
中街山列岛海洋保护区棘头梅童鱼生物学特征与资源密度的季节变化
2015-02-22 01:20张洪亮贺舟挺周永东
浙江海洋大学学报(自然科学版) 2015年5期
张洪亮,王 洋,梁 君,贺舟挺,周永东
(浙江省海洋水产研究所,农业部重点渔场渔业资源科学观测实验站,浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江舟山 316021)
中街山列岛海洋保护区棘头梅童鱼生物学特征与资源密度的季节变化
张洪亮,王 洋,梁 君,贺舟挺,周永东
(浙江省海洋水产研究所,农业部重点渔场渔业资源科学观测实验站,浙江省海洋渔业资源可持续利用技术研究重点实验室,浙江舟山 316021)
根据2012年冬季(2月)、春季(5月)、夏季(8月)与秋季(11月)进行的共4个航次的底拖网渔业资源调查资料,对中街山列岛海洋保护区棘头梅童鱼的生物学特征与资源密度进行了分析。结果表明:四季中,夏季存在明显的幼鱼体长组与体重组;摄食强度以冬季为最低;性腺成熟度春季达最高;重量密度与尾数密度以夏季为最高,春季最低,且夏季与春季间均存在显著性差异(P<0.05),这应与春季卵孵化后的幼体在夏季已能被底拖网所捕获有关;重量密度与平均底盐呈二次曲线相关(P<0.05)。
中街山列岛;棘头梅童鱼;摄食强度;性腺成熟度;资源密度
中街山列岛海洋保护区位于舟山群岛新区东部,优越的地理位置与海洋环境条件使之成为众多海洋生物繁殖、摄食和生长的重要栖息地[1]。有关该保护区内栖息的海洋生物研究主要集中在鱼类、甲壳类、软体动物与底栖生物的资源状况、群落结构与多样性等方面[1-10],但涉及某一具体物种的研究很少,仅曼氏无针乌贼Sepiella maindroni[11-12]与厚壳贻贝Mytilus coruscus[13]。棘头梅童鱼Collichthys lucidus广泛分布于我国沿海[14],并为人民所喜食,它是浙江近海群众渔业的主要捕捞对象之一[15],也是中街山列岛海洋保护区重要的鱼类优势种之一[1]。本文拟根据2012年周年四季的底拖网渔业资源调查资料,对中街山列岛海洋保护区内棘头梅童鱼的体长、体重、摄食强度、尾数密度与重量密度的季节变化进行研究,以丰富该保护区内具体物种的研究数量,同时为该鱼种的管理与可持续利用提供科学依据。
1 材料与方法
1.1 数据来源
数据来源于2012年冬季(2月)、春季(5月)、夏季(8月)与秋季(11月)对中街山列岛海洋保护区进行的共4个航次的渔业资源与温度、盐度调查资料,调查时间为每航次3 d(小潮汛期间),站位设置如图1所示。渔业资源调查采用底拖网,各站位分别调查1次,平均拖速5.19 km/h,每次拖曳1 h。底拖网网口拉紧周长25 m,囊网网目尺寸3.0 cm。温度与盐度数据用温盐深测定仪(CTD)采集,每航次每站位各1次。每航次底拖网样品冰鲜保存后带回实验室进行种类鉴定与体重、体长、性别、摄食强度等常规生物学测定。摄食强度与性腺成熟度按《海洋调查规范》[16]的规定进行鉴别;体重与体长精度分别为0.1 g和1 mm。调查船为普渔4126号,该船长19 m、宽3.3 m,主机功率58.8 kW。
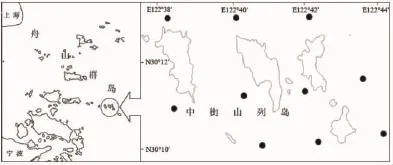
图1 采样区域和站位设置Fig.1 Location of the study area and distribution of the survey positions
1.2 数据处理方法
棘头梅童鱼的资源密度用扫海面积法[17]进行估算,其计算式为:
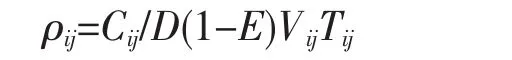
式中:ρij为i月j站位的资源密度(kg/km2或103ind/km2),Cij为i月j站位的网次渔获量(kg)或尾数(ind.),D为网口水平扩张宽度(km)(本网具为5.20×10-3km),Vij为i月j站位的平均拖速(km/h),Tij为i月j站位的作业时间(h),E为逃逸率(取0.5[18])。
显著性检验与相关性分析分别采用SPSS16.0的方差分析(ANOVA)与曲线回归分析(CURVE ESTIMATION)。其中,用于显著性检验的所有数据经过log(x+1)预处理。
2 结果
2.1 体长组成的季节变化
冬季,棘头梅童鱼的体长变化范围为45~136 mm,平均值为91.7 mm,优势组为80~110 mm(图2);春季,体长变化范围为87~144 mm,平均值为110.0 mm,优势组为90~130 mm;夏季,体长变化范围为25~129 mm,平均值为66.8 mm,优势组分2组,分别为30~50 mm与70~90 mm;秋季,体长变化范围为77~143 mm,平均值为98.9 mm,优势组为80~110 mm。

图2 棘头梅童鱼体长分布的季节变化Fig.2 Seasonal variations of body length distribution of C.lucidus
2.2 体重组成的季节变化
冬季,棘头梅童鱼的体重变化范围为1.5~52.1 g,平均值为16.4 g,优势组为9~24 g(图3);春季,体重变化范围为12.3~54.6 g,平均值为26.3 g,优势组为15~30 g;夏季,体重变化范围为0.1~39.0 g,平均值为9.0 g,优势组分2组,分别为0~3 g与6~15 g;秋季,体重变化范围为8.4~55.4 g,平均值为20.0 g,优势组为9~21 g。
2.3 摄食强度的季节变化
从图4可以看出,冬季,棘头梅童鱼的摄食强度从高到低依次为1级、2级、0级、3级与4级,其中1级占绝对优势,为位列第二高的2级的3.70倍;春季,摄食强度从高到低依次为1级、2级、3级、4级与0级,其中1级与2级之和占总数的85.71%;夏季,摄食强度从高到低依次为3级、2级、1级、4级与0级,其中0级仅为位列第二低的4级的0.15倍,显著低于其它各级;秋季,摄食强度从高到低依次为2级、1级、4级、3级与0级,其中0级为位列第二低的4级的0.20倍,明显低于其它各级。另外,除冬季外,春季、夏季与秋季0级的比例均很低,最高不超过3.02%。

图4 棘头梅童鱼摄食强度的季节变化Fig.4 Seasonal variations of feeding level of C.lucidus

图4 棘头梅童鱼摄食强度的季节变化Fig.4 Seasonal variations of feeding level of C.lucidus
2.4 性腺成熟度的季节变化
在可分辨性别(成熟期≥Ⅱ期)的样品中,棘头梅童鱼冬季、春季、夏季与秋季的雌雄性比相近,大致都为6:4。在所有可判别性别的雌性个体中,其四季性腺成熟情况如图5所示。由图5可知,性成熟个体(Ⅳ期与Ⅴ期)比例从高到低依次为春季、夏季、冬季与秋季,最高值为最低值的3.00倍;已产卵个体(Ⅵ)出现在春、夏两季,但春季的比例显著高于秋季。
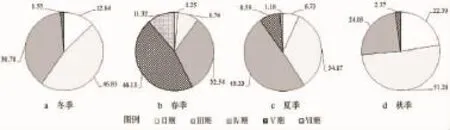
图5 棘头梅童鱼雌性个体性腺成熟度(%)的季节变化Fig.5 Seasonal variations of female gonadal maturity of C.lucidus
2.5 资源密度的季节变化
图6是棘头梅童鱼平均资源密度的四季分布图。由图可看出,棘头梅童鱼的重量密度随夏季、秋季、冬季与春季的顺序呈逐季降低的趋势,其最大值为最小值的6.22倍;尾数密度则随夏季、冬季、秋季与春季的顺序呈逐季降低的趋势,其最大值为最小值的20.72倍。显著性检验结果表明,对于重量密度,春季与夏季间存在显著性差异(P=0.046<0.05);对于尾数密度,春季与冬季、夏季间均差异显著(分别为P=0.036<0.05与P=0.019<0.05)。
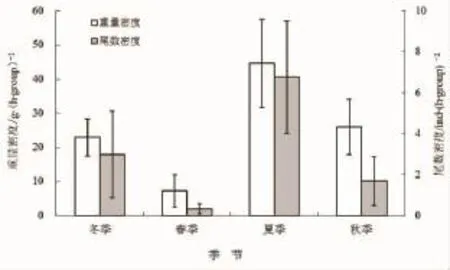
图6 棘头梅童鱼资源密度的季节变化Fig.6 Seasonal variations of abundance density of C.lucidus
3 讨论
3.1 资源密度与底温、底盐之间的关系
调查期间,中街山列岛海洋保护区冬季、春季、夏季、秋季的平均底温分别为11.27℃、16.07℃、20.64℃与21.09℃,底盐分别为29.70、31.13、34.17与29.41。回归分析结果表明,在棘头梅童鱼的重量密度、尾数密度与底盐、底温的关系中,仅重量密度(B)与底盐(S)之间呈二次曲线相关(P=0.019<0.05)。
3.2 体长、体重组成及其与性腺成熟度的关系
在棘头梅童鱼的冬、春、夏、秋四季的体长组成中,仅夏季出现两个优势体长组,这与体重优势组的出现情况一致,两者可相互印证。另外,无论是性成熟个体还是已产卵个体的比例春季均明显高于其它季节,这与浙江近海棘头梅童鱼的产卵盛期为5月下旬至6月中旬的研究结果相符[19]。由于底拖网对渔获物的体长与体重具有一定的选择性,即刚孵化不久的幼体很难被底拖网所捕获,因此,夏季30~50 mm优势体长组(或0~3 g优势体重组)的个体应为春季卵所孵化后长成。
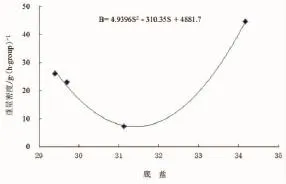
图7 棘头梅童鱼重量密度与底盐的关系Fig.7 Relationship between the weight density of C.lucidus and sea bottom water salinity
3.3 摄食强度的季节变化分析
冬季,棘头梅童鱼的摄食强度为0级(即空胃)的比例为13.91%,显著高于其它各季(春季、夏季与秋季分别为2.38%、1.89%与3.02%),同时1级(少量食物)的比例也为四季中最高(冬季、春季、夏季与秋季分别为56.29%、53.17%、23.58%与23.58%),因此,该季棘头梅童鱼的摄食强度为一年四季中的最低,这与严利平等[20]对黄海南部、东海北部小黄鱼摄食强度变化研究的结论相一致,即越冬期间的食欲较弱,为弱摄食期。这一点可从鱼类行为学方面进行解释,即冬季较低的水温导致鱼类的活动减少,从而摄食活动也相应地降低。
3.4 资源密度的季节变化分析
根据显著性检验结果,无论是棘头梅童鱼的重量密度还是尾数密度,夏季均显著高于春季,这应与春季产卵孵化后的幼体在夏季已能被底拖网所捕获有关。
[1]梁 君,徐汉祥,王伟定.中街山列岛海洋保护区鱼类物种多样性[J].生态学报,2013,33(18):5 905-5 916.
[2]梁 君,王伟定,虞宝存,等.中街山列岛海洋保护区岛礁生境鱼类资源及群落多样性季节变化[J].海洋与湖沼,2014,45 (5):979-989.
[3]张 龙,徐开达,张洪亮,等.中街山海域渔业动物群落结构的季节变化[J].浙江海洋学院学报:自然科学版,2012,31(4): 290-294.
[4]徐开达,卢占晖,薛利建,等.中街山列岛保护区蟹类群落结构特征的研究[J].上海海洋大学学报,2012,21(6):1 032-1 037.
[5]徐开达,宋海棠,张 龙,等.中街山列岛保护区虾类资源状况及其与环境因子的关系[J].浙江海洋学院学报:自然科学版,2012,31(5):420-425.
[6]刘小雄,王伟定,梁 君,等.中街山列岛岛礁生境游泳动物群落多样性及稳定性分析[J].浙江海洋学院学报:自然科学版,2014,33(1):14-18.
[7]徐开达,张洪亮,谢汉阳,等.中街山列岛水域甲壳类资源及其群落多样性[J].海洋渔业,2012,34(3):308-315.
[8]姚海峰,薛 巍,邹广明,等.中街山列岛岩礁潮间带春季大型底栖动物的群落结构[J].安徽农业科学,2012,40(17):9 282-9 284.
[9]吴常文,王志铮,吕永林.中街山列岛软体动物种类组成及资源开发利用建议[J].浙江水产学院学报,1997,16(2):85-95.
[10]谢汉阳,朱文斌,徐开达,等.中街山水域虾类组成及其群落多样性[J].广东海洋大学学报,2012,32(4):1-7.
[11]吴常文,董智勇,迟长凤,等.曼氏无针乌贼(Sepiella maindroni)繁殖习性及其产卵场修复的研究[J].海洋与湖沼,2010,41 (1):39-46.
[12]李继姬,郭宝英,吴常文.浙江海域曼氏无针乌贼资源演变及修复路径探讨[J].浙江海洋学院学报:自然科学版,2011,30 (5):381-385.
[13]梁 君,虞宝存,毕远新,等.中街山列岛海域厚壳贻贝生物学特征及生长规律[J].生态学杂志,2015,34(2):471-476.
[14]贺舟挺,薛利建,金海卫.东海北部近海棘头梅童鱼食性及营养级的探讨[J].海洋渔业,2011,33(3):265-273.
[15]吴常文,王伟洪.棘头梅童鱼Collichthys lucidus个体生殖力的研究[J].浙江水产学院学报,1996,15(3):174-178.
[16]国家质量监督检验检疫总局.海洋调查规范第六部分:海洋生物调查[M].北京:海洋出版社,2015.
[17]张洪亮,梁 君,贺舟挺,等.岱衢洋渔业动物多样性分析[J].水生生物学报,2012,36(5):922-931.
[18]张洪亮,宋之琦,潘国良,等.浙江南部近海春季鱼类多样性分析[J].海洋与湖沼,2013,44(1):126-134.
[19]吴常文,王伟宏.浙江近海棘头梅童鱼的分布生物学与资源变动[J].海洋渔业,1991,13(1):5-10.
[20]严利平,李建生,沈德刚,等.黄海南部、东海北部小黄鱼饵料组成和摄食强度的变化[J].海洋渔业,2006,28(2):117-123.
Seasonal Variations of the Biological Characteristics and Abundance Density of Collichthys lucidus in Zhongjieshan Islands Marine Protected Area
ZHANG Hong-liang,WANG Yang,LIANG Jun,et al
(Marine Fisheries Research Institute of Zhejiang Province,Key Research Station for Fisheries Resources of Main Fishing Ground,Ministry of Agriculture,Key Research Station of Sustainable Utilization for Marine Fisheries Resources,Zhejiang Province,Zhoushan 316021,China)
The biological characteristics and abundance density of Collichthys lucidus in Zhongjieshan Islands Marine Protected Area were analyzed based on the data from the investigations in winter(February), spring(May),summer(August)and autumn(November)in 2012 by bottom trawls.The results showed that in all the four seasons,there were obvious body length group and weight group of younger fishes.The feeding level in winter was the lowest.The gonadal maturity in spring was the highest.Both the weight density and individual density were highest in summer and lowest in spring,due to the juvenile fishes bone in spring fed up and could be caught by bottom trawls in summer.The weight density in summer was specifically different withthat in spring(P<0.05),and similar to the individual density(P<0.05).The relationship between the weight density and sea bottom water salinity can be denoted by binary curve(P<0.05).
Zhongjieshan islands;Collichthys lucidus;feeding level;gonadal maturity;abundance density
S932.4
A
1008-830X(2015)05-0407-04
2015-04-28
浙江省海域海岛管理利用项目(浙海渔计[2013]149号);浙江省渔业资源增殖放流项目(浙财农[2014]277号);中央财政渔业资源保护项目(浙财农[2015]31号);东海区渔业资源和生态调查项目(东渔政[2013]144号与[2014]75号)
张洪亮(1968-),男,浙江嵊州人,高级工程师,研究方向:渔业资源与生物多样性.E-mail:hidalgo310@163.com
周永东.E-mail:zyd511@sina.com
猜你喜欢
品牌研究(2021年7期)2021-11-28
当代水产(2021年1期)2021-03-19
四川蚕业(2021年1期)2021-02-12
当代水产(2019年9期)2019-10-08
江西农业学报(2019年8期)2019-09-04
建材发展导向(2019年11期)2019-08-24
现代园艺(2018年2期)2018-03-15
中学生天地(A版)(2015年7期)2015-08-06
读者(乡土人文版)(2013年10期)2013-04-12
