanese cedar (CryptomeriajaponicaD. Don) show foliar gas exchange characteristics similar to those of intact shoots. Trees, 2005, 19(2): 198- 203.
[35] Marquis R J, Diniz I R, Morais H C. Patterns and correlates of interspecific variation in foliar insect herbivory and pathogen attack in Brazilian cerrado. Journal of Tropical Ecology, 2001, 17(1): 127- 148.
[36] Kikuzawa K. A cost-benefit analysis of leaf habit and leaf longevity of trees and their geographical pattern. The American Naturalist, 1991, 138(5): 1250- 1263.
[37] Mediavilla S, Escudero A. Ontogenetic changes in leaf phenology of two co-occurring Mediterranean oaks differing in leaf life span. Ecological Research, 2009, 24(5): 1083- 1090.
Investigation of bud burst, shoot growth and leaf expansion inPopuluseuphraticaof different ages
ZHENG Yaqiong1,2,FENG Mei1,2,LI Zhijun1,2,*
1KeyLaboratoryofBiologicalResourceProtectionandUtilizationofTarimBasin,XinjiangProductionandConstructionGroup,Alar843300,China2CollegeofPlantSciences,TarimUniversity,Alar843300,China
PopuluseuphraticaOliv., a member in the subgenus populous, family Salicaceae, is the oldest and most primitive desert riparian forest species, mainly distributed in central Asia, west Asia and the Mediterranean region. Over 60% of naturalP.euphraticaforest is distributed in China, of which, 89.1% ofP.euphraticais concentrated in the Tarim Basin of Xinjiang. Earlier research indicated that at different growth and development stages ofP.euphraticaindividuals, leaf morphology was different, showing a clear heterophylly. Leaves of seedlings, saplings and young branches sprouted from adult trees are of linear and lanceolate, but with tree growth, ovate and broadly ovate leaves start to form giving the tree a unique look. In adult tree, the top canopy is dominated by broadly ovate leaves and the bottom canopy is of mainly linear and lanceolate leaves. This is known as exclusive or variant leaves ofP.euphratica. Studies also shown that, in the same growing season of the sameP.euphraticaindividual, it is always the case that the broadly ovate leaves on the top canopy start to burst first followed by the linear, lanceolate and ovate leaves at the bottom of the canopy, indicating a vertical inherent coordination of development. A clear relationship among branching, bud growth, leaf phenology and leaf shape change exist in this unique species worth detailed investigation. Most of the previous research was focused on flowering phenology, but the relationship among branching, bud growth, leaf phenology and leaf shape change and their vertical distribution have not been studied in detail at different developmental stages. We examined the development of these organs and their relationships and interactions usingP.euphraticaindividuals at different developmental stages to lay a foundation for revealing the significance of leaf shape changes inP.euphraticalife history.P.euphraticatrees at five different development stages were investigated to study the characteristics of growth and the spatial distribution pattern of leaf shape changes. Results indicated that different growth pattern and leaf phenology were shown both in spatial distribution and phenological developmental stages at different individual canopy levels and trees of different ages. With the increase of tree age and canopy level (from base to top), new shoot length, leaf number per shoot and leaf shape index have decrease trend, but a gradual increase trend of leaf area and leaf dry weight were observed. Regardless of developmental stages, leaf expansion started on the top of the canopy and gradually moved to wards the base. Bud opened in early April, leaf expansion commenced in mid-April and finished from early to late May. Older trees showed earlier bud burst and leaf expansion than younger tress and they also finish leaf expansion early. Different bud burst and leaf expansion were observed on trees of different developmental stages but the commencement and termination of leaf expansion were relatively similar. Leaf emergence period was positively correlated with branch length, leaf number and leaf shape index but negatively correlated with leaf area and leaf dry weight and both correlations were significant.
P.euphratica; emergence and growth of bud burst; leaf phenology; leaf shape index; leaf area; leaf dry weight
国家自然科学基金项目(31060026, U1303101); 国家973计划前期研究专项课题(2012CB723204)
2013- 04- 24;
日期:2014- 04- 11
10.5846/stxb201304240804
*通讯作者Corresponding author.E-mail: lizhijun0202@126.com
郑亚琼,冯梅,李志军.胡杨枝芽生长特征及其展叶物候特征.生态学报,2015,35(4):1198- 1207.
Zheng Y Q,Feng M,Li Z J.Investigation of bud burst, shoot growth and leaf expansion inPopuluseuphraticaof different ages.Acta Ecologica Sinica,2015,35(4):1198- 1207.
猜你喜欢
作物学报(2022年7期)2022-05-11 13:30:26
东坡赤壁诗词(2022年2期)2022-04-15 01:38:01
烟台果树(2021年2期)2021-07-21 07:18:26
内蒙古林业科技(2021年2期)2021-07-08 07:33:04
生态学报(2020年21期)2020-11-24 09:09:42
山东林业科技(2019年6期)2020-01-02 09:36:30
生态学报(2019年21期)2019-12-18 07:25:30
VOGUE服饰与美容(2019年5期)2019-06-11 10:32:06
科学与财富(2017年26期)2017-09-23 09:56:05
花火B(2016年9期)2016-05-14 17:58:11
