
小江拟尖头鲌的年龄、生长、繁殖及其资源开发状况研究
2015-02-28 08:28段鹏翔杨志唐会元肖琼龚云肖调义刘小燕
水生生物学报 2015年4期
段鹏翔杨 志唐会元肖 琼龚 云肖调义刘小燕
(1. 湖南农业大学动物科学技术学院, 长沙 410000; 2. 水利部中国科学院水工程生态研究所, 水利部水工程生态效应与生态修复重点实验室, 武汉 430079)
小江拟尖头鲌的年龄、生长、繁殖及其资源开发状况研究
段鹏翔1杨 志2唐会元2肖 琼1龚 云2肖调义1刘小燕1
(1. 湖南农业大学动物科学技术学院, 长沙 410000; 2. 水利部中国科学院水工程生态研究所, 水利部水工程生态效应与生态修复重点实验室, 武汉 430079)
拟尖头鲌(Culter oxycephaloides)属鲤形目(Cypriniformes), 鲤科(Cyprinidae), 鲌亚科(Cultrinae), 鲌属(Culter), 为中国特有鱼类, 分布于长江流域的四川、湖北、湖南等省的江河湖泊中。2013年 4—7月以及10—12月在三峡库区支流小江江段收集拟尖头鲌样本1741尾, 对其年龄、生长、繁殖及其资源开发状况进行了研究。结果表明: 所采集的拟尖头 鲌包括 5个年龄组, 其中 2—3龄年龄组为优势年龄组, 占 78.59% (N=425尾); 体长与体重呈幂函数关系, 表达式为W = 0.000007L3.10(R2=0.99, N=789尾), 其生长属于匀速生长类型; 采用 von Bertalanffy生长方程分别拟合体长、体重与年龄的关系, 其表达式分别为: Lt=519.63[1–e–0.22(t+0.12)], Wt=1806.08[1–e–0.22(t+0.12)]3.10; 生长拐点年龄t=5.02龄, 此时对应的体长348 mm, 体重558.8 g; 绝对繁殖力变动范围为20382—88430粒/尾, 相对繁殖力变动范围为54—161粒/g, 体重与绝对繁殖力的关系以二次方程曲线拟合度最优, 其表达式为 Fabs=33462.08+0.11W–44.01W2(R2=0.49, sig.=0.02<0.05, N=31); Ⅳ卵巢卵径变动范围为0.0675—0.1325 cm, 平均值为0.1060 cm; 拟尖头 鲌产卵类型为分批产卵类型; 小江拟尖头鲌的现有开发率为E=0.51/年; Beverton-Holt动态综合模型显示, 目前小江拟尖头 鲌的Emax为0.40/年, E10为0.32/年, E50为0.25/年。根据研究, 目前小江拟尖头鲌的种群处于过度开发状况,应采取措施, 加强其资源保护。
小江; 拟尖头鲌; 年龄与生长; 繁殖; 资源开发
拟尖头鲌(Culter oxycephaloides)属鲤形目(Cypriniformes), 鲤 科 (Cyprinidae), 鲌 亚 科(Cultrinae), 鲌属(Culter), 为中国特有鱼类, 分布于黑龙江和长江流域的四川、重庆、湖北、湖南等省的江河湖泊中[1—3], 其食性为鱼虾等[4]。据《四川鱼类志》(1994)记载[5], 长江上游干流万县(今重庆万州区)下游附近有该种鱼类分布, 但数量不多。
小江又名彭溪河, 是三峡腹地北岸的一级支流,源于大巴山麓, 流经开县、云阳进入长江, 是三峡水库重庆区域万州以下水域中(除长江外)流域面积最大的一条河流, 小江干流全长约182 km, 河道平均坡降3.7%, 流域面积5172.5 m2[6]。在三峡水库蓄水后, 小江水文情势呈周期性变化趋势, 其中三峡低水位运行时, 小江养鹿以上江段保持一定的流水生境, 而在三峡高水位运行时, 整个小江几乎保持宽水面的静、缓流水状态[7]。小江水动力学的改变[8]以及大量陆地营养物质的输入[9], 导致小江水体的小型鱼类数量明显增多[10]鲌。小型鱼类是 属鱼类的重要饵料资源, 鲌饵料丰度的升高促进了 属鱼类资源量的上升[10], 鲌其中的拟尖头 在2013—2014年小江渔获物中的数量也明显增加, 已成为小江主要的经济鱼类之一。目前, 鲌关于拟尖头 的基础生物学及种群资源开发现状等研究, 在国内外仅有零星报道[5], 鲌而对小江区域拟尖头 的相关研究则在国内外尚未见报道。本研究于2013年4—7月及10—12月对小江水域拟尖头鲌进行了样本采集, 并对该区鲌域内拟尖头 的年龄、生长、繁殖以及种群资源进行了研究, 鲌以期了解小江拟尖头 的年龄、生长、繁殖以及种群资源开发现状, 鲌旨在为小江拟尖头的资源保护和合理利用提供科学依据。
1 材料与方法
1.1 样本采集
2013年4—7月以及10—12月在三峡库区支流小江的双江、黄石、高阳、养鹿和渠口江段逐月进行样本收集(图 1), 采集网具为三层刺网, 网目大小5—12 cm, 鲌共计收集拟尖头 样本1741尾, 对所有样本的全长、体长和体重进行测量, 全长、体长精确到1 mm, 体重精确到0.1 g, 并在背鳍起点下方与侧线中间摘取5—10 鲌枚鳞片作为拟尖头 年龄鉴定的材料[11], 共采集到年龄材料 425尾。同时, 选择部分体型较大、具有明显第二性征的个体进行解剖,参考殷名称(1995)的方法鉴定性别和性腺发育期[11],并在 IV期卵巢前、中、后部累计取约 5 g样本, 在5%的福尔马林中固定后带回实验室处理。
1.2 样品处理
年龄鉴定 用清水冲洗鳞片表面胶质及污物,然后置于载玻片上进行固定。采用 SZ61TR体视显微镜进行观察, 并用YM200数码成相系统软件保存粦图像。本研究参照邓中 等的方法[12]进行年龄计数和确认, 以元月 1日为年龄递增日期划分年龄组。如已有年轮数为 a, 若在第 a个年轮的外侧新年轮形成后外缘有一部分环片出现, 将年龄计为 a+; 若在第a个年轮的外侧新年轮尚未完全形成但已出现较少密带的个体则年龄计为(a+1), 即 0+—1龄为 1 龄, 1+—2龄为2龄, 依此类推。年龄鉴定时3人分别鉴定同一样本, 当鉴定结果不一致时, 经查阅相关文献讨论后确定。
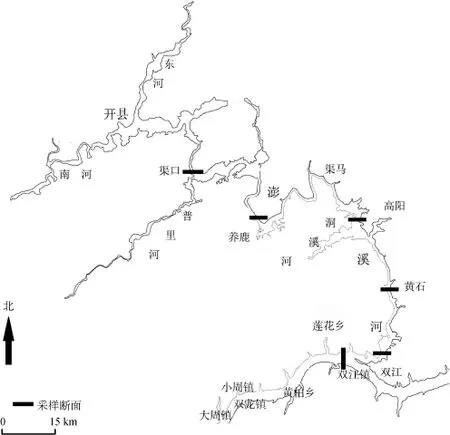
图1 2013年小江采样江段示意图Fig. 1 Schematic diagram of the sampled sections of the Xiao River in 2013
性腺样本处理 对卵巢样品用清水漂洗数次,再取出放在培养皿中, 轻轻揉擦, 使卵粒完全脱离卵巢系膜(卵巢板), 然后去掉系膜, 并滤去清水, 将全部卵子移入搪瓷盘内。将部分卵粒排成一直线:卵粒间保持一定的间距, 每堆卵粒不超过 3粒, 用计数器计数。同时选择卵粒饱满的卵巢Ⅳ期, 从卵巢的前、中、后三个部分随机测量 150粒卵粒, 用基于台测微尺校正的目标微尺在显微镜下测定其卵径, 具体方法为: 将台测微尺和所需测定的卵放置在一个平面上, 然后用以标定每一格目测微尺表示的实际长度单位, 如目测微尺10大格, 每大格5等分, 计 50小格, 用台测微尺标定(放大 20倍时)为5 mm, 则1小格为0.1 mm, 半格为0.05 mm。然后,可用目测微尺直接测卵径[11]。
1.3 数据分析
体长-体重关系 采用幂函数方程拟合拟尖鲌头 的体长-体重关系: W = aLb式中W 为体重(g), L 为体长(mm) , a、b 为关系系数[11]。采用配对样本的t检验检验拟合的b值与期望值3.0的差异[12]。同时采用残差平方和(ARSS)方法进行差异性检验, 比较雌雄个体体长-体重关系式的差异[13]。
生长方程 采用最小二乘法(Least square method, LSQ)拟 合 Von Bertalanffy 方 程 :

式中: Lt(mm) 和Wt(g) 分别表示t 龄时的体长和体重, L∞和 W∞分别表示渐进体长和体重, k 为生长系数, t0为理论生长起点年龄, t 表示时间(以年为单位)[14]。
死亡特征与开发率 分别采用 Pauly(1990)的方法[15]以及 Pauly(1980)的经验公式[16]计算总死亡系数 Z 和自然死亡系数M:

式中: Z为总死亡系数; N为每一个体长组的尾数; ∆t为对应体长组上限经体长转换年龄后的年龄减去该体长组下限经体长转换年龄后的年龄; a为拟合截距; b为拟合方程的斜率, 其绝对值即为Z; ML为基于体长的自然死亡系数; MW为基于体重的自然死亡系数; L∞为渐近体长; W∞为渐近体重; K为生长系数; T为区域年均水温, , ℃ 采用对小江周年采样的结果21.04 ; ℃ M为自然死亡系数。
捕捞死亡系数 F=Z–M; 总死亡率 a=1–exp(–z);自然死亡率 v=a×M/Z; 捕捞死亡率 u=a–v; 开发率E=F/Z[11]。
绝对和相对繁殖怀卵量 绝对怀卵量 Fabs=(Ns/Ws)×Wo和相对怀卵量 Frel=Fabs/Wb, 式中: Ns为样品中沉积卵黄的卵粒数, Ws为样品重量, Wo为卵巢重量, Wb为鱼体空壳重[11]。采用线性模型、对数
模型、2次方程、3次方程、复合模型、幂函数、指数模型7种, 对体重与绝对繁殖力之间的关系进行拟合, 以判定系数(R2)的大小确定拟合模型的优劣[12]。相对单位补充渔获量和生物量 利用
FISATⅡ软件中的刀刃式选择假设模型建立相对单位补充鱼产量和相对单位补充生物量曲线, 该模型基于Beverton和Holt稳定状态模型, 假定捕捞格局
(主要指捕捞强度和开捕年龄)长时间不变, 所有个体一经补充均有被捕的可能。最适捕捞规格可通过设定鱼类资源最适开发率, 将自然死亡系数和生长系数代入推算得出, 具体如下[17]:

式中: Y′/R为相对单位补充鱼产量; B′/R为相对单位补充生物量; E为开发率; L∞为渐近体长; Lc为起捕体长; M为自然死亡系数; Z为总死亡系数; K为生长系数。
根据相对单位补充渔获量模型计算3种不同水平的开发率: (1)Emax: 获得最大渔获量的开发率; (2) E10: Y′/R边际增长减少10%时的开发率; (3) E50: 资源量下降到原始水平50%时的开发率[18]。
本文采用Excel 2003, R软件3.0.1以及CorelD RAW 12进行数据录入、处理分析和绘图。
2 结果
2.1 年轮特征
拟尖头 鲌的鳞片(图 2a)为典型的圆鳞, 鳞片较大, 鳞焦位于鳞片中央。前区以及左右侧区相连, 从鳞片最边缘到鳞焦中心距离几乎等同; 环片在前区以及左右两个侧区排列致密且纤细, 后区则排列比较松散, 具放射沟及稀疏的粒状突起, 造成轮纹在后区不够完整或连续, 但在部分鳞片上也较为清晰和完整(图 2a); 前区及其两侧区的环片排列整齐,近似呈同心圆排列, 连接成较为明显的轮纹。
观察到的拟尖头鲌鳞片上的年轮特征主要为疏密和切割结构, 并伴随有稍微破碎现象, 其主要表现形式为: 环片在前区成疏密排布, 而在左右侧区形成切割(2a、2c: 箭头2、2d: 箭头4、5)。部分个体具有副轮, 表现为年轮上区域性的破碎或不完整(2c, 箭头 1)。部分鳞片特别是年龄较高的个体, 在靠近鳞焦的区域往往出现幼轮, 幼轮通常环片间距较大, 透光性较强(图2d: 箭头3)。
2.2 体长、体重和年龄分布
对1141 尾拟尖头 鲌样本进行体长、体重分析分析, 结果表明: 小江拟尖头 鲌的体长范围为67—450 mm, 平均体长为 165 mm, 优势体长范围为100—150 mm, 占总尾数的 53.12%; 体重范围为2.9—1067.4 g, 平均体重为92.2 g, 优势体重范围为0—100 g, 占总尾数的75.21% (图3)。选取425尾鳞片进行年龄分布分析, 结果表明: 2—3龄年龄组个体为优势年龄组, 共334尾, 占总抽样样本的78.59%, 1龄个体最少, 仅有5尾, 占1.18% (图3)。
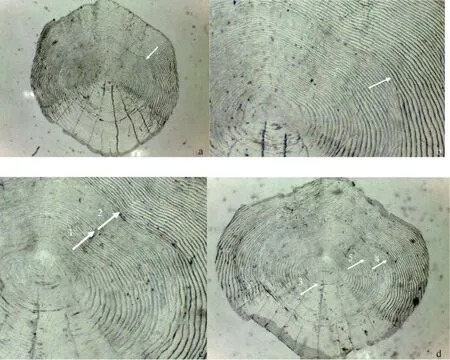
图2 拟尖头 鲌鳞片的形态与年轮特征Fig. 2 Morphology and annual ring characteristics of scale of Culter oxycephaloides
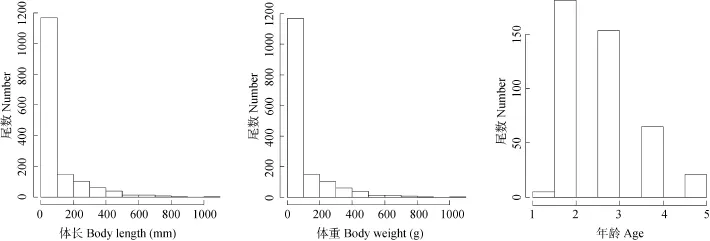
图3 拟尖头 鲌的体长、体重、年龄的尾数分布图Fig. 3 Number distribution of body length, weight and age of Culter oxycephaloides
2.3 体长和体重关系
根据测量的1741 尾拟尖头 鲌的体长和体重数据, 拟合出其体长-体重的回归关系, 两者呈幂函数关系: W = 0.000007L3.10, R2=0.99, N=789(图4)。经t检验, 参数 b=3.10与 3.0近似(P>0.05), 表明拟尖头 鲌属于匀速生长类型, 可以采用 Von Bertalanffy方程进行生长拟合。经残差平方和检验, 雌雄个体(雌鱼197尾; 雄鱼174尾)的体长-体重关系无明显差异(b=3.32和3.04, P>0.05)。
2.4 生长方程
根据最小二乘法计算得到生长方程的参数为L∞=519.63 mm、W∞=1806.08 g、k=0.22/a、, t0=–0.12 龄。据此得出的拟尖头 鲌体长和体重生长方程分别为(图5):

将上述方程分别求一阶和二阶微分, 得到体长和体重的生长速度方程和生长加速度方程分别为:
体长生长速度方程
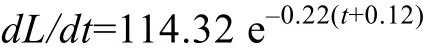
体长生长加速度方程
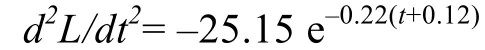
体重生长速度方程
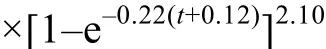
体 重 生 长 加 速 度 方 程
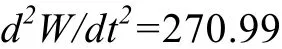
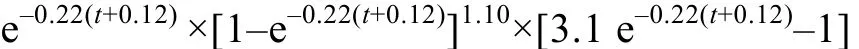
根据上述 4方程, 分别做出体长和体重生长速度、生长加速度曲线(图6)。
拟尖头 鲌的体长的绝对增长随年龄的增加而下降, 其生长速度先以较快的趋势下跌, 随后下跌的速度变缓, 而体长生长加速度一直增加, 最后趋于平缓; 体重的生长速度达到最大或体重加速度为 0时的年龄为5.02龄(体长348 mm、体重558.8 g), 此处即为拟尖头 鲌的生长拐点, 在此之前, 体重生长加速度值为正, 其后则为负值。同时体重生长加速度曲线表现为明显的“S”性状, 其中在 1.85龄以前呈明显的上升趋势, 随后逐渐减少, 直至到加速度为 0, 表明低龄幼鱼个体的生长速度明显高于较高年龄个体鱼类的生长速度(图6)。

图4 小江拟尖头 鲌的体长与体重的关系Fig. 4 The relationship between body length and weight of Culter oxycephaloides in Xiao River
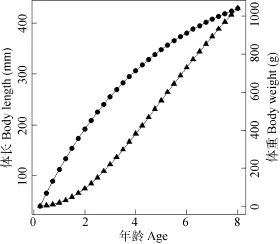
图5 小江拟尖头 鲌的体长(·)、体重(▲) 生长方程Fig. 5 Growth curves of body length (·) and body weight (▲) for Culter oxycephaloides in the Xiao River

图6 小江拟尖头 鲌的体长(·)、体重(▲) 生长速度和生长加速度方程Fig. 6 Growth rate (·) and growth acceleration rate (▲) of body length and body weight for Culter oxycephaloides in the Xiao River
2.5 死亡特征与开发率
总死亡系数的拟合曲线为 lnN/∆t=8.286–0.99t (R2=0.97), 故总死亡系数Z为0.99/a, 95%的置信区间为0.87—1.12/a (图 7)。经 Pauly经验公式, 求得自然死亡系数M=0.49/a。通过Z、M求得捕捞死亡系数F=0.5/a, 并进一步求得开发率E=0.51、总死亡率a=0.63、自然死亡率v=0.31和捕捞死亡率u=0.32。
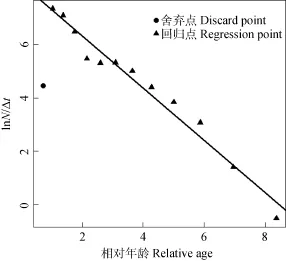
图7 根据体长变换渔获曲线估计小江拟尖头 鲌的总死亡系数Fig. 7 Estimate of total morality coefficient from length converted catch curve for Culter oxycephaloides in the Xiao River
2.6 繁殖特征
对渔获物中 124尾样本进行了抽样解剖, 并全部鉴定了年龄。在 124尾样本中, 卵巢Ⅳ和Ⅴ期样本共有46尾, 其中3龄个体22尾, 4龄个体19尾, 5龄个体5尾; 精巢Ⅳ和Ⅴ期样本共有20尾, 其中2龄个体1尾, 3龄个体10尾, 4龄个体6尾, 5龄个体3尾。因此, 可以认为拟尖头鲌 的最小性成熟年龄为2龄。
选择 31尾发育较好的Ⅳ期鱼卵进行绝对繁殖力和相对繁殖力估算, 结果表明: 拟尖头 鲌的绝对繁殖力变动范围为 20382—88430粒/尾, 平均值为49941 g/尾; 相对繁殖力变动范围为 54—161粒/g,平均值为 104粒/g。采用线性模型、对数模型等 7种模型对拟尖头 鲌的体重与绝对繁殖力的关系(N=31尾)进行曲线拟合, 所有回归模型的模拟相伴概率值均小于显著性水平0.05, 其中又以 2次方程曲线的判定系数(R2)最高, 因此选择2次方程曲线作为小江拟尖头 鲌体重与绝对繁殖力关系的拟合曲线,其关系式为:式中:为绝对繁殖力, W为体重。
选择3尾卵粒饱满的Ⅳ期卵巢, 对其测量卵径,得到卵径频率分布(图8)。从图8可知: 小江拟尖头鲌Ⅳ期卵巢卵径的平均值为 0.1060 cm, 最小值为0.0675 cm, 最大值为 0.1325 cm; 卵径分布存在两个明显的高峰, 表明卵巢中存在两种类群的卵子,小江拟尖头 鲌为分批产卵类型。

图8 小江拟尖头 鲌的卵径分布Fig. 8 Distribution of egg diameter of Culter oxycephaloides in the Xiao River
2.7 相对补充单位渔获量
经体长变换渔获量曲线分析, 当前小江拟尖头鲌的补充体长为88 mm, 但是由于目前小江区域内捕捞强度大, 许多网具捕捞大量幼鱼个体, 因此认为开捕体长等于渔获物中体长最小值, 即小江拟尖头 鲌当前开捕体长 Lc=67 mm。采用 Beverton-Holt动态综合模型分析, 由相对单位补充渔获量Y'/R与开发率 E关系作图估算出理论开发率 Emax=0.40/year, E10=0.32/year, E50=0.25/year(图9), 而当前开发率 Ecur=0.51/year, 高于理论最佳开发率, 小江拟尖头 鲌处于过度捕捞状态, 其种群将在未来有所减少。当Lc=67 mm, E>0.51/year时, Y'/R随开发率的增大呈减小趋势; 当Lc=67 mm, E<0.51/year时, Y'/R随E的增大呈现增加后, 当E值从当前的0. 51降至 Emax(0.40)时, 对应的 Y'/R 达到最大(Y'/R≈0.0147), 若再降至E50, Y' /R值(Y' /R≈0.0126)会有所下降.从生物量资源的维持角度来看, 当 E= 0.40/year时, 只有30% 的生物量资源得到保持, 而当E50= 0.25/year时, 约50%的生物量资源能得到保持(图 9)。在保持当前 M/K=2.23以及开发率

图9 Lc=67 mm时的Y′/R和B′/R二维分析Fig. 9 Two-dimensional analysis for Y′/R and B′/R when Lc=67 mm
(0.51/year)不变的前提下, 为获得更高的 Y′/R 以及
B′/R, 以保持种群不被过度捕捞, 其开捕体长应该控制在140 mm 以上。同时考虑到拟尖头 鲌2—3龄个体为繁殖高峰群体, 而 3龄个体的体长按生长方程反推为255 mm, 因此为保护小江的拟尖头 鲌资源, 其最小起捕体长至少为255 mm。
3 讨论
3.1 年龄鉴定材料的选择
鱼类年龄鉴定是研究生物学和生态学特性的基础, 也是分析和评价鱼类种群数量变动趋势的基本依据之一[11]。鉴定鱼类年龄的材料常见的有臀鳞、耳石、脊椎骨等。长期以来, 鳞片因取材方便, 后期处理容易, 对鱼体损害小等优点在鱼类年龄鉴定, 特别是在淡水鱼的年龄鉴定上被广泛使用[13—15, 19—21]。对于小江拟尖头 鲌而言, 其年龄结构较为简单, 最高年龄个体为5龄, 且生长速度较快(k=0.22/a), 国外研究认为采用鳞片对低龄个体鱼类进行年龄鉴定可以避免因高龄个体导致的年轮忽略或误认为副轮而造成年龄鉴定的误差[23], 而沈建忠等[24]对鲫的鳞片和耳石比较研究则表明: 用鳞片鉴定鲫年龄, 低龄(4龄以下)和生长较快个体精确度较高、与耳石年轮读数的吻合率也较高。基于以上研究结果, 并考虑到鳞片采样容易, 且后期处理耗时较少等因素, 我们采用鳞片作为 拟尖头 鲌的年龄鉴定材料,并采用 3个不同人员对同一鳞片年轮进行鉴定, 并最终经讨论确定该鳞片的最终年龄。尽管采用上式方法, 但随着年龄的增长, 鳞片会存在磨损现象,容易导致鳞片的误判, 需要在未来采用其他材料进行比对。
3.2 年龄结构与生长参数
小江拟尖头 鲌以 1—5龄个体组成, 其中 2—3龄为优势年龄组, 与三峡库区主要经济鱼类如鲢、铜鱼、鲤等年龄结构基本一致, 均以 4龄以下个体为捕捞群体的主要年龄构成[25]。在渔获物中, 1龄个体数量很少, 这很可能与 1龄个体个体规格很小,暂未完全进入渔业有关。
通过对生长参数进行比较, 发现小江拟尖头鲌的生长参数 k (0.22/year)的值比同属的分布在鲇鱼山水库的翘嘴Culter alburnus[26]的k值(0.7089/year)小很多, 但比分布在徐家河水库的翘嘴 鲌的 k值(0.0899/year)[27]大很多。同种鱼类的生长参数k在不同调查区域的差异在许多鱼类中均有出现[14, 28, 29],造成这种现象的原因除与不同区域的水域生态环境、水体饵料生物的丰歉以及种质资源差异有关[28]外, 还与采样方法有关[14]。为减少采样误差, 我们在小 江采用不同网目大小网具对拟尖头 鲌进行了采样, 而非采用单一网目大小网具进行采样, 同时为进一步减少采样误差, 我们将在不同江段的年龄数据合并成一个大的样本。
3.3 小江拟尖头 鲌的繁殖状况
小江拟尖头 鲌的最小性成熟年龄与丁瑞华(1994)的记载[5]一致, 均为2龄, 与同属的翘嘴 鲌也一致[27], 其和翘嘴 鲌一样也是分批产卵类型[27]。同时, 2013年4—7月调查期间, 小江拟尖头 鲌的绝对繁殖力变动范围为 20382—88430粒/尾, 平均值为49941 g/尾, 远小于 鲌亚科中相近种类— 翘嘴 鲌的平均绝对繁殖力[27]。一般而言, 在早期生活史阶段死亡率较高的种类, 其绝对繁殖力往往较高[27, 30],表明拟尖头 鲌的卵苗死亡率很可能相较翘嘴 鲌为低。同时, 小江拟尖头 鲌绝对繁殖力与体重之间的拟合关系以二次方程为最优, 表明小江拟尖头 鲌的绝对繁殖力与体重不成简单的线性关系, 绝对繁殖力的大小还与体重以外的因素相关, 这种情况在其他鱼类如北盘江云南光唇鱼 Schizothorax lissolabiatus等[31]也有出现。
拟尖头 鲌为产漂流性卵的鱼类种类, 其鱼卵的孵化需要在流水江段。根据2013年4—7月的实地调查, 小江分布有拟尖头 鲌的天然产卵场, 且其产卵场分布在养鹿以上的流水江段, 特别是位于渠口附近的砂砾底质江段, 但这些江段的水深较浅, 一般在1—5 m左右, 容易受到人类活动的影响。在实地调查中, 5月底到6月中旬, 渠口附近江段聚集了大量渔民, 有时候 3 km 左右江段分布有渔船10—20艘, 主要捕捞到该江段产卵的拟尖头 鲌、翘嘴 鲌等鱼类。
3.4 小江拟尖头鲌的资源保护及其利用
拟尖头鲌已逐渐成为三峡库区小江江段主要的经济鱼类之一, 包括拟尖头 鲌 在内 鲌类是在库区能够自然增殖的肉食性鱼类, 保持合理的 鲌属鱼类种群数量, 可以将水体中未被利用的小型鱼类转化成经济价值高的鱼产品, 这对于增加渔业经济效益,维持水体中种间平衡具有十分重要的意义[32, 33]。但近年来, 由于当地渔业习惯以及经济利益驱使, 小江拟尖头 鲌的资源状况实际上已开发过度, 主要体现在: (1)拟尖头鲌渔获物主要以 2—3龄个体为主,而拟尖头 鲌的最小成熟年龄为 2龄。此时, 刚进入生殖高峰阶段的个体被大量捕捞, 使得种群繁殖能力大大减弱; (2)渔获物中体长在100—150 mm的个体占总渔获量的比重最大, 表明大量低龄个体被捕捞; (3)小江拟尖头鲌当前开发率Ecur=0.51, 高于获得最大渔获量的开发率Emax=0.40。对此, 针对小江拟尖头鲌资源的现状, 建议采取以下保护措施: (1)控制捕捞是保护鱼类资源的重要方法[34]。由于小江渔获物中拟尖头 鲌的繁殖盛期年龄为2—3龄, 且拐点年龄为5.02龄, 3—5 龄为小江拟尖头 鲌体重增加迅速的年龄, 因此为保护小江的拟尖头 鲌资源, 规范其最小起捕年龄为5龄, 即体长至少为342 mm。同时, 渔政部门应严格限制网目大小来保护幼鱼,禁止库区非法的定置拦网、张网等有害渔具的使用。严厉打击电打、炸鱼、偷鱼等违法捕捞方式, 保证幼鱼和亲鱼的正常生长和繁殖。(2)拟尖头鲌在小江有产卵场, 应加强产卵场的生境保护, 在每年的5—7月杜绝在产卵场附近区域开展采砂和捕捞活动,使鱼类的正常产卵活动不受到严重干扰。最好在小江上游渠口附近设立鱼类资源保护区, 并进行有效的管理。
[1] Chen Y Y. Fauna Sinica, Osteichthyes, Cypriniformes [M]. ⅡBeijing: Science Press. 1998, 185—188 [陈宜瑜. 中国动物志, 硬骨鱼纲, 鲤形目(中卷). 北京: 科学出版社. 1998, 185—188]
[2] Lou Y L. Some clarifications on the Cultrinae fishes of China [J]. Acta Hydrobiologica Sinica, 1994, 18(1): 45—49[罗云林. 鲌 鲌属和红 属模式种的订正. 水生生物学报, 1994, 18(1): 45—49]
[3] Yue P Q, Lou Y L. Preliminary studies on phylogeny of subfamily Cultrinae (Cpriniformes and Cyprinidae) [J]. Acta Hydrobiologica Sinica, 1996, 20(2): 182—185 [乐佩琪, 罗云林. 鲌亚科鱼类系统发育初探. 水生生物学报, 1996, 20(2): 182—185]
[4] Yi B L, Zhu Z R. Studies on culter and culterichthys of China [J]. Acta Hydrobiologica Sinica, 1959, (2): 170—199[易伯鲁, 朱志荣. 鲌 鲌中国的 属和红 属鱼类的研究. 水生生物学集刊, 1959, (2): 170—199]
[5] Ding R H. The fishes of Sichuan, China [M]. Chengdu: Sichuan Publishing House of Sciences and Technology. 1994, 230—232 [丁瑞华. 四川鱼类志. 成都: 四川科学技术出版社. 1994, 230—232]
[6] Lin J, Lei H S, Yin X G, et al. A study on environmental comprehensive harressing of small reservoir basin-an analysis of environment of Xiaojian River valley in Kaixian [J]. Journal of Chongqing University (Social Sciences Edition), 2004, 10(3): 1—3 [林建, 雷亨顺, 尹希果, 等. 水库小流域环境综合治理研究—开县小江流域环境分析.重庆大学学报(社会科学版), 2004, 10(3): 1—3]
[7] Wang X Q, Guo J S. Change of water environmental capacity in the Xiaojiang River after impoundment of the Three Gorges Reservoir [J]. Research of Environmental Sciences, 2012, 25(1): 36—42 [王晓青, 郭劲松. 三峡水库蓄水后小江水环境容量的变化. 环境科学研究, 2012, 25(1): 36—42]
[8] Ran X B, Chen H T, Yao Q Z, et al. Characteristics of nutrients in the Xiangjiang Bay of the Three Gorges Reservoir [J]. Journal of Hydrobiology, 2009, 2(2): 21—27[冉祥滨, 陈洪涛, 姚庆祯, 等. 三峡水库小江库湾水体混合过程中营养盐的行为研究. 水生态学杂志, 2009, 2(2): 21—27]
[9] Wang X Q. Study on the Eutrophication and Models of Hydrodynamic and Water Quality in the Pengxi (Xiaojiang) River of Three Gorges Reservoir [D]. The paper of the Doctor’s Degree of Chongqing University, 2012 [王晓青.三峡库区澎溪河(小江)富营养化及水动力水质耦合模型研究. 重庆大学博士学位论文, 2012]
[10] Li B, Jiang X, Wang Z J, et al. The current situation of fishery resources in the Xiao River of the Three Gorge Reservoir [J]. Freshwater Fisheries, 2011, 41(6): 37—42 [李斌, 江星, 王志坚, 等. 三峡库区蓄水后小江鱼类资源现状. 淡水渔业, 2011, 41(6): 37—42]
[11] Yin M C. Fish Ecology [M]. Beijing: Chinese Agriculture Publishing House. 1995, 11—29 [殷名称. 鱼类生态学. 北京:中国农业出版社. 1995, 11—29]
[12] Sokal R R, Rohlf F J. Introduction to Biostatistics [M]. NewYork: Freeman. 1987, 225—227
[13] Deng Z L, Yu Z T, Xu Y X, et al. Age and growth of major commercial fishes of Han River [A]. In: Transactions of the Chinese Ichthyological Society (No.1) [C]. Beijing: Science Press. 1981, 97—111 [邓 中粦 , 余志堂, 许蕴玕 , 等. 汉江主要经济鱼类的年龄和生长. 鱼类学论文集(第一辑). 北京: 科学出版社. 1981, 97—111]
[14] Chen Y, Jackson D A, Harvey H H. A comparison of von bertalanffy and polinomial functions in modeling fish growth data [J]. Canadian Journal of Fisheries and Aquatic Sciences, 1992, 49(6): 1228—1235
[15] Wang M R, Yang S R, Liu F, et al. Age and growth of Rhinogobio cylindricus Günther in the upper reaches of the Yangtze River [J]. Acta Hydrobiologica Sinica, 2012, 36(2): 262—269 [王美荣, 杨少荣, 刘飞, 等.长江上游圆筒吻年龄与生长的研究, 水生生物学报, 2012, 36(2): 262—269]
[16] Pauly D. Length-converted catch curves and the seasonal growth of fishes [J]. Fishbyte, 1990, 8(3): 33—38
[17] Pauly D.On the interrelationships between natural mortality, growth parameters and mean environmental temperature in 175 fish stocks [J]. Journal du Conseil, 1980, 39(2): 175—192
[18] Pauly D. Fish population dynamics in tropical waters: a manual for use with programmable calculators [J]. Worldfish, 1984, (8): 325
[19] Gudrun G T, Larry D J. Fishery biology and biological reference points for management of ocean quahogs (Arctica islandica) off Iceland [J]. Fisheries Research, 2005, 75(1): 97—106
[20] Cao L W, Chen X L, Zhao J. Studies on age and growth of Megalobrama skolkovii in the Nanshui Reservoir [J]. Fisheries Science, 2001, 20(3): 11—14 [曹立文, 陈湘麟,赵俊. 南水水库鲂鱼的年龄与生长研究, 水产科学, 2001, 20(3): 11—14]
[21] Zhao H T, Wang J W, Hu F P, et al. Age and growth of Acrossocheilus yunanensis in the Upstream Chishui River [J]. Journal of Bijie University, 2009, 27(8): 77—82 [赵海涛,王剑伟, 胡付平, 等. 赤水河上游云南光唇鱼年龄与生长研究. 2009, 27(8): 77—82]
[22] Yang Z, Wan L, Tao J P, et al. Age and Growth of Coreius guichenoti in the Mainstream of the Yangtze River [J]. Journal of Hydroecology, 2011, 32(7): 46—52 [杨志, 万力,陶江平, 等. 长江干流圆口铜鱼的年龄与生长研究. 水生态学杂志, 2011, 32(7): 46—52]
[23] Niewinski B C, Ferreri C P. A comparison of three structures for estimating the age of yellow perch [J]. North American Journal of Fisheries Management, 1999, 19(3): 872—877
[24] Shen J Z, Cao W X, Cui Y B. Comparison of scale and otolith for estimating age of carassius auratusl [J]. Acta Hydrobiologica Sinica, 2001, 25(5): 461—465 [沈建忠, 曹文宣, 崔奕波. 用鳞片和耳石鉴定鲫年龄的比较研究. 水生生物学报, 2001, 25(5): 461—465]
[25] Wu Q, Duan X B, Xu S Y, et al. Studies on fishery resources in the Three Gorges Reservoir of the Yangtze River [J]. Freshwater fisheries, 2007, 37(2): 70—75 [吴强, 段辛斌,徐树英, 等. 长江三峡库区蓄水后鱼类资源现状. 淡水渔业, 2007, 37(2): 70—75]
[26] Gao Z P, Zhang J B, Hu Z Y. Studies on the population control and growth characteristics of Culter alburnus in the Nianyushan Reservoir [J]. Reservoir Fisheries, 2008, 28(3): 39—48 [高志鹏, 张家波, 胡正友. 鲇鱼山水库翘嘴 鲌生长特性与种群控制研究. 水利渔业, 2008, 28(3): 39—48
[27] Qin L. The study on the age, growth and reproduction biology of Culter alburnus in the Xujiahe Reservoir [D]. Huazhong Agricultural University, 2009 [覃亮. 徐家河水库翘嘴 鲌年龄与生长和繁殖生物学研究. 华中农业大学, 2009]
[28] Xin J F, Yang Y F, Liu H Z. Study on the age and growth of Rhinogobio ventralis in the Upper Reaches of the Yangtze River [J]. Sichuan Journal of Zoology, 2010, 29(3):
352—356 [辛建峰, 杨宇峰, 刘焕章. 长江上游长鳍吻年龄与生长的研究. 四川动物, 2010, 29(3): 352—356 ]
[29] David Wells R J, Kohin S, Teo S L H, et al. Age and growth of North Pacific albacore (Thunnus alalunga): Implications for stock assessment [J]. Fisheries Research, 2013, 147: 55—62
[30] Ye F L, Chen G. Study on life-history pattern of nineteen freshwater fishes [J]. Journal of Zhanjiang Ocean University, 1998, 18(3): 11—17 [叶富良, 陈刚. 19种淡水鱼类的生活史类型研究. 湛江海洋大学学报, 1998, 18(3): 11—17]
[31] Xiao H, Dai Y G. Study on individual fecundity of Schizothorax lissolabiatus in Beipan River [J]. Journal of Hydroecology, 2010, 3(3): 64—70 [肖海, 代应贵. 北盘江光唇裂腹鱼个体繁殖力的研究. 水生态学杂志, 2010, 3(3): 64—70]
[32] Qin L, Xiong B X, Wang J S, et al. Ecological function of culter in natural water and countermeasures of resources proliferation [J]. Hubei Agricultural Sciences, 2009, 48(1): 233—236 [覃亮, 熊邦喜, 王基松, 等. 鲌属鱼类在天然水域中的生态功能及资源增殖对策. 湖北农业科学, 2009, 48(1): 233—236]
[33] Hu H Y, Di Y, Zhao Y F, et al. Comparative studies on the morphological characteristics of four species of Culter and Culterichthys in Lihu Lake [J]. Journal of Yunnan Agricultural University, 2011, 26(4): 488—494 [胡海彦, 狄瑜, 赵永锋, 等. 蠡湖 4 种鲌 鱼形态特征的比较研究. 云南农业大学学报(自然科学版), 2011, 26(4): 488—494]
[34] Cao W X. Several problems about fish resources protection in the Yangtze River Basin [J]. Resources and Environment in the Yangtze Basin, 2008, 17(2): 163—164[曹文宣. 有关长江流域鱼类资源保护的几个问题. 长江流域资源与环境, 2008, 17(2): 163—164]
STUDY ON AGE, GROWTH, REPRODUCTION AND THE STATUS OF RESOURCE DEVELOPMENT OF CULTER OXYCEPHALOIDES
(KREYENBERG ET PAPPENHEIM) IN THE XIAO RIVER
DUAN Peng-Xiang1, YANG Zhi2, TANG Hui-Yuan2, XIAO Qiong1, GONG Yun2, XIAO Tiao-Yi1and LIU
Xiao-Yan1
(1. College of Animal Science and Technology,Hunan Agricultural University, Changsha 410000, China; 2. Key Laboratory of Ecological Impacts of Hydraulic-projects and Restoration of Aquatic Ecosystem of Ministry of Water Resources, Institute of Hydroecology, Ministry of Water Resources and Chinese Academy of Sciences, Wuhan 430079, China)
Culter oxycephaloides (Kreyenberg et Pappenheim), which belongs to genus Culter, subfamily Cultrinae, family Cyprinidae, order Cypriniformes. They are a unique species in China and mainly live in the rivers and lakes of the Yangtze River Basin. Due to the impoundment of the Three Gorges Reservoir, the ecosystem of the upper Yangtze River basin especially the tributaries of the Three Gorges Reservoir has been profoundly changed. The Xiao River is an important tributary of the Three Gorges Reservoir, and the habitat of a variety of organisms especially fishes. According to the data collected in 2013, some sections of the Xiao River have become major spawning grounds of Culter oxycephaloides. Hence it is critical to investigate the age structure and the growth characteristics of Culter oxycephaloides, in order to better manage and protect this fish resource. In April-July and October-December of 2013, we collected 1741 individuals from the Xiao River of the Three Gorges Reservoir and conducted the fish biology and ecology study. Our 245 samples could be divided into 5 age groups, and 2—3 year-old individuals accounted for 78.75% of the total samples. The relationship between the body length (L) and the body weight (W) could be fitted as W = 0.000007L3.10(R2=0.9, N=789). The Von Bertalanffy growth function was Lt=519.63[1–e–0.22(t+0.12)],Wt=1806.08 [1–e–0.22(t+0.12)]3.10. The growth inflexion point of the body weight was 5.02 years, and the corresponding body length was 348 mm and the body weight was 558.8 g. The absolute fecundities ranged between 20382 and 88430 eggs per fish, and the relative fecundities were between 54 and 161 eggs/g. The eggs of mature females were 0.03—0.14 cm in diameter, with an average of 0.11 cm. Based on the frequency distribution plot of egg diameters, we deduced that the development of oocytes in the ovaries of Culter oxycephaloides was in a successive process and the female could spawn twice within a reproductive period. We compared the coefficients of the linear equation, the logarithmic equation, the quadratic equation, the Cubic equation, the compound equation, the power equation and the exponential equation, and found that quadratic equation best described the relationship between the body weight and the absolute fecundity (R2=0.49, sig.= 0.02<0.05, N=31). The exploitation rate of Culter oxycephaloides in the Xiao River was 0.51/year in 2013. The Beverton-Holt dynamic model indicated that Emax(the exploitation rate for the maximum yield), E10(the exploitation rate at which the marginal increase of relative yield-per-recruit is 1/10th of the value at E=0) and E50(the exploitation rate at which the stock is reduced to 50% of the original biomass) were 0.40/year, 0.32/year, and 0.25/year respectively. The comparison between Emax and the exploitation rate in 2013 demonstrated that Culter oxycephaloides in the Xiao River were excessively exploited in 2013. Therefore, the future fishing activity in the Xiao River should be strictly supervised in order to maintain and enlarge the population of Culter oxycephaloides.
Xiao River; Culter oxycephaloides; Age and growth; Reproduction; Resource development
S932.4
A
1000-3207(2015)04-0695-10
10.7541/2015.92
2015-01-12;
2015-05-04
国家自然科学基金(51209151,51379134); “十二五”国家科技支撑计划课题(2012BAC06B01); 三峡后续工作科研项目(2013HXKY2-3)资助
段鹏翔(1988—), 男, 湖南娄底人; 硕士研究生; 研究方向为鱼类生态学与鱼类病害学。E-mail: 18066966@qq.com
刘小燕(1965—), 女, 湖南长沙人; 博士, 教授; 主要研究方向为鱼类病害学。E-mail: liuxy186@163.com
猜你喜欢
天天爱科学(2022年9期)2022-09-15
昆明医科大学学报(2022年1期)2022-02-28
快乐语文(2020年32期)2021-01-15
猪业科学(2016年1期)2016-03-30
中国海洋大学学报(自然科学版)(2014年9期)2014-02-28
优雅(2014年2期)2014-02-24
阅读与鉴赏·学术版上旬刊(2010年11期)2010-01-19
民间文学(2009年6期)2009-07-07
新作文·小学低年级版(2009年9期)2009-02-09
故事会(2008年16期)2008-05-14
