
盐碱胁迫对喜树幼苗生长及体内离子选择性运输的影响1)
2015-03-07 07:16郁万文曹福亮蔡金峰高长忠
东北林业大学学报 2015年11期
郁万文 曹福亮 蔡金峰 高长忠
(南京林业大学,南京,210037) (盐城市红多草莓专业合作社)
责任编辑:任 俐。
全球土壤盐碱化日益严重[1-2],已成为限制世界农林业进一步发展的重要环境因子之一。我国现代盐碱土壤、残余盐碱土壤和潜在盐碱化土壤总面积达到9 913.3 万hm2[3],主要分布于西北、华北、东北和滨海地区,严重影响着我国现代化农林业的发展。因此,植物耐盐/碱性及其对盐/碱胁迫适应机理研究日益受到重视。许多研究发现,中性盐胁迫与碱性盐胁迫实际是两种性质不同的胁迫[4],二者间有一定的相关性,但其对植物的胁迫作用机制以及植物对其生理适应机制均有明显不同[5-6]。碱胁迫比盐胁迫具有更大的生态破坏力[7],由于土壤所含盐分的种类、比例、数量等各不相同,致使土壤碱化往往与盐化交织在一起相伴发生,造成了问题的复杂性。迄今为止,有关植物抗盐碱研究仍多以NaCl 为研究对象,以Na+离子代谢、抗盐性相关基因的分子生物学及盐胁迫信息传导等为主要研究方向。尽管目前在认识植物抗盐机制方面有了长足进步,但在碱性盐胁迫及盐碱混合胁迫方面仍研究很少。我国林木资源丰富,生态类型多,但有关耐水树种的耐盐碱机理的研究相对缺乏。刘慧娟[8]、郝峰鸽等[9-10]对喜树的生长及生理生化特性对NaCl 的响应进行了研究,张露婷等[11]对不同喜树种源的耐盐能力进行了评价并筛选出了耐盐评价指标。在前人研究的基础上,本研究以喜树幼苗为材料,研究在盐碱胁迫下喜树生长及体内离子吸收平衡特性的变化,探讨喜树耐盐碱机理,为开展滨海盐碱地区林木的耐盐碱研究和耐盐碱树种筛选提供理论和技术参考。
1 材料与方法
材料培育:2014年4月中旬将规格一致(苗高(20±5)cm,地径(4.0±0.5)mm)的喜树半同胞家系1年生苗栽植于20 cm(径)×30 cm(高)的塑料盆中,每盆定植2 株,盆中装有10 kg 的干沙土,盆底垫托盘,防止盐分流失。每5 d 透灌1 次1/2Hoagland溶液,常规培养2 个月。本底土壤理化特征为沙土,pH 值7.18,有机质质量分数3.71 g·kg-1,全氮质量分数0.54 g·kg-1,碱解氮质量分数16.04 mg·kg-1,全磷质量分数0.52 g·kg-1,速效磷质量分数2.56 mg·kg-1,全钾质量分数1.10 g·kg-1,速效钾质量分数23.2 mg·kg-1。
盐碱处理:6月中旬在温室中对喜树苗进行盐碱处理。选择生长健壮、长势一致(苗高(50±5)cm,地径(8.0±0.5)mm)的苗木54 盆,随机分成9组,每组6 盆,即每2 盆为一个重复,重复3 次。其中8 组分别用NaCl(50、100 mmol·L-1)、NaHCO3(50、100 mmol·L-1)和Na2SO4(25、50 mmol·L-1)、Na2CO3(25、50 mmol·L-1)的1/2Hoagland 溶液每2 d 浇灌1.0 L 相应浓度溶液1 次,连续进行7 次,此时仅就50、100 mmol·L-1NaCl 两个处理的土壤中NaCl 质量分数(土壤质量以干质量计,同时忽略本底土壤中NaCl)分别为0.205%、0.410%。以后每3 d 称质量补蒸馏水,维持土壤最大田间持水量的70%~80%。另外一组用清水处理作为对照。
指标测定:盐碱处理1 个月后,小心取出每盆中的完整植株,先用自来水冲洗,再用蒸馏水洗净全株,用吸水纸吸去附着水分,分根、茎、叶称取鲜质量。60 ℃烘干至恒质量后称取干质量,粉碎过50目筛备用。
称取干样0.5 g,用HSO4-H2O2法消煮,火焰原子吸收法检测组织中Na+、K+、Ca2+质量分数[12]。按照下列公式计算不同部位对矿质元素选择性运输或吸收(吸收选择性系数RS,X,Na+)能力。
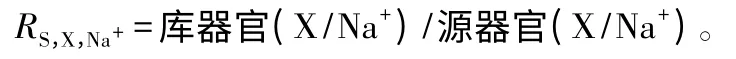
式中:X 代表K+、Ca2+矿质元素;RS,X,Na+值越大,表明源器官控制Na+、促进X 向库器官的运输能力越强,即库器官的选择性运输能力越强[13]。
数据处理及方差分析均用SPSS18.0 完成。文中所有数据均以3 次重复的平均值±标准误差(S.E.)表示。显著水平为5%。
2 结果与分析
2.1 盐碱胁迫对喜树幼苗生物量的影响
如表1所示,盐碱胁迫抑制了喜树根、茎生物量的积累,且随着胁迫强度的增加被进一步抑制;对叶生物量的影响因盐分种类和胁迫强度表现出一定的差异。与对照相比,低浓度中性盐使叶生物量变化不大或略有升高,表明在低强度胁迫下植株能够主动适应盐碱逆境;而在高浓度处理时有所增加,则是植株通过增加叶片的生长来稀释高浓度的Na+来部分适应高盐环境。与对照相比,喜树在低浓度碱性盐胁迫下叶生物量积累变化不大,而在高浓度胁迫下生物量呈现较大幅度地降低,表明高盐、高pH 值环境严重抑制喜树叶生物量积累。总体上说,碱性盐对生物量的影响高于中性盐;盐碱胁迫下喜树通过牺牲喜树根、茎生物量的积累保证叶的生长主动适应盐碱逆境,但在高盐、高pH 值条件下,叶生物量的积累也被不同程度抑制。

表1 盐碱胁迫下喜树幼苗生物量的变化
2.2 盐碱胁迫对喜树幼苗体内Na+质量分数的影响
如表2所示,盐碱胁迫的喜树苗根、茎、叶中的Na+质量分数极显著高于对照(p<0.01)。不同器官的增幅存在差异,由高到低的顺序为茎、叶、根。茎、叶中Na+质量分数显著低于根中的,但盐碱胁迫削弱了这种质量分数差。与对照相比,随着盐碱浓度的增加,喜树根、茎、叶中的Na+质量分数均有不同程度地增加。其中中性盐NaCl 和Na2SO4胁迫对根、茎、叶中Na+质量分数的影响相当,但其影响低于碱性盐NaHCO3和Na2CO3。NaHCO3和Na2CO3相比,Na2CO3使喜树根、茎、叶中的Na+质量分数增加的幅度更大些。

表2 盐碱胁迫下喜树幼苗体内Na+质量分数的变化
2.3 盐碱胁迫对喜树幼苗体内K+、Ca2+质量分数的影响
如表3所示,盐碱胁迫对喜树苗根、茎、叶中的K+质量分数的影响较小(p>0.05)。K+存在由地下器官向地上器官运输的趋势,且盐碱胁迫强度增强这种趋势。与对照相比,低浓度中性盐NaCl(50 mmol·L-1)和Na2SO4(25 mmol·L-1)胁迫下喜树根、茎、叶中的K+质量分数有所增加,而高浓度的中性盐使喜树根中K+质量分数降低,但茎、叶中的K+质量分数有所增加。与对照相比,低浓度的碱性盐NaHCO3使喜树根、茎、叶K+质量分数有所增加,高浓度的NaHCO3降低了喜树中根、茎中的K+质量分数,但叶中K+质量分数有所增加。低浓度Na2CO3使喜树中根的K+质量分数降低,茎、叶中的质量分数增加,高浓度使喜树根、茎、叶中的K+质量分数均降低。
与对照相比,盐碱胁迫使喜树植株体内Ca2+存在由地上器官向地下器官转移的趋势,且盐碱胁迫强度增强这种趋势。NaCl 处理使根、茎叶中Ca2+有所积累,且地下部分积累的幅度较地上部分大;Na2SO4处理使根、茎中Ca2+质量分数增加,而叶中Ca2+降低;NaCl 与Na2SO4对根、茎、叶中Ca2+质量分数的影响随着胁迫强度的增加而加剧。与对照相比,NaHCO3处理使根、茎中Ca2+质量分数增加,而叶中Ca2+降低;低浓度Na2CO3处理(25 mmol·L-1)使根、茎中Ca2+质量分数升高,叶中Ca2+有所降低,而高浓度(50 mmol·L-1)处理使根中Ca2+质量分数升高,但茎、叶中Ca2+质量分数降低;随着胁迫强度的增加,NaCl 与Na2SO4对根、茎、叶中Ca2+质量分数的影响的正效应降低,负效应加剧。
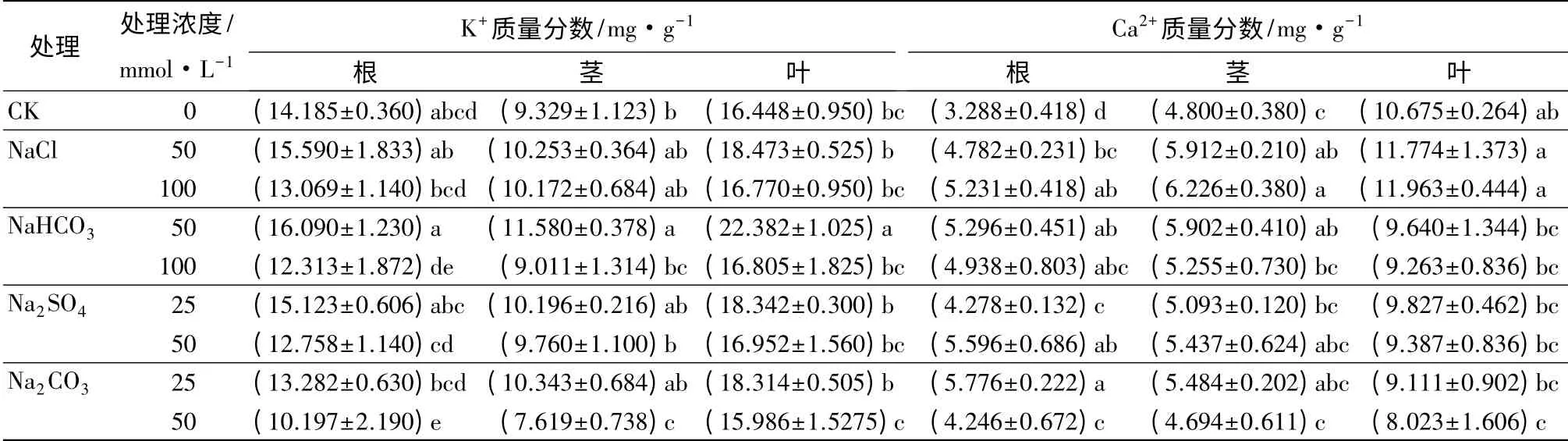
表3 盐碱胁迫下喜树幼苗组织中K+、Ca2+质量分数的变化
2.4 盐碱胁迫对喜树幼苗体内K+质量分数/Na+质量分数、Ca2+质量分数/Na+质量分数的影响
如表4所示,与对照相比,盐碱胁迫降低了根、茎、叶中K+质量分数/Na+质量分数,且随着浓度增加其降低幅度增加。盐碱胁迫对根、茎、叶中K+质量分数/Na+质量分数影响的程度由小到大的顺序为Na2SO4、NaCl、NaHCO3、Na2CO3。4 种盐碱胁迫对根、茎、叶中K+质量分数/Na+质量分数影响的程度由小到大的顺序为根、叶、茎。同样,盐碱胁迫降低了根、茎、叶中Ca2+质量分数/Na+质量分数,且随着浓度增加其降低幅度增加。盐碱胁迫对根、茎、叶中Ca2+质量分数/Na+质量分数影响的程度由小到大的顺序为Na2SO4、NaCl、NaHCO3、Na2CO3。4 种盐碱胁迫对根、茎、叶中Ca2+质量分数/Na+质量分数影响的程度由小到大的顺序为根、叶、茎。
2.5 盐碱胁迫对喜树幼苗选择性运输K+、Ca2+能力的影响
如表5所示,盐碱胁迫改变了喜树幼苗茎、叶选择性运输和吸收K+、Ca2+的能力,具体表现为盐碱胁迫降低了由根向茎选择性运输K+、Ca2+的能力,且总体上随着胁迫强度的增加而加剧,同时增加了叶对K+、Ca2+的选择性吸收,但总体上随着胁迫的增加而降低,但仍高于对照。中性盐NaCl 和NaHCO3对喜树由根向茎选择性运输K+、Ca2+的能力的影响低于碱性盐NaHCO3和Na2CO3的影响,但对由茎向叶中选择运输K+、Ca2+的能力的影响则与之相反。
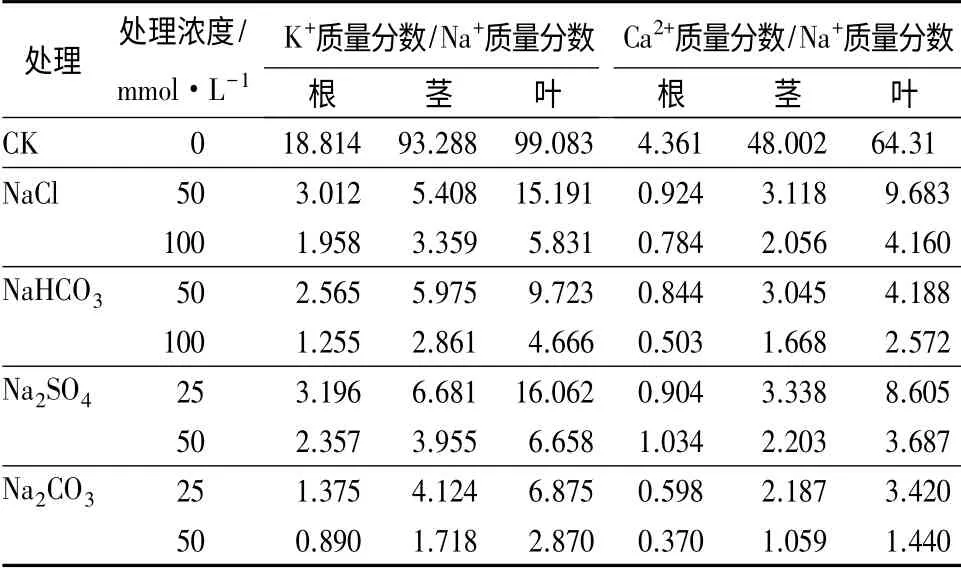
表4 盐碱胁迫对喜树幼苗的K+质量分数/Na+质量分数、Ca2+质量分数/Na+质量分数的影响

表5 盐碱胁迫对喜树幼苗K+、Ca2+选择性运输能力的影响
3 结论与讨论
林木在盐碱环境下生存就要以降低生长为代价[14]。生物量是植物对盐碱胁迫反应的综合表现,也是植物耐盐碱性的直接指标之一。本研究结果表明,盐碱胁迫下喜树通过牺牲喜树根、茎生物量的积累,保证叶生物量的积累的方式主动适应盐碱逆境,即植株通过增加叶片的生长来稀释高浓度的Na+来部分适应盐碱环境;碱性盐对喜树根、茎、叶生物量的影响高于中性盐,表明离子胁迫和高pH 值胁迫下喜树叶生物量的积累也被不同程度抑制。
植物的耐盐性主要取决于根系对盐分的选择吸收及其在器官、组织和细胞3 个层次上的区域化。植物抗盐机制实质上就是Na+及其他离子代谢问题。大多数植物随体内Na+质量分数上升,K+、Ca2+质量分数下降,使细胞内Na+增加和K+外渗,K+质量分数/Na+质量分数、Ca2+质量分数/Na+质量分数比率减小,打破原有的离子平衡,导致K+、Ca2+营养失调,同时因土壤中多量的Na+和碱性盐,使土壤pH 值上升,影响养分的有效性和对养分的吸收。一般认为,在盐胁迫下,植物Na+上升和K+、Ca2+下降的幅度越小,即维持较高的K+质量分数/Na+质量分数、Ca2+质量分数/Na+质量分数,则表明植物对该种盐的耐受能力越强[17]。本研究通过比较不同盐碱胁迫下喜树根茎叶矿质离子分布及选择性运输的变化,证明了不同盐碱逆境下喜树的适应性反应有其相似性,但也存在其特异性反应。盐碱胁迫下喜树通过大量吸收Na+以降低细胞渗透势来维持根系对水分和营养元素的吸收。这与双子叶植物主要以无机离子为渗透调节剂的研究结论是一致的[15]。不同器官Na+质量分数增幅由大到小的顺序为茎、叶、根,这种增幅的变化也与以无机离子为主要渗透调节剂的结论一致。茎木质部主要作为通道运输水分和矿质离子,生理活动相对较弱,所以Na+在此富集则是喜树主动适应盐碱逆境的表现,另外,叶中相对较高的离子浓度,则有利于根中水分和矿质元素的上运。盐碱胁迫处理使喜树体内K+由地下器官向地上器官运输,使Ca2+由地上器官向地下器官转移。随着盐碱浓度和pH 值的升高,K+、Ca2+质量分数沿转移方向呈“升高→降低”的趋势,其中碱性盐对Ca2+质量分数的影响较大,Na2CO3尤甚。盐碱胁迫降低了根、茎、叶中K+质量分数/Na+质量分数、Ca2+质量分数/Na+质量分数,且随着浓度增加其降低幅度增加;4 种盐对其影响的程度由小到大的顺序为Na2SO4、NaCl、NaHCO3、Na2CO3。盐碱胁迫改变了喜树幼苗茎叶选择性运输和吸收K+、Ca2+的能力,具体表现为盐碱胁迫降低了由根向茎选择性运输K+、Ca2+的能力,同时增加了叶对K+、Ca2+的选择性吸收,表明喜树对K+、Ca2+的由茎到叶的选择性吸收和运输能力较强,从而维持叶中相对较好的离子平衡。根系环境的高pH 值直接降低植物必需矿质元素的游离度和活度,造成钙、镁等元素的大量沉淀[16],致使根系周围离子供应严重失衡。从根际营养角度来看,pH 值上升,必然会影响根际大部分营养元素的有效性,最终扰乱植物正常生理代谢。所以,在以NaHCO3、Na2CO3为主的碱化地上,采用降低pH 值的中和措施无疑是一种有效的治理途径。
综上所述,喜树在低强度的盐碱胁迫下通过牺牲根、茎生物量积累维持叶的生长来稀释叶中迅速升高的Na+浓度,同时喜树通过吸收Na+降低渗透势维持对水分和矿质营养的吸收,另外,通过由根向叶运输K+部分消除叶中大量积累的Na+毒害,又通过由叶向根中运输Ca2+部分消除根中高浓度Na+的毒害,表现出一定的主动适应性。但长期的盐碱胁迫必然造成叶中Ca2+和根中K+的显著降低及Na+毒害,致使矿质营养的失衡和匮乏。碱胁迫对喜树的影响甚于盐胁迫。盐胁迫主要涉及渗透胁迫和离子毒害[18-19],碱胁迫除涉及以上因素还涉及高pH 值胁迫,要适应碱胁迫必然要消耗更多的物质和能量,因此,生长抑制更严重[15]。
[1] Munns R.Comparative physiology of salt and water stress[J].Plant,Cell and Environment,2002,25:239-250.
[2] Shi Decheng,Wang Deli.Effects of various salt-alkaline mixed stresses on Aneurolepidium chinense (Trin.)Kitag[J].Plant and Soil,2005,271(1):15-26.
[3] 刘小京,李向军,陈丽娜,等.盐碱区适应性农作制度与技术探讨以河北省滨海平原盐碱区为例[J].中国生态农业学报,2010,18(4):911-913.
[4] Shi Decheng,Sheng Yanmin.Effect of various salt-alkaline mixed stress conditions on sunflower seedlings and analysis of their stress factors[J].Environmental and Experimental Botany,2005,54(1):8-21.
[5] 颜宏,石德成,尹尚军,等.盐碱胁迫对羊草体内N 及几种有机代谢产物积累的影响[J].东北师大学报:自然科学版,2000,32(3):47-52.
[6] 周婵,张卓,杨允菲.实验羊草种群幼苗对不同梯度盐碱胁迫的生理响应[J].东北师大学报:自然科学版,2003,35(4):62-67.
[7] Shi Decheng,Yin Lijuan.Difference between salt (NaCl)and alkaline (Na2CO3)stresses on Puccinellia tenuiflora (Griseb.)Scribn.et Merr.plants[J].Acta Botanica Sinica,1993,35(2):144-149.
[8] 刘慧娟.NaCl 胁迫下喜树组培苗生长及生理生化特性研究[D].临安:浙江农林大学,2013.
[9] 郝峰鸽,周俊国,周秀梅.NaCl 胁迫对喜树幼苗生长和叶片生理特性的影响[J].东北林业大学学报,2010,38(1):18-19.
[10] 郝峰鸽,周凯,杨立峰,等.NaCl 胁迫对喜树幼苗生长和光合特性的影响[J].福建林学院学报,2009,29(3):247-251.
[11] 张露婷,吴江,梅丽,等.喜树种源耐盐能力评价及耐盐指标筛选[J].林业科学,2011,47(11):66-72.
[12] 王保山,赵可夫.小麦叶片中Na、K 提取方法比较[J].植物生理学通讯,1995,31(1):50-52.
[13] 杨敏生,李艳华,梁海永,等.盐胁迫下白杨无性系苗木体内离子分配及比较[J].生态学报,2003,23(2):271-277.
[14] 刘志华,赵可夫.盐胁迫对獐茅生长及Na+和K+含量的影响[J].植物生理与分子生物学学报,2005,31(3):311-316.
[15] 赵可夫,冯立田,卢元芳,等.九龙江口秋茄和白骨壤的渗透调节剂及其贡献[J].海洋与湖沼,1999,30(1):58-61.
[16] 石德成,赵可夫.NaCl 和Na2CO3对星星草生长及营养液中主要矿质元素存在状态的影响[J].草业学报,1997,6(2):51-61.
[17] Parida A K,Das A B.Salt tolerance and salinity effects on plants:A review[J].Ecotoxicology and Environmental Safety,2005,60(3):324-349.
[18] Moghaieb R E A,Saneoka H,Fujita K.Effect of salinity on osmotic adjustment,glycinebetaine accumulation and the betaine aldehyde dehydrogenase gene expression in two halophytic plants,Salicornia europaea and Suaeda maritime[J].Plant Science,2004,166:1345-1349.
[19] 李长有,金昌民,倪福太,等.盐碱胁迫对抗碱盐生植物碱地肤生长特性的影响[J].吉林师范大学学报:自然科学版,2008,29(3):43-45,50.
猜你喜欢
金桥(2021年11期)2021-11-20
现代畜牧科技(2021年4期)2021-07-21
第一财经(2019年8期)2019-08-26
中成药(2018年4期)2018-04-26
西藏科技(2016年8期)2016-09-26
中成药(2016年4期)2016-05-17
现代食品(2016年14期)2016-04-28
中国麻业科学(2015年5期)2015-12-28
哈尔滨医药(2015年2期)2015-12-01
西藏科技(2015年10期)2015-09-26
