
大型浅水湖泊太湖中浮游病毒的时空分布特征及其与环境因子的相关性分析*
2015-03-08 05:27段翠兰胡志新陈光芸王一鸣张永江
海洋与湖沼 2015年4期
段翠兰 胡志新 邹 勇 陈光芸 陈 静 王一鸣 张永江①
(1. 江苏省渔业生态环境监测站 南京 210036; 2. 南京工程学院环境工程学院 南京 211167)
浮游病毒是湖泊生态系统中的重要组成部分(张全国,2014),在微食物环中起着重要的作用(Bratbaket al,1994)。水体中的浮游病毒数量巨大,主要为噬菌体(张奇亚,2014)和浮游藻类病毒(赵以军等,1996),它们通常会感染水体中的浮游细菌和浮游藻类,导致浮游细菌和浮游藻类的死亡、裂解,从而对水体中浮游细菌和浮游藻类的丰度和种群产生重要影响(Liuet al,2006)。
对浮游病毒的研究始于20世纪60年代,后随着电子显微镜的广泛应用(Torrellaet al,1979),浮游病毒成为生态学的研究热点。浮游病毒的类群、丰度、生态作用以及差异性进行了一系列研究(Wilhelmet al,2008)。有学者对全球的浮游病毒总量进行估算,发现浮游病毒可能是仅次于原核生物的第二大生物量群(孙辉,2014)。目前,国内对海洋浮游病毒的生态分布的认识已较为全面(李洪波等,2012; 王艳等,2013),湖泊淡水浮游病毒的研究还较为薄弱,仅见对武汉东湖(刘艳鸣等,2005)、长江口(白晓歌等,2007)、湖北湿地(孙小磊等,2009)等的初步探索。太湖是我国第三大淡水湖泊,空间异质性高,具有草、藻型两个截然不同的生态系统,风浪对水体扰动作用明显,无温跃层,四季温差大,是一个典型的大型浅水湖泊(秦伯强等,2004); 太湖又是国家重点治理的富营养水体之一。为了揭示太湖水体中浮游病毒的分布规律,了解其在微食物环中的生态功能,本实验采用表面荧光计数法,研究了太湖水体中浮游病毒丰度的时空分布特征,探讨了影响太湖水体中病毒丰度的主要环境因子,为进一步揭示太湖水体中浮游病毒的生态位提供基础。
1 材料与方法
1.1 采样点设置与样品采集
为研究太湖水体中病毒的时空分布,在太湖不同功能区域中共设置 19个采样点位,位置见图1。于2013年9月和2014年5月,在上述采样点位用5 L的Niskin采水器,采集水样。将50 mL的水样,放入事先清洗干净、灭菌的玻璃瓶中,加入戊二醛固定(终浓度为 0.5%),4°C低温保存,用于细菌、病毒的分析,样品在1个月内分析完毕。另将2.5 L的水样放入事先清洗干净的塑料瓶中,用于叶绿素a及其它相关水质参数的分析。
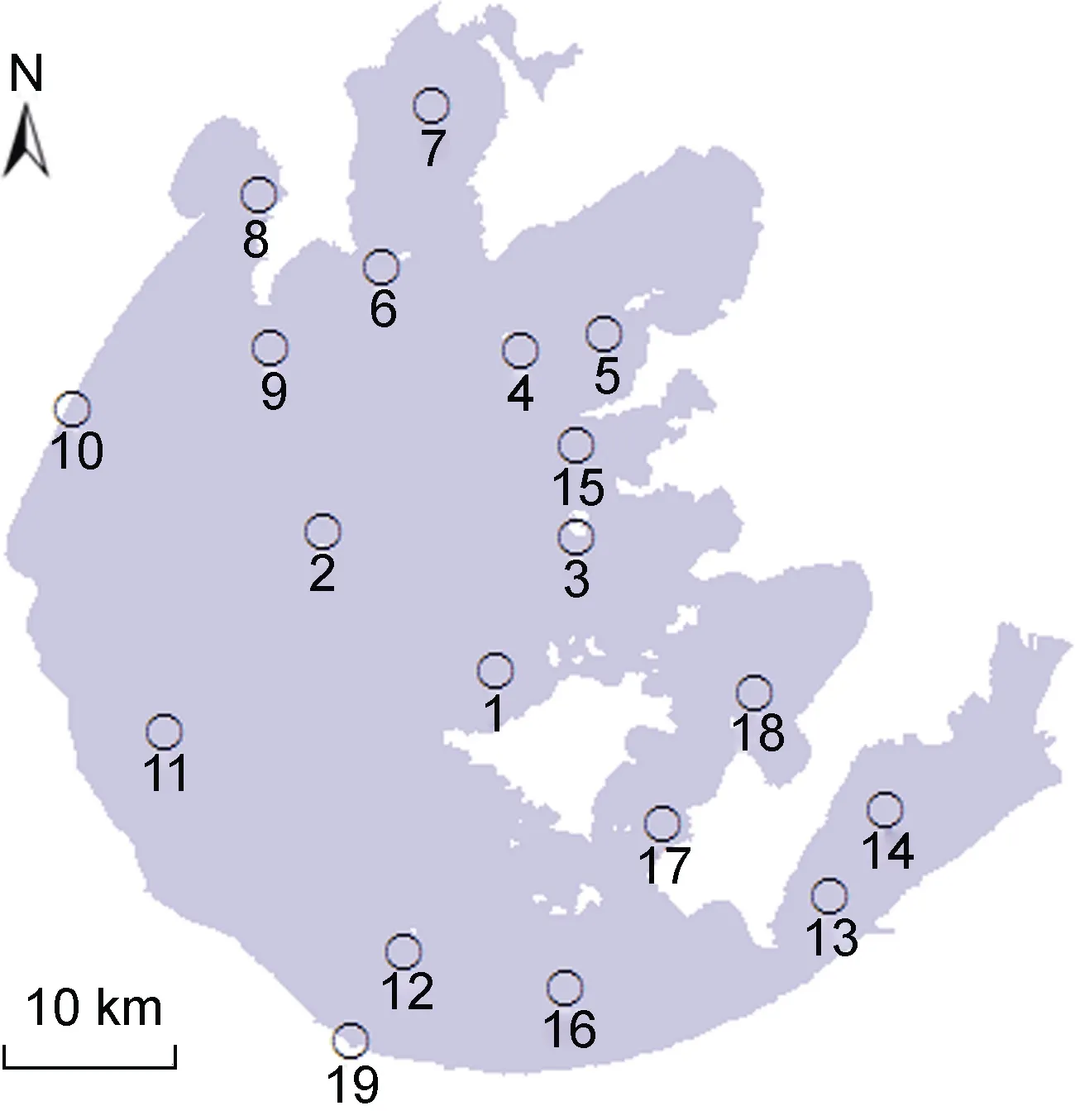
图1 太湖采样点位图Fig.1 Sampling sites in Taihu Lake
1.2 样品的分析
1.2.1 浮游病毒和浮游细菌的分析 取1 mL上述固定后的样品,经SYBR Green I染色后,利用荧光显微镜(Nikon 80i,蓝光,油镜100×) 测定浮游细菌、浮游病毒的丰度(Patelet al,2007; Suttleet al,2010)。
1.2.2 叶绿素a的分析 取200 mL的水样,用直径25 mm的GF/F滤膜过滤后,将滤膜剪碎、用90%的丙酮,–20°C 黑暗条件下萃取 24 h,离心后,用Thermo scientific LUMINA Fluorescence Spectrometer测定其荧光强度,换算为叶绿素a浓度(梁玉波等,2006)。过滤后的滤膜,如不能立即测定,可用铝箔包装后,–20°C 保存。
1.3 数据分析与绘图
数据分析和统计使用 SPSS16.0软件,绘图使用MapInfo7.0和Excel软件。
2 结果
2.1 太湖水体中浮游病毒的空间分布特征
在太湖水体中,浮游病毒的丰度呈现出显著的空间差异。在不同样点间,其丰度变动于(0.91—74.61)×106ind./mL 之间,均值为(16.38±1.17)×106ind./mL (图 2)。

图2 太湖浮游病毒丰度的水平分布(×106 ind./mL)Fig.2 The horizontal distribution of virioplankton abundance in Taihu Lake
根据采样点位,将采样区域划分为四个类型,分别为敞水区(点位1—12)、网围养殖区(点位13—14)、保护区(点位15—18)、入湖河道(点位19)。太湖水体中浮病毒的丰度具有显著的空间差异,网围养殖区的病毒丰度较高,为(3.52±0.28)×107ind./mL,敞水区的病毒丰度较低,为(4.06±0.90)×106ind./mL(图 3)。
2.2 太湖水体中浮游病毒丰度的季节变动
从季节变化来看,春季的浮游病毒丰度变化范围是(0.91—13.22)×106ind./mL(表 1),均值为 4.64×106ind./mL,秋季的浮游病毒丰度变化范围是(3.55—74.61)×106ind./mL,均值为 28.31×106ind./mL,浮游病毒的丰度呈现出显著的时间差异(图 3),秋季水体中浮游病毒的丰度(2.83±0.74)×107ind./mL,显著大于春季(3.11±0.67)×106ind./mL (图 4)。
2.3 病毒与细菌比率
病毒与细菌比率(virus-to-bacteria ratio,VBR)一般用来分析原核生物与病毒之间的关系(Wommacket al,2000)。在此次太湖研究中,VBR变化范围很广,最大值为 100.28,出现在秋季敞水区乌龟山测点附近;最小值为 1.01,出现在春季敞水区平台山测点附近;太湖 VBR平均值为 16.93。VBR的季节变化表现为秋季>春季(表 1)。VBR与异养细菌丰度在春、秋两个季节中均无显著的相关性。
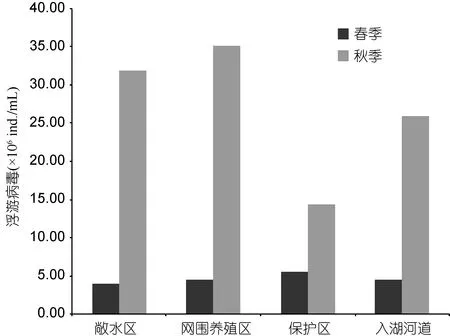
图3 太湖浮游病毒丰度的区域分布Fig.3 The spatial virioplankton distribution in Taihu Lake
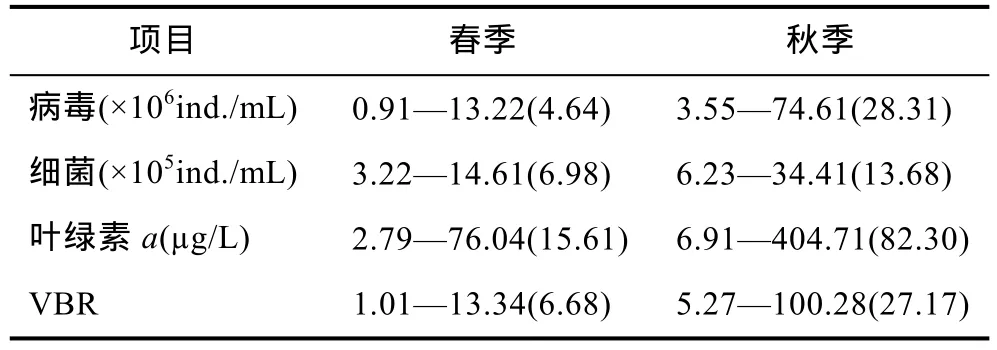
表1 浮游病毒、细菌、叶绿素a和VBR丰度的季节变化Tab.1 The values of virioplankton,bacterioplankton,Chl a and VBR in two seasons
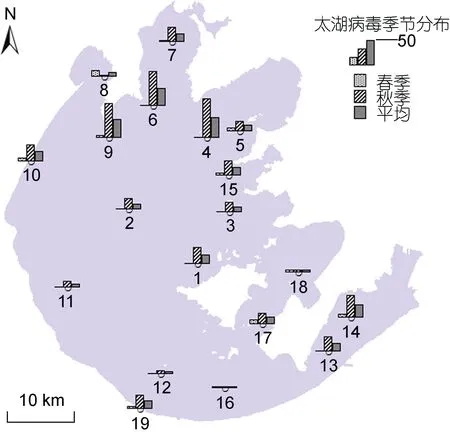
图4 太湖浮游病毒丰度的季节分布(×106 ind./mL)Fig.4 The seasonal distribution of virioplankton in Taihu Lake
2.4 病毒与叶绿素a、细菌的关系
浮游植物和细菌作为浮游病毒的宿主,与其有较强的相关性。而在不同季节,它们之间的相关性存在一些差异(表2)。春季浮游病毒与叶绿素a之间有显著正相关(r=0.645,P<0.05),秋季浮游病毒与叶绿素a之间有极显著正相关(r=0.756,P<0.01)。春、秋季浮游病毒与异养细菌之间无显著相关性。

表2 浮游病毒与叶绿素a、异养细菌的相关系数Tab.2 The correlation coefficient of virioplankton to Chl a and bacterioplankton
3 讨论
根据现有文献资料报道(表 3),我国淡水浮游病毒丰度范围较广,最小值出现在 2014年春季太湖大雷山区域,为 0.91×106ind./mL,而最大值出现在2003年武汉东湖,为2.44×109ind./mL。这表明浮游病毒丰度在不同季节、不同区域变化较大。太湖的病毒丰度在(0.91—47.91)×106ind./mL,而东湖的病毒丰度高达(2.40±0.04)×109ind./mL。在太湖中,可能因其水体面积大,浮游病毒的丰度较低,明显不同于中小型淡水水体的分布情况,其分布状况和江苏近岸海域中的情况类似(段翠兰等,2012)。
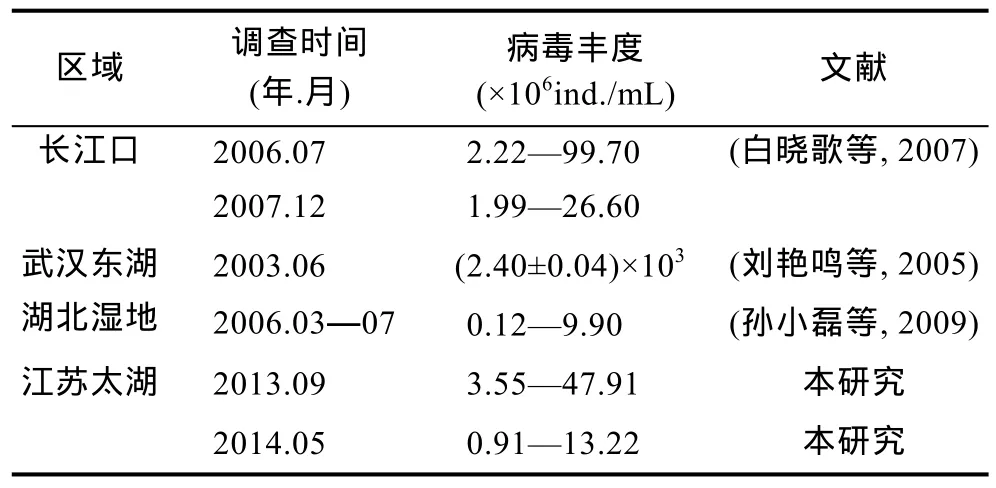
表3 我国淡水浮游病毒丰度分布状况Tab.3 The virioplankton abundance in Chinese fresh waters
Cochran等(1998)研究发现,一般浮游病毒丰度最低值出现于冬季,而最高值出现于秋季。本研究结果表明太湖浮游病毒丰度秋季大于春季,秋季的浮游病毒丰度远远超过春季,差异极为显著。
Vrede等(2003)研究认为细菌丰度与叶绿素a浓度在特定情况下可预测浮游病毒的丰度。干净水体中,浮游病毒丰度与叶绿素a浓度的相关性较高,说明浮游藻类病毒比重较大(Peduzziet al,2004)。在太湖以藻型湖与草型湖为主体的大型水体中,春季浮游病毒丰度与叶绿素a浓度之间呈显著正相关; 秋季浮游病毒丰度与叶绿素a浓度之间呈极显著正相关; 春、秋季浮游病毒与异养细菌之间无显著相关性,这说明浮游藻类病毒在太湖水体中所占比重较大,是太湖的优势种类。而孙小磊等(2009)对湖北湿地研究结果显示,浮游病毒与活菌数的相关性高于与叶绿素a浓度的相关性,认为噬菌体是湿地水体中浮游病毒的优势种类。
一般情况,VBR介于3到 10; 在初级生产力高,营养盐丰富的区域,VBR值会提高(Wommacket al,2000)。本研究中,VBR范围为1.01—100.28,平均值为16.93。武汉东湖水体浮游病毒的研究中,VBR范围为49—56(刘艳鸣等,2005)。从VBR值可看出,太湖VBR值范围比较广,而东湖VBR值范围相对集中,这可能与太湖不同水域的富营养化程度不同,而东湖整个湖区的富营养化程度比较相似的状况有关。这进一步表明太湖有些湖区(如东太湖水草区),营养化程度低于东湖,水质状况优于东湖,而有些湖区(如梅梁湾),营养化程度远远超过东湖,富营养化现象严重。
有研究报道在高浮游植物丰度情况下会产生较高的VBR(Paulet al,1991)。在太湖研究中,发现春季浮游病毒与叶绿素a呈显著正相关(r=0.645,P<0.05),秋季浮游病毒与叶绿素a之间呈极显著正相关(r=0.756,P<0.01)。这说明在春季42% (r2=0.42)的浮游病毒与浮游植物有关,秋季 56%的浮游病毒与浮游植物有关,太湖水体浮游病毒的主要宿主是浮游植物。
Wommack等(2000)年研究发现浮游细菌丰度与VBR间的相关性可作为宿主菌种群多样性指标。在宿主种群较单一时,异养细菌丰度与 VBR呈现负相关性; 而宿主菌种群较多样时,异养细菌丰度与VBR无相关性(赵苑等,2010)。通过相关性分析发现,在太湖春季和秋季水体中,异养细菌与 VBR之间无显著相关性。由此可以认为太湖水域中,噬菌体的宿主菌种群多样性较高。这可能与太湖的地理特征有关,浮游病毒各种类间竞争激烈,优势种不明显。
4 结论
太湖水体中浮游病毒的时空分布主要有五个特点。(1) 太湖因其独特的地理特征,浮游病毒的丰度较低,分布不均衡,明显不同于中小型淡水水体中的分布。(2) 太湖水体中浮游病毒的丰度,具有显著的季节差异,秋季水体中浮游病毒的丰度远高于春季。(3) 浮游病毒丰度与叶绿素a浓度具有显著正相关性。浮游藻类病毒是太湖水体的优势种类,噬菌体所占比重较小。(4) 春季42%的浮游病毒是由浮游植物决定的,秋季 56%的浮游病毒是由浮游植物决定的,浮游病毒的主要宿主是浮游植物。(5) 太湖水域中,VBR与异养细菌之间无显著相关性。噬菌体的宿主菌种群多样性较高,优势种不明显。
王 艳,汪 岷,杨 琳等,2013. 南黄海秋季浮游病毒丰度分布及其与宿主和环境因子的相关性研究. 海洋与湖沼,44(1): 198—204
白晓歌,汪 岷,马晶晶等,2007. 冬季和春季长江口及其近海水域浮游病毒丰度的分析. 海洋与湖沼,38(4):368—372
刘艳鸣,张奇亚,袁秀平,2005. 武汉东湖浮游病毒的丰度及多样性. 水生生物学报,29(1): 1—6
孙 辉,2014. 秦皇岛扇贝养殖区浮游病毒丰度及多样性的研究. 青岛: 中国海洋大学硕士学位论文,13
孙小磊,赵以军,刘 妮等,2009. 淡水湿地浮游病毒的空间分布. 生态学报,29(2): 1048—1054
李洪波,杨 青,周 峰,2012. 海洋微食物环研究新进展.海洋环境科学,31(6): 927—932
张全国,2014. 病毒及其生态功能. 生命科学,26(2): 107—111
张奇亚,2014. 噬藻体生物多样性的研究动态. 微生物学通报,41(3): 545—559
赵 苑,武洪庆,李洪波等,2010. 胶州湾浮游病毒的分布研究. 海洋科学,34(7): 86—92
赵以军,石正丽,1996. 真核藻类的病毒和病毒类粒子(VLPs).中国病毒学,11(2): 93—102
段翠兰,李洪波,邹 勇等,2012. 江苏沿岸海域浮游病毒的时空分布. 水生生物学报,36(5): 971—977
秦伯强,胡维平,陈伟民等,2004. 太湖水环境演化过程与机理. 北京: 科学出版社,1—2
梁玉波,宁修仁,王长云等,2006. 海洋生物生态调查技术规程. 北京: 海洋出版社,13—19
Bratbak J,Thingstad F,Heldal M,1994. Viruses and the microbial loop. Microbial Ecology,28(2): 209—221
Cochran P K,Paul J H,1998. Seasonal abundance of lysogenic bacteria in a subtropical estuary. Applied and Environmental Microbiology,64(6): 2308—2312
Liu Y M,Zhang Q Y,Yuan X Pet al,2006. Seasonal variation of virioplankton in a eutrophic shallow lake. Hydrobiologia,560(1): 323—334
Patel A,Noble R T,Steele J Aet al,2007. Virus and prokaryote enumeration from planktonic aquatic environments by epifluorescence microscopy with SYBR Green I. Nature Protocols,2(2): 269—276
Paul J H,Cazares L H,David A Wet al,1991. The distribution of dissolved DNA in an oligotrophic and a eutrophic river of Southwest Florida. Hydrobiologia,218(1): 53—63
Peduzzi P,Schiemer F,2004. Bacteria and viruses in the water column of tropical freshwater reservoirs. Environmental Microbiology,6(7): 707—715
Suttle C A,Fuhrman J A,2010. Enumeration of virus particles in aquatic or sediment samples by epifluorescence microscopy.In: Wilhelm S W,Weinbauer M G,Suttle C A eds. Manual of Aquatic Viral Ecology. Waco TX: American Society of Limnology and Oceanography,145—153
Torrella F,Morita R Y,1979. Evidence by electron micrographs for a high incidence of bacteriophage particles in the waters of Yaquina Bay,oregon: ecological and taxonomical implications. Applied and Environmental Microbiology,37(4): 774—778
Vrede K,Stensdotter U,Lindström E S,2003. Viral and bacterioplankton dynamics in two lakes with different humic contents. Microbial Ecology,46(4): 406—415
Wilhelm S W,Matteson A R,2008. Freshwater and marine virioplankton: a brief overview of commonalities and differences. Freshwater Biology,53(6): 1076:LINK
Wommack K E,Colwell R R,2000. Virioplankton: viruses in aquatic ecosystems. Microbiology and Molecular Biology Reviews,64(1): 69—114
猜你喜欢
作文与考试·小学高年级版(2021年17期)2021-09-23
潍坊学院学报(2020年2期)2021-01-18
东坡赤壁诗词(2020年4期)2020-09-02
生物工程学报(2020年6期)2020-07-31
少儿美术(2019年2期)2019-12-14
流行色(2019年10期)2019-12-06
环境科技(2016年2期)2016-11-08
学习月刊(2015年5期)2015-07-09
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28
