
腹毛类纤毛虫
——杆形戴维虫的形态学和细胞发生
2015-03-01 09:41:14李凤超李延博吕昭梅玉明高素伟邵
水生生物学报 2015年6期
李凤超李延博吕 昭梅玉明高素伟邵 晨
(1. 河北大学生命科学学院, 河北省动物系统学与应用重点实验室, 保定 071002; 2. 中国海洋大学海洋生物多样性与进化研究所, 青岛 266003; 3. 西安交通大学生命科学与技术学院, 生物医学信息工程教育部重点实验室, 西安 710049)
研究简报
腹毛类纤毛虫
——杆形戴维虫的形态学和细胞发生
李凤超1, 2李延博1吕 昭3梅玉明1高素伟1邵 晨3
(1. 河北大学生命科学学院, 河北省动物系统学与应用重点实验室, 保定 071002; 2. 中国海洋大学海洋生物多样性与进化研究所, 青岛 266003; 3. 西安交通大学生命科学与技术学院, 生物医学信息工程教育部重点实验室, 西安 710049)
腹毛类纤毛虫广泛见于海、淡水及土壤等各种生境中[1—5]。在生态系统中, 腹毛目纤毛虫种类多、数量大、食性广, 在物质循环和能量流动过程中扮演重要的角色[6—8]。腹毛类纤毛虫结构复杂、形态多样, 分类鉴定困难, 而其细胞发生学研究, 不仅可为腹毛类纤毛虫的分类学和系统学提供重要依据, 同时也可为细胞生物学、遗传学等领域提供参考资料[9—12]。在土壤中纤毛虫种类丰富, 据估计至少有 2000种[13]; 在已发现的约 1000种土壤纤毛虫中, 1/4—1/3的种类为腹毛类[14]。
戴维虫属(Deviata)由Eigner于1995年建立[15], 隶属于腹毛目, 目前其科级分类地位不明。本属已见报道的有9种[15—23], 即收缩戴维虫 Deviata abbrevescens、杆形戴维虫D. bacilliformis、巴西戴维虫D. brasiliensis、埃氏戴维虫D. estevesi、四核戴维虫D. quadrinucleata、 罗氏戴维虫D. rositae、多毛戴维虫D. polycirrata、旋口戴维虫D. spirostoma和拟杆形戴维虫D. parabacilliformis。杆形戴维虫D. bacilliformis种群在以色列、非洲及阿根廷相继被发现[17—19], 但其细胞发生过程尚未见详细报道。
在2012年11月自天津郊区滨保高速小韩庄服务区及其周边的一次采样中, 获得了拟杆形戴维虫 Deviata parabacilliformis和杆形戴维虫Deviata bacilliformis两个种, 并对拟杆形戴维虫的形态发生以及两个种的 18S rDNA进行了研究。为积累和补充戴维虫属的形态发生学资料, 本文进一步对杆形戴维虫的细胞发生过程进行了详细报道。
1 材料与方法
土壤样品采自天津市郊滨保高速(G2501)小韩庄服务区内的草坪, 为沙质土壤, 带回实验室风干后, 采用“非浸没培养皿法”培养虫体, 加入大米粒以富集细菌, 于室温下(约25 )℃建立了纯培养。虫体分离、培养、观察、染色制片等参照宋微波等[24]; 相关分类术语参考Berger[16]。
2 结果
2.1 杆形戴维虫的形态特征(图版Ⅰ-1—6)
虫体呈长圆柱形, 前后两端略尖, 活体大小约(70—160) µm× (20—55) µm, 平均体长 135 μm, 体宽30 μm。体内部充满(1—3) µm的内质颗粒, 呈暗灰色(图版Ⅰ-1—3)。刚分裂的个体体形较短(图版Ⅰ-4)。伸缩泡位于虫体中部偏左缘, 未见收集管。皮层无色, 不具皮层颗粒。口区约占体长的 1/6—1/4, 口围带由 16—28片小膜组成。额棘毛3根, 口棘毛1根, 口侧棘毛1根。额腹棘毛6列, 右缘棘毛3列, 左缘棘毛4列(其中最外侧的两列前端分别有两根触毛), 背触毛 1列。无横棘毛和尾棘毛(图版Ⅰ-5、6)。大核2—4枚, 通常2枚, 小核2—6枚。体棘毛长约(10—15) µm, 背触毛长约5 µm。额棘毛通常由9个毛基粒组成, 口棘毛和口侧棘毛由9个毛基粒组成,缘棘毛、2列长额腹棘毛通常由 2个基粒组成(其中每列的前1—3根棘毛由4个毛基粒组成)。运动方式为附底爬行, 偶尔旋游于水中, 并绕身体长轴不断翻转。
2.2 细胞发生过程(图版Ⅰ-7—13、图版Ⅱ)
口器发生 杆形戴维虫的口原基为独立发生, 发生过程中没有老结构的参与。最初, 位于第V列额腹棘毛列和第 1列左缘棘毛列之间的裸毛斑增殖, 形成一些无序排列的毛基粒群(图版Ⅰ-7, 图版Ⅱ-14、15)。随后毛基粒不断增殖, 形成后仔虫的口原基(图版Ⅰ-8, 图版Ⅱ-16、17)。随着虫体的进一步发育, 该原基向后延伸、增大, 在右前端逐渐形成新的小膜, 逐渐出现条带(图版Ⅰ-8)。口原基继续由前向后组装成规整排列的小膜, 并最终形成后仔虫的新口围带(图版Ⅰ-9)。在整个过程中老口围带被完全保留, 成为前仔虫的口围带。
腹面体棘毛的发生 第I—III列额腹棘毛原基: 在细胞分裂中期, 老口侧膜(PM)前端反分化形成前仔虫的第I列额腹棘毛原基, 口棘毛和口侧棘毛反分化分别形成前仔虫的第 II和第 III列额腹棘毛原基(图版Ⅰ-9、图版Ⅱ-19); 后仔虫的第I—III列额腹棘毛原基均来源于口原基(图版Ⅰ-9、图版Ⅱ-19)。
第IV列额腹棘毛原基: 前仔虫的第IV列原基来源于老结构的第IV列额腹棘毛列; 而后仔虫的第IV列原基则来源于老结构的第V列额腹棘毛列(图版Ⅰ-10)。
第V列额腹棘毛原基: 前仔虫第V列原基来源不明(图版Ⅰ-10、图版Ⅱ-24), 推测来源于第VI列额腹棘毛列,后仔虫的第V列原基为独立发生(图版Ⅰ-9, 图版Ⅱ-19、22)。
第VI列额腹棘毛原基: 前后仔虫的第VI列额腹棘毛原基起始于老结构中部的一个棘毛反分化形成的原基(图版Ⅰ-8、图版Ⅱ-17), 这个原基向两端延伸(图版Ⅱ-18),在分化中期从中间断开后, 形成前后仔虫的第VI列额腹棘毛原基。
最终, 由第I列额腹棘毛原基形成新虫体的最左侧的额棘毛和波动膜, 第 II列额腹棘毛原基形成的中间的额棘毛和口棘毛, 第III列额腹棘毛原基贡献最右侧的额棘毛和口侧棘毛(图版Ⅱ-21、23), 第IV—VI列额腹棘毛原基分别形成第IV-VI列额腹棘毛。
缘棘毛发生 缘棘毛原基均源自于老结构。首先在左、右缘棘毛内部出现后仔虫的缘棘毛原基(图版Ⅰ-9、图版Ⅱ-19), 随后出现前仔虫的缘棘毛原基。随着原基的发育, 将会向两端延伸, 新生的缘棘毛列逐渐取代老结构(图版Ⅰ-12、图版Ⅱ-25)。
背触毛发生 形态发生中早期, 在背触毛列的前后各 1/3 处出现了背触毛原基(图20)。随着发生的进行,原基中的毛基粒不断增多, 并向两极延伸。新生背触毛列逐渐向细胞两端延伸, 取代老结构(图版Ⅰ-11、13, 图版Ⅱ-26)。
核器的发生 核器演化遵循一般过程。在中期阶段, 大核融合为椭球形, 随后伴随前后仔虫的相互分离而再次分裂(图版Ⅰ-11、13, 图版Ⅱ-27)。小核的发生过程不明。
实验中发现一个重组的个体, 在第VI列额腹棘毛原基的右侧出现一列原基(图版Ⅱ-28)
3 讨论
3.1 杆形戴维虫天津种群与其他种群的比较在已报道的杆形戴维虫种群中, 以色列和阿根廷两个种群描述最为详细, 故本文主要与上述两个种群比较。与以色列和阿根廷种群相比, 本种群的纤毛图式特征与之完全吻合, 各项指标统计数据的波动范围略有差异(表1)。以色列和阿根廷种群都具有收集管, 而本种群不具收集管, 这是与上述两个种群的主要差异。同时本种群和阿根廷种群一样, 最外缘的两列右缘棘毛列前端各具两根刚毛(图6)。阿根廷种群大核总是2枚, 而天津种群和以色列种群大核2—4枚, 通常2枚。
3.2 与戴维虫属其他种发生特征的比较杆形戴维虫的主要发生特征可归纳如下: 后仔虫口原基来源于第V列额腹棘毛列和第1列左缘棘毛之间的裸毛斑, 口原基的发生过程中没有棘毛的参与; 老口围带完全保留; 额腹棘毛为6列发生式, 老结构的第VI列额腹棘毛列中的一个棘毛反分化成原基, 最终形成前后仔虫的第VI列额腹棘毛原基; 背触毛、缘棘毛原基产生于老结构中; 大核在发生中期完全融合。在已报道的9种戴维虫中, 6个种的个体发生过程已有报道, 即模式种收缩戴维虫, 杆形戴维虫阿根廷种群, 巴西戴维虫, 多毛戴维虫, 埃氏戴维虫和拟杆形戴维虫[15—23]。据Küppers & Claps报道[19], 杆形戴维虫阿根廷种群的口原基来源于第V列额腹棘毛列和第1列左缘棘毛列之间的 1—3对裸毛斑, 在口原基发生的过程中, 没有老结构的参与, 这与本种群相一致。而收缩戴维虫和巴西戴维虫的口原基来源于老的第IV列额腹棘毛列。杆形戴维虫与拟杆形戴维虫的形态发生过程极为相似: 均为 6列原基发生式, 6列额腹棘毛原基的来源和贡献基本相同, 后仔虫口原基的形成均没有老结构的参与, 缘棘毛、背触毛发生过程相同, 为常见的两组发生式; 不同之处在于口原基的来源, 杆形戴维虫为独立发生, 而拟杆形戴维虫的口原基来源于腹面中部的裸毛斑。

表1 不同杆形戴维虫种群间重要形态特征的比较Tab. 1 Comparison of Tianjin population of Deviata bacilliformis with other two populations
[1] Jiang J M, Ma H G, Shao C. Morphology and morphogenesis of Sterkiella histriomuscorum (Ciliophora, Hypotricha) [J]. Acta Hydrobiologica Sinica, 2013, 37(2): 227—234 [姜佳枚,马洪钢, 邵晨. 变藓棘毛虫的形态学重描述及细胞发生学研究. 水生生物学报, 2013, 37(2): 227—234]
[2] Chen L Y, Liu W W, Liu A, et al. Morphology and molecular phylogeny of a new marine hypotrichous ciliate, Hypotrichidium paraconicum n. sp. (Ciliophora, Hypotrichia) [J]. Journal of Eukaryotic Microbiology, 2013, 60(6): 588—600
[3] Chen X M, Miao M, Ma H G, et al. Morphology, morphogenesis and small subunit (SSU) rRNA gene sequence of the new brackish water ciliate Strongylidium orientale sp. nov. (Ciliophora, Stichotrichia) from Hong Kong, southern China [J]. International Journal of Systematic and Evolutionary Microbiology 2013, 63(3): 1155—1164
[4] Jiang J M, Xing Y, Miao M, et al. Two new marine ciliates, Caryotricha rariseta n. sp. and Discocephalus pararotatorius n. sp. (Ciliophora, Spirotrichea), with phylogenetic analyses inferred from the small-subunit rRNA gene sequences [J]. Journal of Eukaryotic Microbiology, 2013, 60(4): 388—398
[5] Shao C, Li L Q, Zhang Q Q, et al. Molecular phylogeny and ontogeny of a new ciliate genus, Paracladotricha salina n. g., n. sp. (Ciliophora, Hypotrichia) [J]. Journal of Eukaryotic Microbiology, 2014, 61(4): 371—380
[6] Dai R H, Xu K D, He Y Y. Morphological, physiological, and molecular evidences suggest that Euplotes parawoodruffi is a junior synonym of Euplotes woodruffi (Ciliophora, Euplotida) [J]. Journal of Eukaryotic Microbiology, 2013, 60(1): 70—78
[7] Fan Y B, Chen X M, Hu X Z, et al. Morphology and morphogenesis of Apoholosticha sinica n. g., n. sp. (Ciliophora, Hypotrichia), with consideration of its systematic position among urostylids [J]. European Journal of Protistology, 2014, 50(1): 78—88
[8] Shao C, Lü Z, Jin L, et al. Morphogenesis of a unique pseudourostylid ciliate, Trichototaxis songi (Ciliophora, Urostylida) [J]. European Journal of Protistology, 2014, 50(1): 68—77
[9] Li F C, Xing Y, Li J M, et al. Morphology, morphogenesis and small subunit rRNA gene sequence of a soil hypotrichous Ciliate, Perisincirra paucicirrata (Ciliophora, Kahliellidae), from the shoreline of the Yellow River, north China [J]. Journal of Eukaryotic Microbiology, 2013, 60(3): 247—256
[10] Modeo L, Petroni G, Lobban C S, et al. Morphological, ultrastructural, and molecular characterization of Euplotidium rosati n. sp. (Ciliophora, Euplotida) from Guam [J]. Journal of Eukaryotic Microbiology, 2013, 60(1): 25—36
[11] Park K M, Jung J H, Min G S. Morphology, morphogenesis, and molecular phylogeny of Anteholosticha multicirrata n. sp. (Ciliophora, Spirotrichea) with a note on morphogenesis of A. pulchra (Kahl, 1932) Berger, 2003 [J]. Journal of Eukaryotic Microbiology, 2013, 60(6): 564—577
[12] Singh J, Kamra K, Sapra G R. Morphology, ontogenesis, and molecular phylogeny of an Indian population of Cyrtohymena (Cyrtohymenides) shii, including remarks on the subgenus [J]. European Journal of Protistology, 2013, 49(2): 283—297
[13] Chao A, Li P C, Agatha S, et al. A statistical approach to estimate soil ciliate diversity and distribution based on data from five continents [J]. Oikos, 2006, 114(3): 479—493
[14] Foissner W, Agatha S, Berger H. Soil ciliates (Protozoa, Ciliophora) from Namibia (Southwest Africa), with emphasis on two contrasting environments, the Etosha region and the Namib Desert [J]. Denisia, 2002, 5: 1—1459
[15] Eigner P. Divisional morphogenesis in Deviata abbrevescens nov. gen., nov. spec., Neogeneia hortualis nov. gen., nov. spec., and Kahliella simplex (Horváth) Corliss andredefinition of the Kahliellidae (Ciliophora, Hypotrichida) [J]. European Journal of Protistology, 1995, 31(3): 341—366
[16] Berger H. Monograph of the Gonostomatidae and Kahliellidae (Ciliophora, Hypotricha) (Monographiae Biologicae, vol. 90) [M]. Springer, Berlin. 2011, 1—742
[17] Berger H, Foissner W. Morphology and biometry of some soil hypotrichs (Protozoa: Ciliophora) [J]. Zoologisches Jahrbuch, Systematik, 1987, 114(2): 193—239
[18] Dragesco J. Infaciliature et morphometrie de vingt espèces de ciliés hypotriches recoltés au Rwanda et Burundi, comprenant Kahliella quadrinucleata n. sp. Pleurotricha multinucleata n. sp. et Laurentiella bergeri n. sp. [J]. Travaux du Museum d'Histoire Naturelle “Grigore Antipa”, 2003, 45(1): 7—59
[19] Küppers G C, Claps M C. Morphology and notes on morphogenesis during cell division of Deviata polycirrata n. sp. and of Deviata bacilliformis (Ciliophora: Kahliellidae) from Argentina [J]. Journal of Eukaryotic Microbiology, 2010, 57(3): 273—284
[20] Küppers G C, Lopretto E C, Claps M C. Description of Deviata rositae n. sp., a new ciliate species (Ciliophora, Stichotrichia) from Argentina [J]. Journal of Eukaryotic Microbiology, 2007, 54(5): 443—447
[21] Paiva T S, Silva-Neto Idda. Deviata estevesi sp. n. (Ciliophora: Spirotrichea), a new ciliate protist from a restinga lagoon in Rio de Janeiro, Brazil [J]. Acta Protozoologica, 2005, 44(4): 351—362
[22] Li F C, Lü Z, Yi Z Z, et al. Taxonomy and phylogeny of two Deviata-species (Protista, Ciliophora) from China, with description of a new soil form, D. parabacilliformis spec. nov [J]. International Journal of Systematic and Evolutionary Microbiology, 2014, 64(11): 3775—3785
[23] Siqueira-Castro ICV, Paiva T S, Silva-Neto Idda. Morphology of Parastrongylidium estevesi comb. nov. and Deviata brasiliensis sp. nov. (Ciliophora: Stichotrichia) from a sewage treatment plant in Rio de Janeiro, Brazil [J]. Zoologia, 2009, 26(4): 774—786
[24] Song W B, Xu K D, Shi X L, et al. Progress in Protozoology [M]. Qingdao: Qingdao Ocean University Press. 1999, 362[宋微波, 徐奎栋, 施心路, 等. 原生动物学专论. 青岛:青岛海洋大学出版社. 1999, 362]
ON MORPHOLOGY AND MORPHOGENESIS OF A SOIL HYPOTRICHOUS CILIATE, DEVIATA BACILLIFORMIS (GELEI, 1954) EIGNER, 1995 (PROTOZOA, CILIOPHORA)
LI Feng-Chao1, 2, LI Yan-Bo1, LÜ Zhao3, MEI Yu-Ming1, GAO Su-Wei1and SHAO Chen3
(1. The key Laboratory of Zoological Systematics and Application, College of Life Sciences, Hebei University, Baoding 071002, China; 2. Laboratory of Protozoology, Institute of Evolution & Marine Biodiversity, Ocean University of China, Qingdao 266003, China; 3. The Key Laboratory of Biomedical Information Engineering, Ministry of Education, Xi’an Jiaotong University, Xi’an 710049, China)
腹毛类纤毛虫; 纤毛图式; 细胞发生; 原生动物; 戴维虫
Hypotrichous ciliate; Infraciliature; Morphogenesis; Protozoa; Deviata
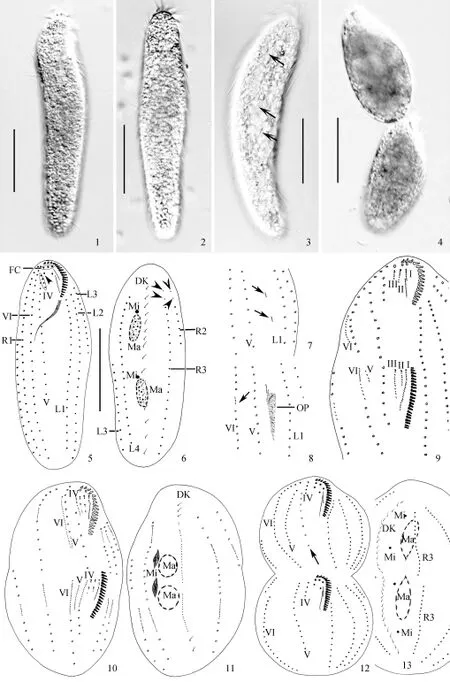
图版 Ⅰ 杆形戴维虫的活体形态、纤毛图式及发生过程PlateⅠ Morphology and morphogenesis of Deviata bacilliformis

图版 Ⅱ 杆形戴维虫发生时期蛋白银染后的显微照片PlateⅡ Micrographs of morphogenetic stages of Deviata bacilliformis after protargol impregnation
Q959.11
A
1000-3207(2015)06-1255-06
10.7541/2015.164
2014-08-28;
2014-12-20
国家自然科学基金(31372148, 31172063); 河北省重点实验室运行补助费(14967611D); 河北省教育厅重点项目(2D2015038)资助
李凤超 (1970—), 男, 河北乐亭人; 博士; 研究方向为原生动物学。E-mail: lifengchao2000@sina.com
邵晨, E-mail: shaochen@mail.xjtu.edu.cn
猜你喜欢
生态学报(2022年9期)2022-05-26 10:03:04
计算机应用(2018年10期)2018-11-22 09:37:54
渔业科学进展(2018年1期)2018-04-03 08:45:38
华南师范大学学报(自然科学版)(2017年4期)2017-09-11 09:16:23
浙江大学学报(农业与生命科学版)(2017年2期)2017-05-19 07:41:15
海洋与湖沼(2016年1期)2016-01-15 03:52:09
西藏科技(2015年6期)2015-09-26 12:12:08
地球学报(2015年5期)2015-06-06 10:45:11
地球学报(2015年5期)2015-06-06 10:45:11
湖北农业科学(2014年6期)2014-07-02 11:16:14
