
2010年冬季胶州湾浮游植物群落结构特征*
2015-03-08 06:31:21石晓勇王丽莎杨世民
海洋与湖沼 2015年2期
石晓勇 王丽莎 杨世民
(1. 中国海洋大学化学化工学院 青岛 266100; 2. 中国海洋大学船舶中心实验室 青岛 266003;3. 国家海洋局海洋减灾中心 北京 100194)
2010年冬季, 受北方寒潮的影响胶州湾潮间带及浅海大面积结冰, 养殖业损失严重, 这一时期胶州湾的环境状况深受各界关注。至2月初, 除胶州湾北部还有少量海冰外, 其余海域海冰已基本消融, 因此我们及时对胶州湾进行了环境调查。低温的海水环境不利于浮游植物的生长, 海冰的覆盖会造成光照不足, 更加剧了对浮游植物生长的抑制作用(Hegseth et al, 2002; Fonda et al, 2005; 李宏亮等, 2007; 张芳等,2012)。此时的大面站调查, 有助于我们了解低温自然灾害情况下浮游植物群落结构的特点, 本文即是通过本次调查采集的水样对浮游植物群落结构进行了分析研究, 并参阅了以往国内学者的历史资料(钱树本等,1983; 王文海, 1993; 齐雨藻, 2004; 吴玉霖等, 2004,2005; Li et al, 2005; 李艳等, 2005; 杨世民等, 2009),为胶州湾生态系统动力学的研究提供基础资料, 也为胶州湾这一时期海洋环境的评价提供科学依据。
1 材料与方法
胶州湾冬季调查共设立 23个大面站, 其分布见图1。调查时间为2010年2月4日。采样方式依照《海洋调查规范》(国家技术监督局, 1992)进行, 于每站位各水层(0m, 5m, 10m, 20m, 30m, 水深浅于5m的站位只取表层水)采集500mL水样, 样品用1%中性福尔马林固定保存, 水样采集工具为 HQM-2球盖式有机玻璃采水器。采水样品的分析和计数采用国际通用的 Utermöhl方法(Utermöhl, 1958)。取 25mL水样置于Hydro-Bios计数框内, 静置24h后在NikonTS100倒置显微镜下进行浮游植物物种鉴定(金德祥等,1965; 郭玉洁, 2003; 杨世民等, 2006; Tomas, 1997)和细胞计数。
物种多样性指数的计算采用香农-威纳指数(H′,Shannon-Wiener index) (Shannon et al, 1949), 物种均匀度指数(J)采用Pielou的计算公式(Pielou, 1969)。
2 结果与讨论
2.1 物种组成
胶州湾冬季浮游植物样品经初步鉴定, 调查期内共检出浮游植物69种, 分别隶属于5门44属。其中硅藻门32属54种(包括2变型); 甲藻门9属12种;金藻门1属1种; 蓝藻门1属1种; 隐藻门1属1种。种名录见表1。在各站位各水层硅藻在物种和数量上都占绝对优势。硅藻占细胞丰度的80%—100%, 平均98.81%; 甲藻占细胞丰度的0—20%, 平均1.18%; 金藻、蓝藻、隐藻只是在个别站位的个别水层偶尔出现。调查区浮游植物的生态类型多为广布性和温带性物种, 也有少量冷水性物种。
冬季胶州湾浮游植物的主要优势种均为硅藻:柔弱几内亚藻(Guinardia delicatula)、扭链角毛藻(Chaetoceros tortissimus)、诺氏海链藻(Thalassiosira nordenskiöldii)和 尖 刺 伪 菱 形 藻 (Pseudo-nitzschia pungens)。其中柔弱几内亚藻的优势度最高, 为0.213;尖刺伪菱形藻的出现频度最高, 为0.952。
图1 胶州湾冬季取样站位Fig.1 The sampling station in the Jiaozhou Bay in winter
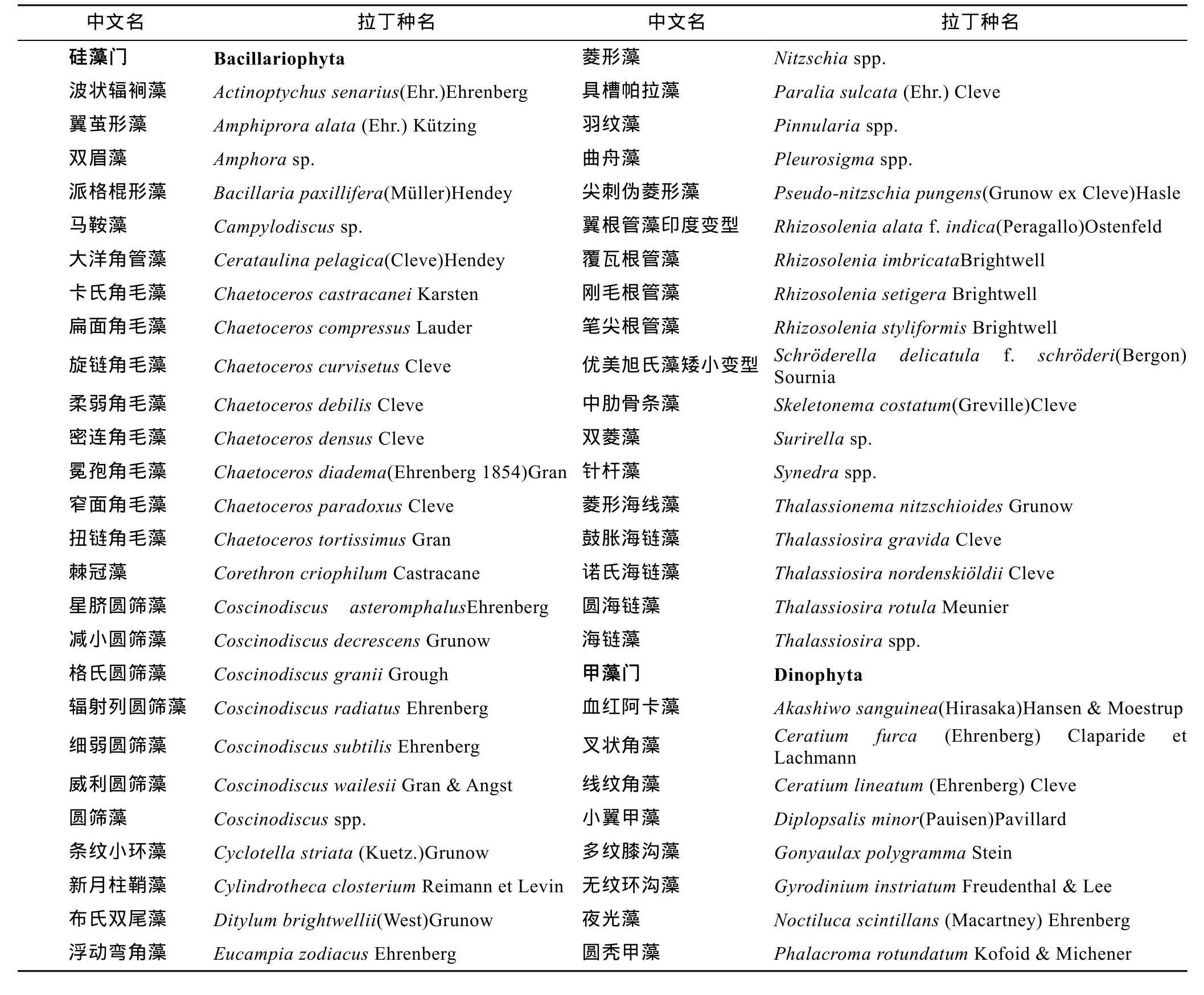
表1 胶州湾冬季浮游植物的物种组成Tab.1 Species composition of phytoplankton assemblage in the Jiaozhou Bay in winter
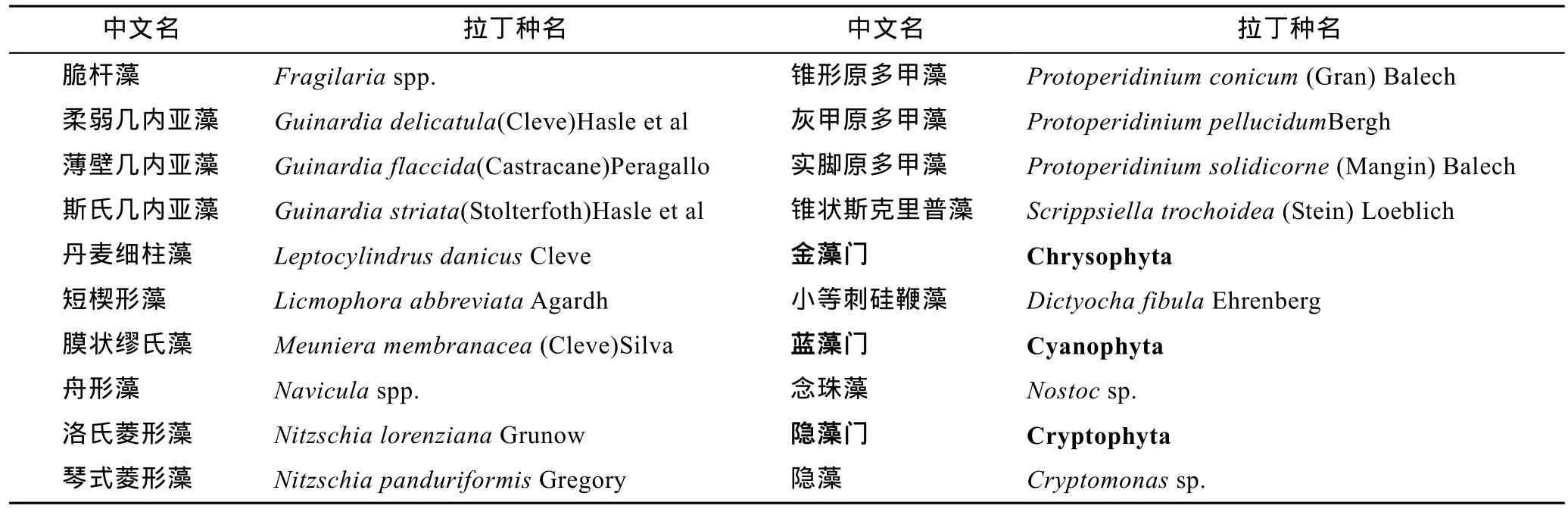
续表
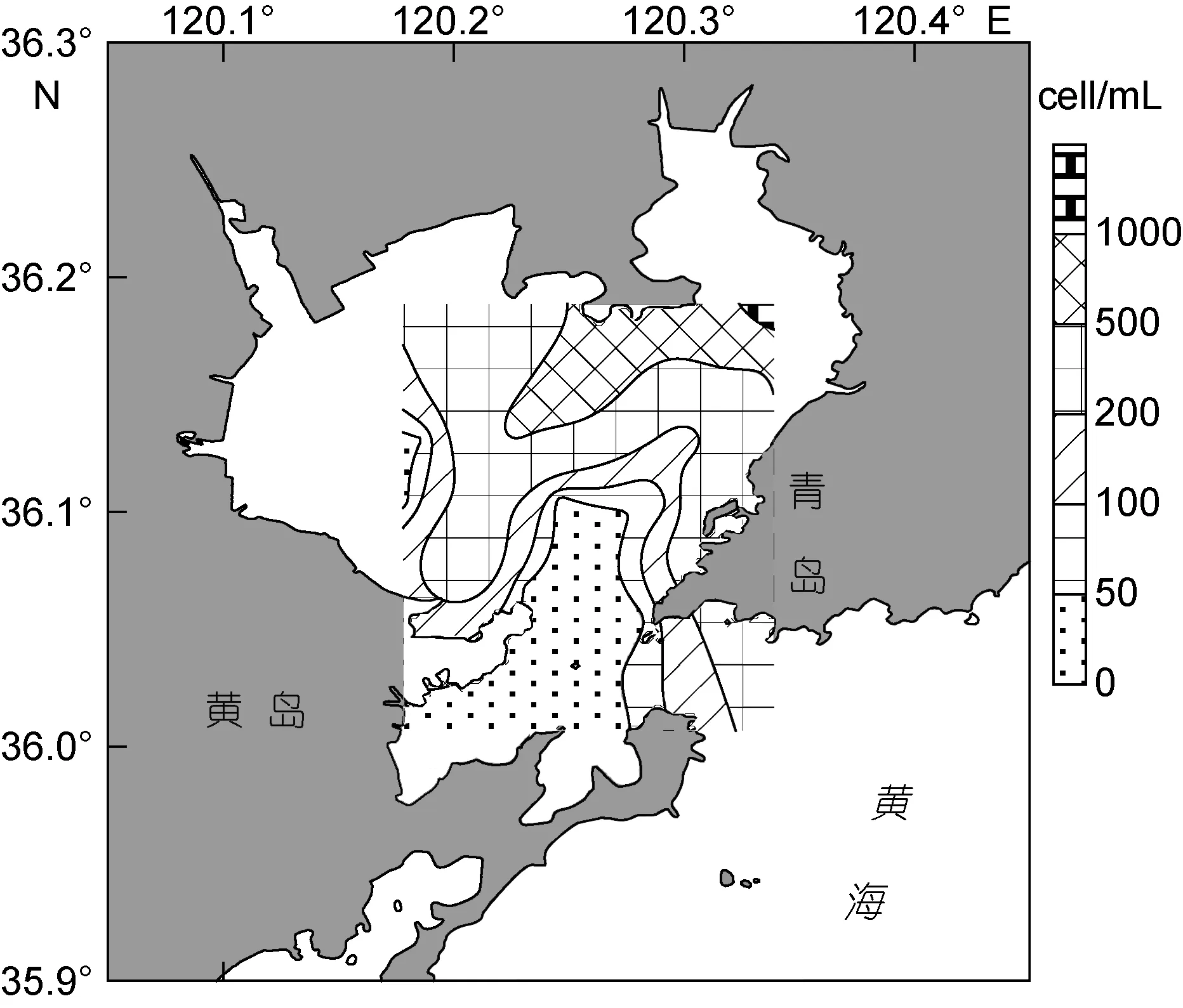
图2 表层水体浮游植物细胞丰度的平面分布Fig.2 Horizontal distribution of phytoplankton cell abundance
2.2 浮游植物的细胞丰度
冬季胶州湾浮游植物的细胞丰度介于 0.5—1270.6cell/mL, 平均 222.8cell/mL, 最高值位于湾东北部区域 9号站表层。硅藻细胞丰度介于 0.4—1270.6cell/mL, 平均 222.4cell/mL, 最高值亦位于 9号站表层; 甲藻细胞丰度介于 0—2.5cell/mL, 平均0.3cell/mL, 最高值位于湾口区域24号站表层。
2.2.1 表层平面分布特征 冬季胶州湾表层水体浮游植物细胞丰度介于 2.5—1270.6cell/mL, 平均275.0cell/mL, 略高于各水层浮游植物平均细胞丰度。最高值位于湾东北部区域9号站, 并在胶州湾东北部形成浮游植物细胞丰度密集区, 总体分布趋势为北部高于南部(图2)。
表层水体硅藻细胞丰度介于2.5—1270.6cell/mL,平均274.7cell/mL。最高值亦位于湾东北部区域9号站, 其分布趋势与表层水体浮游植物细胞丰度的分布趋势极为相似(图3a)。表层水体甲藻细胞丰度介于0—2.5cell/mL, 平均0.3cell/mL, 总体细胞丰度较低。最高值位于湾口区域 24号站, 分布趋势为胶州湾东北部和南部湾口海域较高, 中西部海域较低(图3b)。
与 1977年网采历史调查资料相比(钱树本等,1983), 浮游植物细胞丰度的分布趋势有所不同, 1977年冬季胶州湾浮游植物在湾东南部形成细胞丰度密集区, 但本次调查细胞丰度密集区在湾东北部, 而且本次调查的细胞丰度也低于1977年同期细胞丰度。出现这种情况的原因主要是1977年2月浮游植物群落的优势种中肋骨条藻(Skeletonema costatum)和角毛藻属(Chaetoceros spp.)的物种在本次调查中细胞丰度值较低。尤其是角毛藻属的物种, 1977年冬季在胶州湾东南部形成细胞丰度高值区, 而本次调查中无此情况, 这也是本次调查与 1977年同期细胞丰度分布趋势不同的主要原因。另外, 相对于中肋骨条藻和角毛藻属的物种, 本次调查的最优势物种柔弱几内亚藻更适应低温的环境(杨世民等, 2009), 这也从反面证明了本次调查期间海冰、低温对浮游植物群落结构变化的影响。
2.2.2 垂直分布特征 冬季胶州湾浮游植物的垂直分布如图 4a, 由于胶州湾浅水样品的数量大大多于深水样品的数量, 因此虽然表面上看浮游植物细胞丰度随着水深的增加逐渐降低, 但浮游植物真正的垂直分布是趋向于均匀, 除个别站位外各水层之间细胞丰度相差不大。
冬季胶州湾硅藻垂直分布如图 4b, 其垂直分布趋势与浮游植物的垂直分布趋势也是极为相似。甲藻的垂直分布如图4c, 其主要分布在表层、5m和10m层。如果扣除表层样品较多的因素外, 总体趋势应为由表至底细胞丰度逐渐升高。
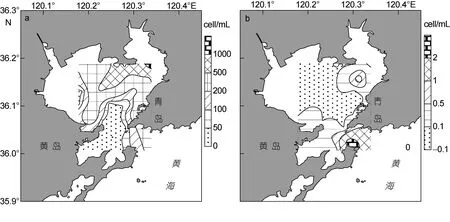
图3 表层水体浮游硅藻细胞丰度和浮游甲藻细胞丰度的平面分布Fig.3 Horizontal distribution of planktonic diatom cell abundance, horizontal distribution of planktonic dinoflagellate cell abundance in surface water
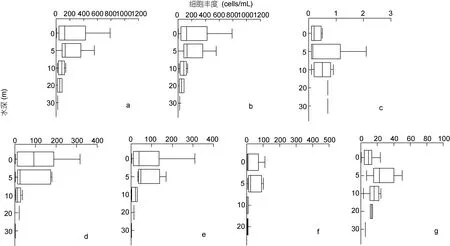
图4 冬季胶州湾浮游植物、硅藻、甲藻及优势种细胞丰度的垂直分布Fig.4 Vertical distribution of phytoplankton cell abundance, planktonic diatom cell abundance, planktonic dinoflagellate cell abundance and dominant species cell abundance in Jiaozhou Bay in winter
2.2.3 表层优势种平面分布特征 胶州湾冬季浮游植物呈现优势种较多、优势种之间优势度差别不明显的特点, 优势种的表层平面分布如图5所示。柔弱几内亚藻为温带近岸性种, 是调查期内胶州湾的最优势物种。细胞丰度介于 0—317.2cell/mL, 平均62.5cell/mL, 最高值位于湾西北部区域14号站表层。柔弱几内亚藻的出现频度并不高, 在优势种中是最低的, 但其出现的细胞丰度较高的站位最多。在表层水体分布的高值区位于胶州湾北部水域, 呈现北高南低的分布趋势。
扭链角毛藻为北温带近岸性种。细胞丰度介于0—310.0cell/mL, 平均 55.8cell/mL, 最高值位于湾东北部区域9号站表层。扭链角毛藻主要出现在胶州湾东部和北部海域, 出现频度为 0.833。在表层水体分布的高值区亦位于胶州湾东部和北部, 呈现东北高、西南低的分布趋势。
诺氏海链藻为北方或北极近海性种。细胞丰度介于0—474.6cell/mL, 平均37.8cell/mL, 最高值亦位于湾东北部区域9号站表层。诺氏海链藻主要出现在胶州湾北部海域, 出现频度同样为 0.833。在表层水体的分布趋势为北高南低。
尖刺伪菱形藻为广温性近岸种。细胞丰度介于 0—87.0cell/mL, 平均 16.6cell/mL, 最高值位于湾中部区域26号站10m层。尖刺伪菱形藻在胶州湾分布很广, 出现频度在优势种中最高。在表层水体的分布趋势为湾中部较高, 由此向北、向南逐渐降低。
与历史资料相比(钱树本等, 1983; 齐雨藻, 2004;杨世民等, 2009)这些物种的优势种地位并没有发生改变, 尤其是柔弱几内亚藻在青岛前海细胞丰度曾达到近103cell/mL(杨世民等, 2006, 2009), 因此其在今后的调查研究中应予以重点关注。
2.2.4 优势种垂直分布特征 冬季胶州湾浮游植物优势种柔弱几内亚藻和扭链角毛藻的垂直分布如图4d、4e, 其垂直分布趋势与浮游植物总的分布趋势相似, 扣除表层样品较多的因素, 垂直分布是趋向于均匀, 各水层之间细胞丰度相差不大。
优势种诺氏海链藻和尖刺伪菱形藻的垂直分布如图4f、g, 其分布趋势与甲藻相似, 总体趋势为由表至底细胞丰度逐渐升高。
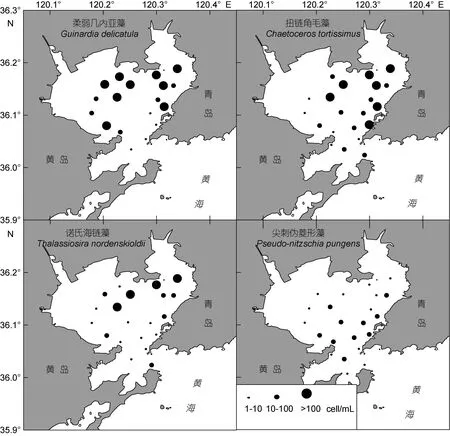
图5 表层水体浮游植物优势种细胞丰度的平面分布Fig.5 Horizontal distribution in cell abundance of dominant phytoplankton species
2.3 浮游植物群落的多样性
冬季胶州湾浮游植物的 Shannon-Wiener多样性指数值介于0.497—3.404, 平均值2.335, 其值总体较高。多样性指数在表层水体的平面分布见图6a, 高值区位于胶州湾口, 而低值区位于胶州湾西北部海域。胶州湾冬季浮游植物优势种较多, 在某一站位某一水层往往有多个优势种出现, 从而形成了较为复杂的群落结构, 而在湾西北部海域, 可能受到海冰等自然条件的影响, 浮游植物形成了以柔弱几内亚藻为单一优势种的较为简单的群落结构。
冬季胶州湾浮游植物的均匀度指数值介于0.150—0.831, 平均值 0.600, 其值亦总体较高。均匀度指数在表层水体的平面分布见图 6b, 其分布趋势为东南至胶州湾口为高值区, 由此向西北逐渐降低,因此胶州湾冬季东南至胶州湾口海域浮游植物群落更加稳定。
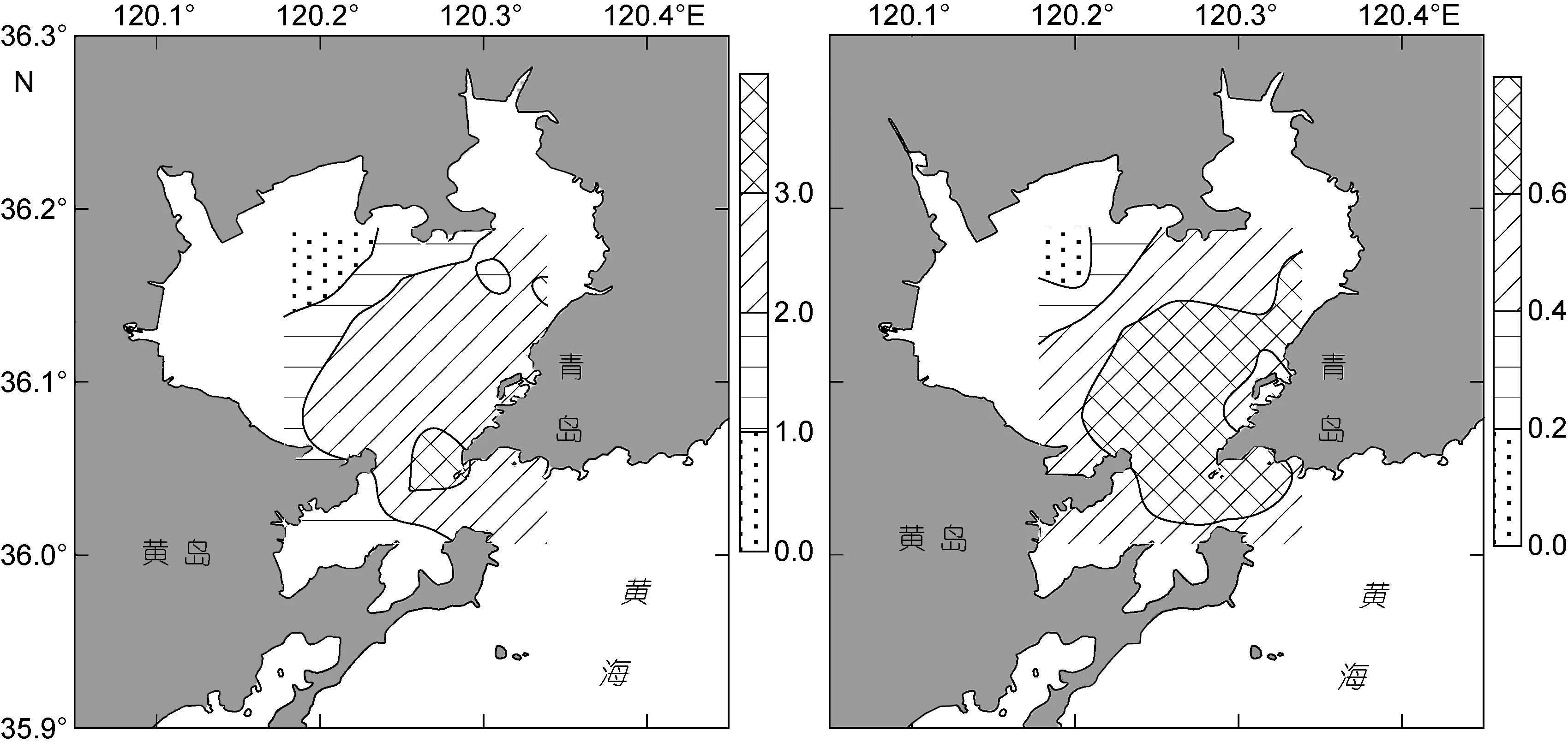
图6 表层水体Shannon-Wiener指数和均匀度指数的平面分布Fig.6 Horizontal distribution of Shannon-Wiener diversity index and evenness index in surface water
2.4 浮游植物与环境因子的关系
胶州湾冬季浮游植物细胞丰度与环境因子的相关性分析(表2)表明, 浮游植物细胞丰度和硅藻细胞丰度与铵盐、硝酸盐、亚硝酸盐、硅酸盐均呈密切的正相关, 优势种中柔弱几内亚藻、扭链角毛藻、诺氏海链藻亦与此四种营养盐呈密切的正相关。本次调查期间浮游植物和多数优势种与氮、硅的相关关系非常显著, 说明氮、硅是这一时期促进浮游植物生长的主要因素。而调查期间硅酸盐浓度总体较低, 这是由于硅藻生长繁殖需要硅酸盐组成硅质细胞壁和进行生物合成的调节(De La Rocha et al, 1998;杨小龙等, 1990; 彭欣等, 2006), 从而消耗了大量的硅, 这也从另一方面证明了硅酸盐对浮游植物生长的促进作用。
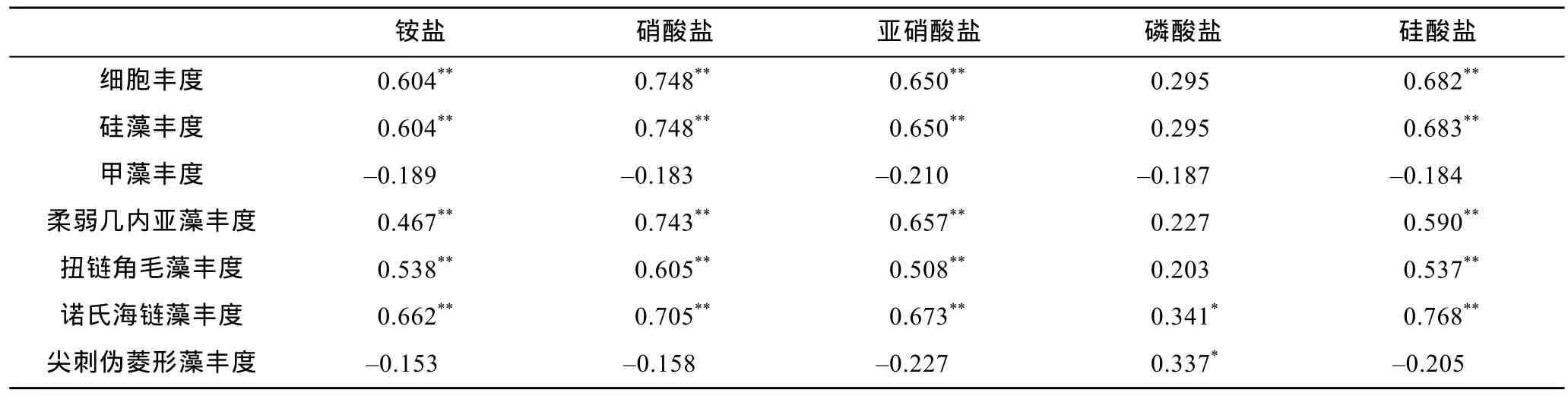
表2 浮游植物细胞丰度与环境因子的Pearson相关分析Tab.2 Pearson correlation analysis between phytoplankton cell abundance and environmental factors
2.5 浮游植物的季节变化
本次调查与2009年5月春季(杨世民等, 2014)、2009年8月夏季(杨世民等, 2015)的调查结果相比较,虽然细胞丰度低于 2009年春季, 但从与环境因子显著的相关关系可以看出浮游植物的生长非常旺盛,说明随着胶州湾水温的升高、海冰的消融, 浮游植物开始利用营养盐等有利因素大量繁殖, 这一时期浮游植物的生长状态是三次调查中最好的。
3 结论
(1) 2010年冬季胶州湾共发现浮游植物 5门 44属 69种, 其中硅藻在物种数量和细胞丰度上都占有绝对优势。浮游植物物种的生态类型多为广布性和温带性物种, 也有少量冷水性物种。
(2) 胶州湾冬季平均细胞丰度 222.8cell/mL, 平面分布趋势为北部高于南部, 垂直分布则是趋向于均匀, 各水层之间细胞丰度相差不大。
(3) 调查期内胶州湾浮游植物优势种为柔弱几内亚藻、扭链角毛藻、诺氏海链藻和尖刺伪菱形藻。平面分布除尖刺伪菱形藻高值区位于胶州湾中部海域外, 其余优势种均呈现细胞丰度北高南低的分布趋势; 垂直分布趋势柔弱几内亚藻和扭链角毛藻为各水层之间细胞丰度相差不大, 而诺氏海链藻和尖刺伪菱形藻为由表至底细胞丰度逐渐升高。
(4) 冬季胶州湾浮游植物 Shannon-Wiener指数和均匀度指数的值均较高, 其分布显示胶州湾东南至胶州湾口海域浮游植物群落更加稳定, 而湾西北部海域浮游植物群落较为单一, 这种分布趋势可能与胶州湾西北部海域海冰的存在有关。
(5) 胶州湾冬季浮游植物和硅藻细胞丰度以及优势种中的柔弱几内亚藻、扭链角毛藻、诺氏海链藻与铵盐、硝酸盐、亚硝酸盐、硅酸盐均呈显著的正相关关系。
王文海, 1993. 中国海湾志, 第四分册(山东半岛南部和江苏省海湾). 北京: 海洋出版社, 157—260
齐雨藻, 2004. 中国沿海赤潮. 北京: 科学出版社, 1—348
李宏亮, 陈建芳, 刘子琳等, 2007. 北极楚科奇海和加拿大海盆南部颗粒生物硅的粒级结构. 自然科学进展, 17(1):72—78
李 艳, 李瑞香, 王宗灵等, 2005. 胶州湾浮游植物群落结构及其变化的初步研究. 海洋科学进展, 23(3): 328—334
杨小龙, 朱明远, 1990. 浮游植物营养代谢研究新进展. 黄渤海海洋, 8(3): 65—74
杨世民, 董树刚, 2006. 中国海域常见浮游硅藻图谱. 青岛:中国海洋大学出版社, 1—267
杨世民, 董树刚, 窦明武等, 2009. 2004—2005年青岛前海定点54周次浮游植物群落结构特征. 海洋科学进展, 27(4):523—536
杨世民, 王丽莎, 石晓勇, 2014. 2009年春季胶州湾浮游植物群落结构特征. 海洋与湖沼, 45(6): 1234—1240
杨世民, 王丽莎, 石晓勇, 2015. 2009年夏季胶州湾同步调查浮游植物群落结构特征. 海洋与湖沼, 46(1): 102—108
吴玉霖, 孙 松, 张永山等, 2004. 胶州湾浮游植物数量长期动态变化的研究. 海洋与湖沼, 35(6): 518—523
吴玉霖, 孙 松, 张永山, 2005. 环境长期变化对胶州湾浮游植物群落结构的影响. 海洋与湖沼, 36(6): 487—498
张 芳, 何剑锋, 郭超颖等, 2012. 夏季北冰洋楚科奇海微微型、微型浮游植物和细菌的丰度分布特征及其与水团的关系. 极地研究, 24(3): 238—246
国家技术监督局, 1992. 海洋调查规范. 北京: 中国标准出版社, 17—22
金德祥, 陈金环, 黄凯歌, 1965. 中国海洋浮游硅藻类. 上海:上海科学技术出版社, 1—230
钱树本, 王筱庆, 陈国蔚, 1983. 胶州湾的浮游藻类. 山东海洋学院学报, 13(1): 39—56
郭玉洁, 2003. 中国海藻志(第五卷, 硅藻门, 第一册中心纲).北京: 科学出版社, 1—493
彭 欣, 宁修仁, 蔡昱明等, 2006. 浮游植物生长上行效应的研究进展. 海洋学研究, 24(3): 64—75
De La Rocha C L, Brzezinski M A, DeNiro M J et al, 1998.Silicon-isotope composition of diatoms as an indicator of past oceanic change. Nature, 395(6703): 680—683
Fonda U S, Monti M, Bergamasco A et al, 2005. Plankton community structure and dynamics versus physical structure from Terra Nova Bay to Ross Ice Shelf (Antarctica). Journal of Marine Systems, 55(1—2): 31—46
Hegseth E N, Von Quillfeldt C H, 2002. Low phytoplankton biomass and ice algal blooms in the Weddell Sea during the ice-filled summer of 1997. Antarctic Science, 14(3):231—243
Li G Y, Lu J, He Y J, 2005. Relation between diversity of phytoplankton and environmental factors in the Jiaozhou Bay. Marine Geology Letters, 21(4): 10—13
Pielou E C, 1969. An Introduction to Mathematical Ecology. New York: Wiley-Interscience, 1—286
Shannon C E, Weaver W, 1949. The Mathematical Theory of Communication. Urbana IL: University of Illinois Press,1—125
Tomas C R, 1997. Identifying Marine Phytoplankton. San Diego:Academic Press, 1—858
Utermöhl H, 1958. Zur Vervollkommnung der quantitativen Phytoplankton-Methodik. Mitteilungen der Internationale Vereinigung für Theoretische und Angewandte Limnologie,9: 1—38
猜你喜欢
Chinese Physics B(2024年3期)2024-03-25 09:33:24
大自然探索(2023年7期)2023-11-14 13:07:36
课堂内外(小学版)(2023年9期)2023-10-11 14:39:26
人民黄河(2022年4期)2022-04-07 09:03:16
黑龙江水利科技(2020年8期)2020-01-12 06:26:00
东方企业家(2018年8期)2018-08-22 05:44:08
上海建材(2018年1期)2018-04-18 12:15:16
欧亚学刊(2015年1期)2015-09-04 06:12:31
Chinese Journal of Chemical Engineering(2014年4期)2014-07-18 11:56:14
中国海洋大学学报(自然科学版)(2014年6期)2014-02-28 12:20:57

- 海洋与湖沼的其它文章
- 俄罗斯鲟(Acipenser gueldenstaedtii)家系构建及不同家系间生长性能比较*
- 杂色鲍(Haliotis diversicolor)硒结合蛋白1基因的克隆及其应激表达*
- 盐度调控对花鲈(Lateolabrax maculatus)生长相关基因表达的影响*
- 鳗弧菌(Vibrio anguillarum)侵染对青蛤(Cyclina sinensis)髓样分化因子88基因表达的影响*
- 连续降温对大菱鲆(Scophthalmus maximus)成鱼血清生化指标及Wap65-1基因表达的影响*
- 洞庭青鲫(Carassius auratus var. Dongtingking)与三个鲫品系群体遗传多样性的ISSR分析*