
湖南白背飞虱前期迁入种群中小尺度虫源地及降落机制
2015-02-05 06:03吴秋琳陆明红朱秀秀张仲刚翟保平
生态学报 2015年22期
吴秋琳,胡 高,陆明红,王 标,朱秀秀,粟 芳,张仲刚,翟保平,*
1 南京农业大学昆虫学系,农业部&教育部病虫害监测与治理重点开放实验室, 南京 210095 2 全国农业技术推广服务中心, 北京 100125 3 湖南省植保站, 长沙 410005 4 洪江市植保站, 洪江 418200 5 芷江侗族自治县植保站, 芷江 419100
湖南白背飞虱前期迁入种群中小尺度虫源地及降落机制
吴秋琳1,胡 高1,陆明红2,王 标3,朱秀秀3,粟 芳4,张仲刚5,翟保平1,*
1 南京农业大学昆虫学系,农业部&教育部病虫害监测与治理重点开放实验室, 南京 210095 2 全国农业技术推广服务中心, 北京 100125 3 湖南省植保站, 长沙 410005 4 洪江市植保站, 洪江 418200 5 芷江侗族自治县植保站, 芷江 419100
湖南是白背飞虱(Sogatellafurcifera(Horváth))南北往返的必经之路和主害区。明确白背飞虱前期(至5月20日)迁入种群的中小尺度虫源地和降落机制,对实现精细化异地预测至关重要。择出2012年湖南稻区白背飞虱前期迁入种群的上灯峰日,用新一代中尺度数值预报模式WRF(Weather Rsearch and Forecast)做三维轨迹分析,明确虫源地分布,并对其迁飞过程进行多因素的时空动态分析。结果表明:(1)2012年湘西白背飞虱迁入虫源主要来自越南北部、广西西南部,少量来自广西东南部与广东南部。湘东南、湘中迁入虫源主要来自广东南部、广西东南部,少部分来自海南。此外,每年4月下旬与5月上旬我国南部盛行西南气流,湘东南位置偏东,加之南岭山脉的阻挡,该区迁入峰次与迁入量较湘西较少。(2)低空急流为白背飞虱的北迁提供了运载气流,锋面气旋与切变线等天气尺度系统以及强烈上升气流带来的降水、低温屏障与下沉气流是湖南稻区白背飞虱集中迫降的3个直接因素;(3)白背飞虱前期迁入种群在湖南的主要落点基本沿湘西武陵山区至湘南雪峰山区一带以及“湘桂走廊”东端分布,地形是导致湘西降虫发生频率高、迁入量大的主要因素。由于山体阻塞与地面摩擦形成的小型垂直环流、夜间山风的作用及山谷间的峡谷风可导致白背飞虱的集中降落,较大的灯诱峰次主要位于谷地或者是山体的迎风坡。
白背飞虱;中小尺度虫源地;降落机制;WRF模式;轨迹分析
湖南地处亚热带季风湿润气候区,是我国粮食主产区之一,其耕地面积占全国耕地面积的1/32,但生产的粮食占全国的1/19[1],其中水稻种植以双季稻为主、单双季稻混合,水稻播种面积和总产长期稳居全国第一,具有明显的水稻生产区位优势[2]。单双季稻共存的种植制度和高强度的水肥投入及滥用农药引发的抗药性和致害力不断加强,导致湖南白背飞虱(sogatellafurcifera(Horváth))的危害程度愈演愈烈[3]。2000—2012年,湖南白背飞虱年平均发生面积分别为250.618万hm2,发生程度为4级[2],直接威胁我国的粮食安全。因此,明确近年来白背飞虱暴发成灾的内外因素,提出有效准确的预警方法,是保障我国粮食安全和可持续发展的重中之重。
白背飞虱是亚洲稻区一种远距离迁飞性害虫,每年3—6月自中南半岛乘西南气流陆续迁入我国南部稻区[4]。从20世纪90年代以来,其发生面积日益扩大,为害程度加剧,暴发频率显著增加。2012年更出现了历年来规模最大的白背飞虱迁入峰,不仅迁入早、峰次多,而且迁入量特大。到4月底,广西灯下始现单灯单晚万头虫峰,5月4日桂北和黔东南始现单灯单晚十万头量级虫峰;5月1—17日连续出现超大规模主迁峰,同期突增区纵横千余公里(21°N—30°N、99°E—118°E),覆盖云贵川渝两湖两广闽赣10省区。其中广西融安5月2、4、5日分别出现67320头、192200头和556700头的上灯峰,5月6—10日5天累计诱虫808548头。到5月6日湖南已出现3次大的迁入峰,全省22个监测点灯下累计虫量3.9万头,是特大发生年2007年同期的2.4倍。湘西洪江5月4—6日单灯虫量分别为3596、3625和5935头,8日3万头;湘西怀化9日上灯3万多头。湘南蓝山县5月3日单灯诱虫过8070头,湘中邵阳5月6日上灯过万头,邵东7日2496头,均超历史灯诱记录。这些迁入种群源自何处?迄今尚无系统研究。
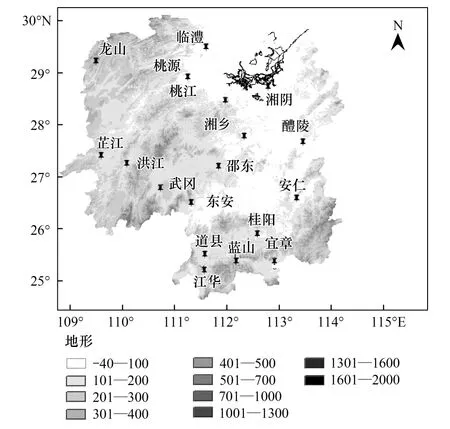
图1 湖南地形及各站点位置 Fig.1 Geographic and geomorphic environment and location of trap site of Hunan
湖南地处南岭山脉北麓(图1),东、南、西3面环山,全境有15座主峰在1400m以上的大山体;中部为丘陵盆地,北部为洞庭湖堆积平原,形似马蹄形盆地。湘东南东邻罗霄山脉,南倚南岭山脉。湘西武陵山、雪峰山和越城岭所夹山间谷地与位于越城岭与海洋山之间的“湘桂走廊”相接,构成夏季风北上和冬季风南下的重要通道,有利于从西南或东南方向北迁来的稻飞虱虫源进入该区而常年大发生[5]。地形导致迁飞性昆虫的降落主要有两方面因素:(1)气流遇山脉被迫爬升,随着山体地势的升高而温度降低,迫使稻飞虱由于生理适应而降低迁飞高度进而被迫降落[6]。同时,气流爬升引起的成云降雨会迫降空中虫群在迎风坡形成集中降落。(2)地形的阻碍作用使低层大气进入两山之间的谷地时产生“狭管效应”从而形成很强的峡谷风和辐合上升气流,导致地形雨[7],空中虫群遇雨而集中降落。究竟哪种特定的条件(包括天气系统、天气过程、地理条件等)是导致湖南稻区稻飞虱的集中降落的主要因素?影响湖南稻飞虱年度间数量变动的主要因子是什么?目前尚无深度研究。
为此,我们在UBUNTU12.04平台下运用Fortran语言编写的三维轨迹分析程序,模拟分析2012年湖南白背飞虱前期迁入种群可能的虫源地;通过分析WRF模式输出的高分辨率大气动力背景下的大气环流形势和大气流场时空变化过程,结合湖南地形地貌格局,确定湖南白背飞虱迁入种群的降落机制,为湖南白背飞虱的精细化异地预测提供科学依据。
1 材料与方法
1.1 灯诱数据与田间调查资料
全国农业技术推广服务中心病虫测报处提供的2012年湖南省24个植保站的白背飞虱灯下诱捕资料和田间系统调查资料。永州市植保站提供的永州市各县区灯下虫量数据。
1.2 地形数据
包括MODIS和GWD数据,覆盖180°W—180°E,90°S—90°N的所有区域,来自WRF(Weather Research and Forecast)官方网[8- 10],分辨率为30″、2′、5′和10′,本文模拟的Domain1与Domain2地形数据分辨率分别为10′与2′。
1.3 地图资料
采用中国省级行政区图(1∶4000000),资料来于国家基础地理信息中心[11]。
1.4 气象资料
包括美国国家环境预报中心(NCEP(National Centers for Environmental Prediction))/美国国家大气研究中心(NCAR(National Center for Atmospheric Research))的每6小时1次的1°×1°的全球分析资料[12];降水资料采用国家气象信息中心提供的小时融合降水产品(实时数据)[13]。
1.5 WRF模式
WRF模式系统是由NCAR、NCEP和美国预测系统实验室(FSL(Forecast Systems Laboratory))/美国国家海洋和大气管理局(NOAA, National Oceanic and Atmospheric Administration)等多家研究部门和大学联合开发的新一代中尺度数值模式。模式采用全可压、非静力学方程, 欧拉高度坐标及欧拉质量坐标两种坐标系,水平格点采用Arakawa C类格点。
1.6 WRF参数化方案(表1)
采用的WRF模式网格设计为二重嵌套,使用FNL全球分析资料(Final Operational Global Analysis Data)作为模式初始场和边界条件,二重嵌套网格的控制试验中边界层、辐射过程和微物理处理方案相同。资料输入WRF模式进行数值模拟后输出每小时1次的20Km×20Km格距的气象要素场,并经ARWpost转换为GrADS(Grid Analysis and Display System)格点数据。
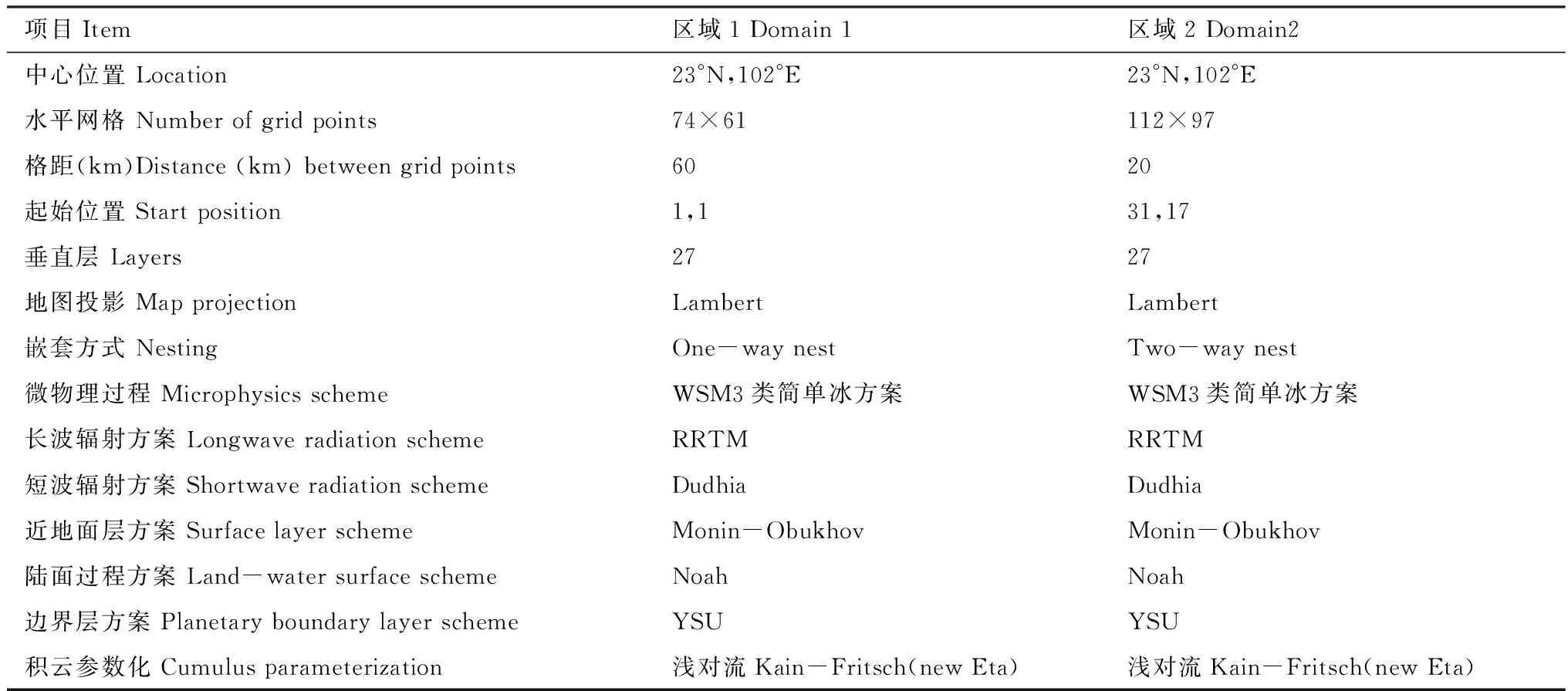
表1 WRF模式方案与参数Table 1 Selection of scheme and parameters of WRF
1.7 轨迹分析
应用WRF模式输出的高分辨率风温场以及高度场为基础,参照朱艳秋和廖洞贤的三维空气质点分析方法[14], 用Fortran语言编程计算稻飞虱的三维轨迹。轨迹计算基于以下前提[15- 18]:(1)稻飞虱顺风迁飞;(2)稻飞虱起飞迁出日节律为“晨昏双峰型”,清晨起飞峰始于晨光始时止于日出,此期迁出虫量少,峰期短。傍晚起飞峰始于日落后,到昏影终时结束,此间迁出量大,持续20—30 min左右;(3)稻飞虱空中飞行的低温阈值为16.5 ℃,当高空气温低于该值时停止飞行;(4)稻飞虱飞行持续时间不超过36h。回推轨迹以降虫区为起点,在上灯峰日06:00逐时回推至白背飞虱迁出时刻。将得到的轨迹点坐标用Visual FoxPro 9.0转换处理制成属性数据库,再导入ArcGIS 10.0中与地图叠加后成图显示。有效轨迹判定标准:虫源地轨迹终点落在稻区且存在起飞虫源及水稻生育期适合稻飞虱的起飞迁出。
1.8 大气动力背景模拟与分析
利用GrADS 2.0平台提取WRF模式模拟出的高分辨率气象要素数据中不同高度层的水平流场、垂直速度、温度场等要素,结合灯下峰日逐时降水数据对相应站点做天气背景的诊断分析。
2 结果与分析
2.1 湖南省稻飞虱的前期迁入
利用湘阴、湘乡、桂阳、临澧、洪江等若干站点13a(2000—2012年)的前期(3—6月)迁入灯诱数据进行分析始见期、峰次、迁入虫量的变化。其中湘乡2001年灯诱数据与桂阳2011年灯诱数据缺失。始见期先进行数值化处理,将日期转化为以3月20日为起始的气候学日序。峰次以峰日数来表示,以3—4月份上灯虫量5头以上,5—6月份上灯虫量50头以上的天数来表示。
2.1.1 始见期
湖南稻区白背飞虱平均始见期为4月下旬至5月上旬。通过比较2000—2012年间的始见期(图2)湘北的湘乡、湘阴及临澧的平均始见期比湘南的桂阳、湘西南的洪江迟10—20 d,2000—2005年大部分站点始见期迟于平均始见期;2006年后湖南站点白背飞虱始见期均相应有所提前,2007年全国大发生时5个站点提前10—25 d,2010年洪江始见期提前达30 d。2012年3月上中旬,广西沿海稻区即出现灯诱高峰,而湖南各站点始见期与往年平均值趋于一致,并未明显提前,2012年湖南省白背飞虱前期大规模的迁入数量之大并不是受始见期的影响。

图2 2000—2012年湖南白背飞虱前期迁入种群灯诱始见期逐年距平Fig.2 Annual anomalies form the year 2000 to 2012 for the first appearance time of WBPH light-trap catches in Hunan
2.1.2 前期迁入上灯峰日数
统计2000—2012年3—6月白背飞虱上灯峰日(图3),3、4月份白背飞虱乘西南气流可达湘南(主要是湘西南)。除桂阳外,4个站点2006—2012年白背飞虱3—6月迁入峰次均有明显增加,其中洪江2007年上灯高峰日达48次,2012年峰次达52次多,创历史新高。
2.1.3 前期迁入量
分析2000—2012年3月20日至5月20日湖南稻区若干县市白背飞虱前期迁入种群累计灯诱资料,结果表明,3月下旬至5月上旬,迁入种群的主要落点基本沿湘西山地分布,较大的灯诱峰次主要位于山间谷地的迎风坡下,迁入量有着明显的地域差异(图1,图4)。如地处湘西山区的洪江和芷江2012年3—6月总上灯虫量分别为14740头、93019头;而湘南的道县、桂阳、宜章三县同期总上灯虫量6552头,东安为459头(始见期极早,3月27日)。从2000—2012年3—6月湖南白背飞虱前期迁入种群上灯虫量的年际变化(图5)可看出,13年来大部分站点3—6月白背飞虱上灯虫量呈上升趋势,洪江尤为明显,从2006年3—6月累计上灯虫量的1万头至2012年的5万头;再如湘阴2008年3—6月累计上灯虫量1414头,而2012年增至2825头。总之,2012年白背飞虱前期虫源迁入量达到新的高峰,加大了湖南稻区白背飞虱的虫源基数。
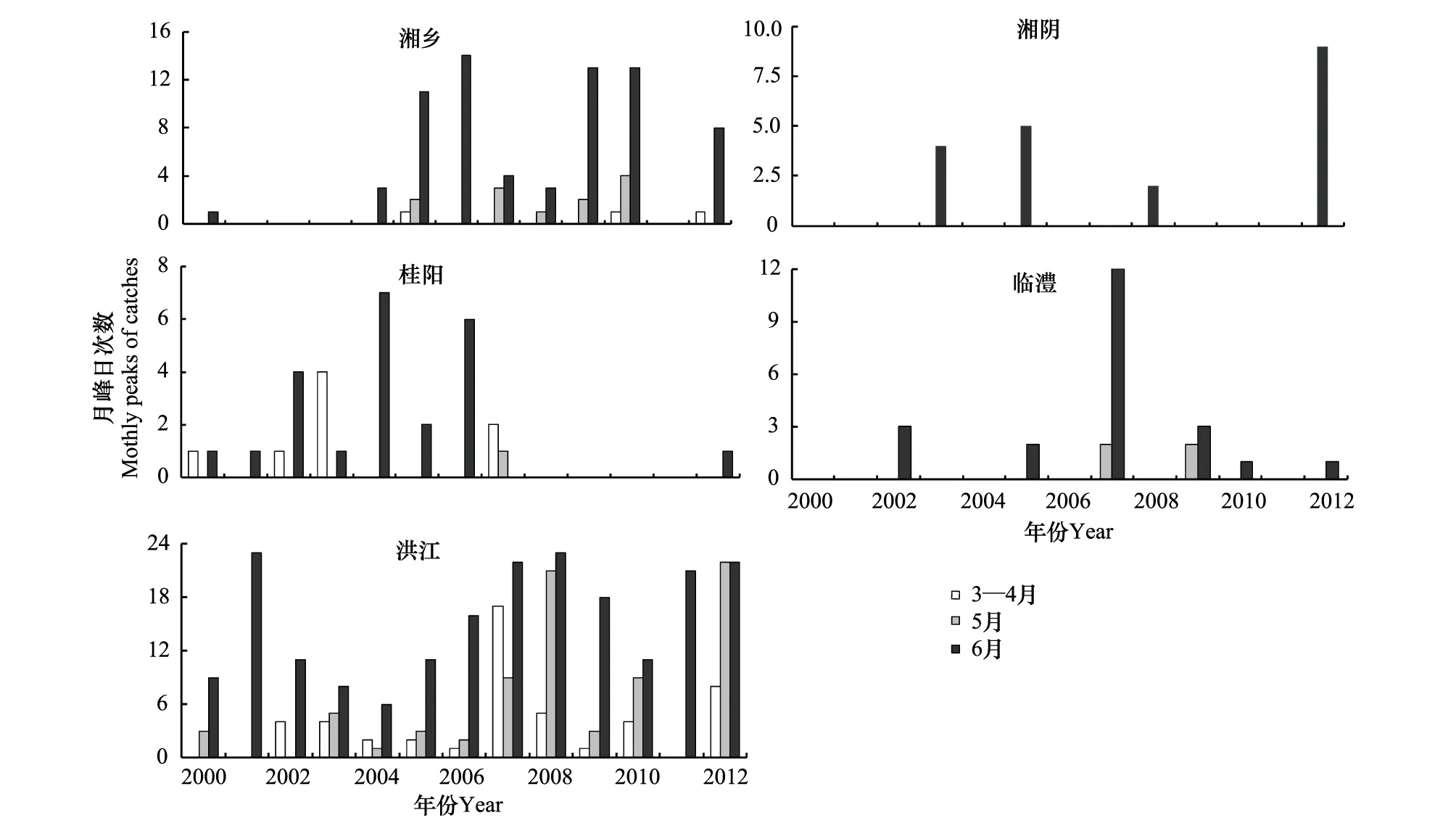
图3 2000—2012年3—6月湖南白背飞虱每月上灯峰日Fig.3 The monthly peaks of WBPH from March to June during the year 2000 to 2012 in Hunan province
2.2 前期迁入种群中小尺度虫源地分析
取湖南省各站点2012年4月份单日上灯虫量大于100头、5月份大于1000头的日期为上灯高峰日,大于10000头作重点分析(表2)。
2.2.1 2012年4月下旬
湘西的洪江4月24日与26日出现白背飞虱灯诱高峰,其回推轨迹显示(图6):逐时回推12h以内的轨迹终点落在广西北部,广西北部稻区单季早稻一般在3月20日至4月5日前播种,单季中稻在4月20日至5月10日前播种,位于“湘桂走廊”东端的兴安4月下旬正值插秧期,因此广西北部稻区不可能为洪江提供迁入虫源。逐时回推13—24h轨迹落点在广西西南部以及越南北部。4月下旬广西北部湾稻区水稻处于分蘖—拔节期,越南北部红河三角洲的冬春稻处于拔节—孕穗期,正是白背飞虱的大量迁出期,可为洪江提供迁入虫源。
安仁位于湖南省东南部。4月29日与30日,以安仁为起点逐时回推13—24h的轨迹终点落在广西西南部及雷州半岛,逐时回推25—36h的轨迹终点落在海南岛东北部地区,落在海中的终点为无效落点(图6)。
龙山4月24日与29日出现上灯峰,逐时回推13—24h的回推轨迹终点落在贵州西南与广西西北接壤地区,落点覆盖地区为西南单季稻稻作区之一,此时尚未插秧,无法为龙山提供迁入虫源。4月24日和29日逐时回推25—36h轨迹终点分别落在越南北部和广西西南地区(图6)。结合越南北部以及广西西南的水稻生育期与田间主虫态,并考察白背飞虱长翅成虫数量及其迁出期,两稻区均为有效虫源地。
4月27日与29日,芷江逐时回推25—26h回推轨迹落点只有较少终点落于广西北部湾以及越南北部(图6)。

图4 2000—2012年3月20日至5月20日湖南白背飞虱前期迁入种群累计灯诱虫量及降虫分布Fig.4 Accumulated number of light-trap catches and distribution of landing of WBPH from 20 March to 20 May 2012, in Hunan
2.2.2 2012年5月上旬
如图7所示,5月3日,湘南的蓝山逐时回推13—24h的轨迹落点分布在广西东南部。5月6日江华和蓝山为起点的逐时回推24h以内终点落在广东西南雷州半岛,回推25—36h终点仅有少部分分布在海南稻区。5月7日,蓝山回推12h以内的轨迹终点落在广东西南雷州半岛。湘中邵东5月7日回推24h以内的轨迹落点可达广西南部。武冈5月8日逐时回推25—36h轨迹终点落于广西西南以及越南北部稻区;5月10日回推13—24h终点落在广西西南部稻区(图7)。
以湘西洪江为起点5月6日逐时回推36h内的轨迹终点仅有较少终点落在广西南部;5月7日,逐时回推13—24h轨迹终点位于广西西南部和越南北部;5月9日,逐时回推24h以内轨迹终点分布在广西西部及越南北部,同日芷江回推13—24h轨迹终点落于广西西南部至越南北部;5月10日,洪江逐时回推13—24h轨迹终点落在广西西南部与越南北部交界处(图7)。

表2 2012年湖南省白背飞虱主要迁入峰次与虫量Table 2 Main immigration peaks and quantity of WBPH in 2012, in Hunan

图5 2000—2012年3—6月湖南白背飞虱迁入种群上灯虫量的年际变化Fig.5 The annual fluctuation of the light-trap catches of WBPH from March to June during the year 2000 to 2012 in Hunan

图6 2012年4月下旬湖南白背飞虱前期迁入种群回推轨迹Fig.6 Endpoints of backward trajectories of primary immigration of WBPH on late April 2012, in Hunan
湘西龙山5月7日逐时回推13—24h轨迹终点于云南东南部和越南北部交界地区(图7);5月9日,逐时回推25—36h轨迹终点较少,落在越南北部以及老挝北部与越南接壤的地区(图7);5月上旬,越南北部地区水稻正处于孕穗—齐穗期,可为湘西提供有效虫源;而老挝东北部乃层峦叠嶂的高原山脉且无旱季水稻种植,故为无效虫源地。

图7 2012年5月初湖南白背飞虱前期迁入种群回推轨迹Fig.7 Endpoints of backward trajectories of primary immigration of WBPH on early May 2012 in Hunan
统计上述白背飞虱前期迁入种群有效虫源地的概率分布(表3),2012年4月下旬至5月上旬湘西(洪江、芷江、龙山、武冈)的迁入虫源主要来自广西西南部以及越南北部,少部分峰次虫源来自广西东南部与广东南部;湘中腹部(邵东)的迁入虫源主要来自广西东南部,广西西南部次之;湘南(蓝山、江华)与湘东(安仁)的迁入虫源主要来自广东南部(主要为雷州半岛稻区)及广西东南部,极少部分峰次虫源来自广西西南部与海南。
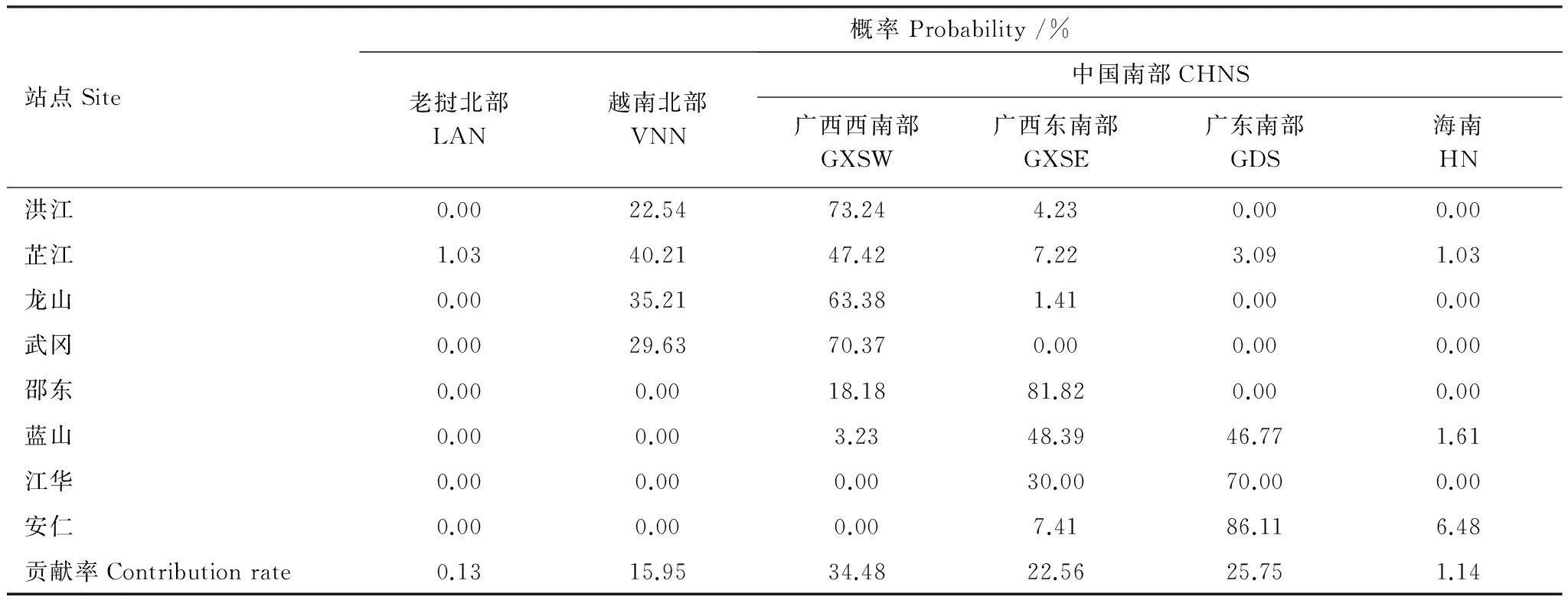
表3 2012年4月下旬至5月上旬湖南稻区白背飞虱中小尺度有效虫源地分布概率Table 3 Probability distribution of valid mesoscale source areas of WBPH in late April to early May of 2012, in Hunan
2.3 降落机制
2.3.1 风切变和降水
4月24日20:00(GMT+8,下同),湖南大部分地区受冷空气影响(图8),自西南而上的暖湿气流与冷空气交绥过境。4月25日02:00,低层切变线南移至湘桂交界处(图8),龙山、芷江以及洪江于冷锋后,洪江4月24日20:00至25日06:00累计降水8.55mm(图9),当夜出现白背飞虱灯诱高峰(表2)。27日20:00,芷江850hPa处出现风切变(图8),28日02:00切变线消失(图8)。此期间27日21:00—23:00,芷江累计降水达8.25mm(图9),出现一个降虫峰(表2)。29日20:00至次日06:00,龙山、芷江、安仁一线出现气流的辅合与风向切变(图8),促使白背飞虱空中聚集并遇雨降落(图9)。5月8日20:00,龙山出现切变线(图8),结合逐时降水图9,其中龙山出现短时强降水过程,该地在20:00至21:00两小时累计降水11.6mm,洪江9日04:00至06:00,洪江出现两次强降水过程,空中的白背飞虱遇雨而集聚降落(图9,表2)。东北冷涡南伸发展、低层切变线南移带来的降雨过程是湖南西部稻区多个站点白背飞虱“同期突发”的主要因素。2012年5月,湖南全省平均降雨量426.5 mm。频繁的降水加之各中小尺度虫源地白背飞虱种群的倍增,进而带来了2012年5月份较大的迁入虫峰。
2.3.2 低温屏障
2012年3月湖南省大部分地区月平均温度低于15 ℃,4月份湘南平均温度达21 ℃,湘北则为19.5 ℃。当气温低于白背飞虱飞行阈值(16.5 ℃)时,迁飞中的种群会被迫降。随着纬度的增加,天然的低温屏障是4月迁入峰主降湘西南的主要原因。
2012年5月3日20:00,蓝山、江华同处切变线南缘,根据湖南稻区800hPa高空温度场分析(图10),2012年5月4日03:00,800hPa高空上低于16 ℃的等温线形成一道低温屏障使空中虫群无法穿越低温区进入湘中腹地,因此在位于16℃等温线南缘的蓝山形成单灯单晚8070头上灯的大规模降虫,而过境区江华则无上灯峰。5月9日02:00,乘西南气流迁飞至湘北稻区上空的白背飞虱种群遇低温屏障被迫降落,龙山恰处16℃等温线南缘(图10),当夜的降水加之低温屏障使龙山形成多达3万多头的灯诱高峰。
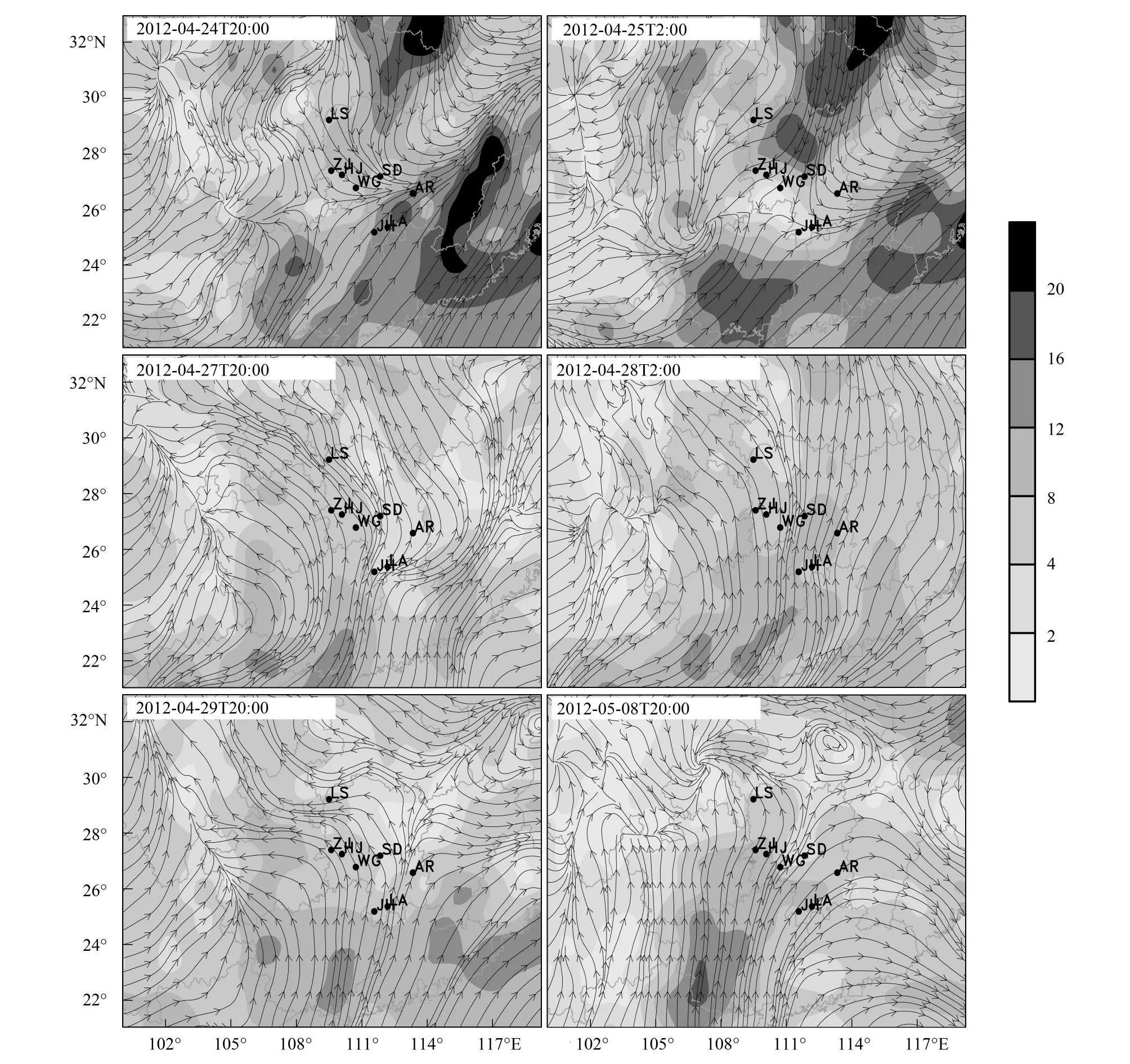
图8 2012年4月下旬至5月上旬850hPa水平流场与风速(站点缩写见表2)Fig.8 Winds field at 850hPa at 20:00 from late April to early May 2012 (Location acronym is showed in Table 2)
2.3.3 下沉气流
4月26日与5月4日,洪江出现灯诱高峰(表2)。分析洪江上空750hPa垂直速度场(图11),可知这2天当晚20:00与次日02:00,洪江上空750 hPa气流均为较明显的下沉运动。另据时间—高度垂直速度剖面(图12),可知这2夜750 hPa以下高度均为下沉气流,其中26日最大绝对垂直速度达0.6 Pa/s,5月4日为0.3 Pa/s。
芷江5月12日、16日及17日均出现了灯诱高峰(表2),通过对芷江上空750hPa垂直速度场(图13)与时间—高度垂直速度剖面(图14)分析,可得这3夜750 hPa以下高度为一致的下沉气流,其中16日、17日下沉气流强度明显,最大绝对垂直速度分别为0.4 Pa/s与0.8 Pa/s,这对此次空中的白背飞虱的集聚降落非常有利。强下沉气流是这几次白背飞虱迫降的决定性因素。

图9 湖南省各站点2012年4月下旬至5月初20:00至次日06:00降水日逐时降水量Fig.9 Hourly precipitation data from 2000 BST to 0600 BST the next day at each site in late April and early May 2012, in Hunan

图10 2012年5月4日03:00(GMT+8)800hPa温度场,5月9日02:00(GMT+8)800hPa温度场Fig.10 Temperature fields at 800hPa at 03:00 BST on 4 May and at 02:00 BST on 9 May, in 2012
2.3.4 地形与垂直流场
湖南为独特的“马蹄形”盆地地貌,东、南、西三面山地环绕,中间为低缓的丘岗,北部则是洞庭湖平原(图1)。芷江与洪江海拔高度差异不大,两站点地形高度约450 m,处山谷地段(图15)。白背飞虱4月至5月迁飞高度为500—1500 m,因此山谷间的“峡谷风”可使虫群集聚,当遇到降水、下沉气流等条件时降落。
4月24日洪江出现白背飞虱灯诱高峰(表2)。4月24日夜间,在沿27.325°N、109°E—110.5°E区域内,气流由西向东移动,芷江为背风坡,洪江相应地为迎风坡(图15),21:00芷江上空气流并无明显下沉现象,表明小尺度背风坡(坡向较缓)的地形下降并不会导致气流下沉,而洪江上空正处下沉气流区,并伴有不明显的垂直环流圈,表明850 m高度的地形可对气流产生阻碍作用。而且,4月24日21:00洪江便有少量降雨,23:00至4月25日02:00共4h累计降水达8.25 mm(图9),垂直环流的聚集、气流的下沉运动与降水的共同作用下,洪江当夜形成上千头的灯诱高峰。
4月26日21:00,110.0°E—115.0°E区域内的经向环流剖面上垂直速度ω分布显示气流在垂直方向上的强烈下沉运动(图16),这一区域750 hPa垂直速度可达0.2 Pa/s以上(图11)。气流强烈下沉后向西运动,洪江(110.09°E;27.25°N)相应地处于背风坡,气流沿着背风坡下沉;而芷江(109.60°E;27.40°N)相对洪江正处迎风坡,由于气流的强烈运动并没有遭遇地形的阻塞作用,气流遇芷江一侧沿山体爬升,形成上升气流,最终在109.5°E—110.5°E区域形成一个纬向垂直环流圈,并于21:00—23:00在洪江两小时累计降水0.45 mm(图9)。轻微少量的降水对空中虫群的迫降作用并不显著,因此26日夜间垂直环流的聚集作用以及背风坡气流的下沉运动是导致洪江灯诱高峰的主要原因。
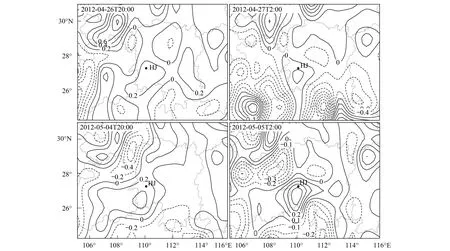
图11 2012年洪江前期迁入白背飞虱种群上灯高峰日750hPa垂直速度场(Pa/s) Fig.11 Pressure vertical velocity field (Pa/s) at 750hPa in the days with large light-trap catches of Hongjiang WBPH in early migration period in 2012
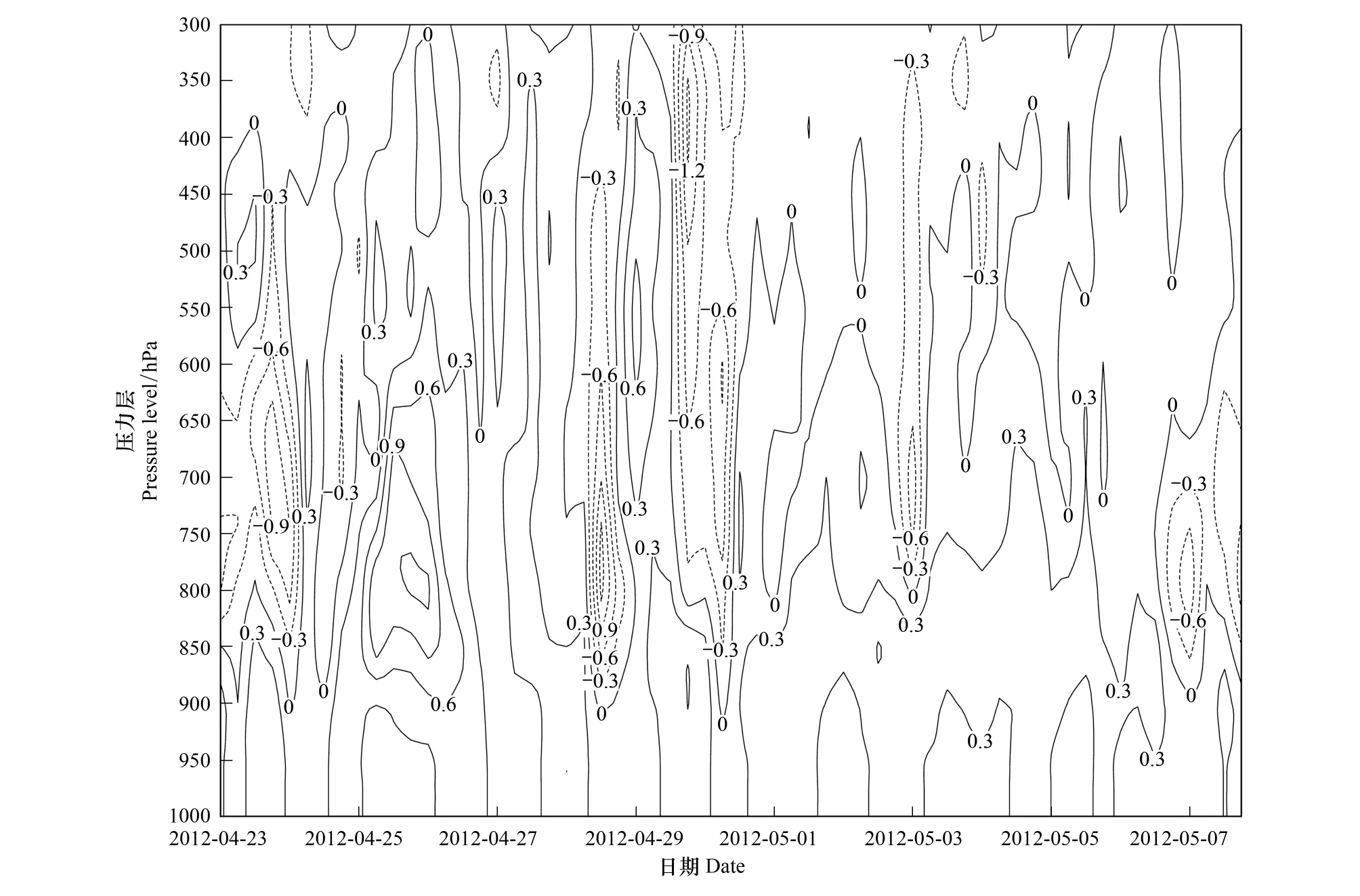
图12 2012年4月23日00:00(UTC)至5月7日18:00(UTC)洪江气压垂直速度(Pa/s)的时间-高度剖面 Fig.12 Time-Altitude cross section of pressure vertical velocity field (Pa/s) of Hongjiang from 00:00 UTC on 23 April to 18:00 UTC on 7 May 2012

图13 2012年芷江前期迁入白背飞虱种群上灯高峰日750hPa气压垂直速度场(Pa/s)Fig.13 Pressure vertical velocity field (Pa/s) at 750hPa in the days with large light-trap catches of Zhijiang WBPH in early migration period in 2012

图14 2012年5月8日18:00(UTC)至5月18日18:00(UTC)芷江垂直速度(Pa/s)的时间-高度剖面Fig.14 Time-Altitude cross section of pressure vertical velocity field (Pa/s) of Zhijiang from 18:00 UTC on 8 May to 18:00 UTC on 18 May in 2012
4月27日21:00,108.5°E—109.7°E区域内的纬向垂直环流剖面上,800—300 hPa处气流有强烈的上升运动,800 hPa以下气流受地面摩擦力的作用,到4月28日03:00形成一个纬向环流圈(图17)。由图9可得,4月27日20:00至22:00,芷江两小时累计降水量达4.25 mm,空中虫群途经该地时随垂直环流聚集并遭遇降水及其下沉气流而降落。而109.8°E—110.2°E区域存在气流轻微的向东辐散,于洪江所处的迎风坡的阻塞作用形成一个风速很小的涡旋,因此洪江当夜并未形成灯诱峰。
龙山(109.50°E;29.22°N)海拔高度比芷江高约200 m(图18),地处湘西北的武陵山脉南侧迎风坡,山脉主体最高海拔高度在1200 m左右,但是这种海拔高度并不会从地域空间上完全阻碍白背飞虱的迁飞。4月29日22:00,迎风坡山麓由于地形阻塞和地面摩擦作用,加之夜间山体冷却辐射,气温迅速降低,而同一高度山谷上空的大气冷却较慢并沿山坡下滑,下层风由山坡吹向山谷并在龙山形成小型垂直涡旋(roller),即由于山体阻塞、地面摩擦及夜间垂直涡旋的作用而使空中虫群集中迫降,使该地出现116头灯诱高峰。与此同时,芷江正处上升气流区,强烈的上升气流带来一次短时强降水过程(19:00至22:00,其中22:00的1h累计降水量达19.1 mm)(图9),使空中虫群大部分迫降于芷江(1256头上灯),少量随气流继续迁至龙山后又遇下沉气流而降落(116头上灯)。
从上节分析来看,龙山地处山体迎风坡下,夜间易受山风的影响而导致虫群降落。但在5月8日19:00,迎风坡下的下沉气流并非是由于地形引起的,而是缘于对流层中上层空气强烈的下沉运动,气流遇地面阻塞,形成小型垂直涡旋,可进一步加深空中虫群的聚集作用。结合5月8日19:00—22:00,龙山3h累计降水达23.49 mm(图9),此次强降水造成龙山上空大气强烈的下沉运动并形成涡旋(图19),加之5月9日02:00—05:00降水带来的降温使龙山位于16℃等温线南缘共同导致虫群迫降,5月8日龙山灯诱虫量达3万多头。
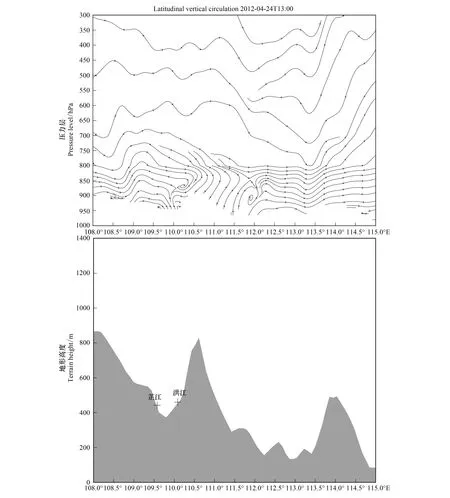
图15 沿27.325°N纬向地形(阴影部分)和2012年4月24日13:00(UTC)沿27.325°N纬向垂直环流Fig.15 Topography (bottom, shaded) along 27.325°N (Bottom), and Latitudinal vertical circulation along 27.325°N at 13:00 UTC on 24 April 2012矢量线,ω扩大50倍;ω: 垂直风速(m/s)

图16 2012年4月26日1300(UTC)沿27.325°N纬向垂直环流 Fig.16 Latitudinal vertical circulation along 27.325°N at 1300 UTC on 26 April 2012 矢量线,ω扩大50倍;ω: 垂直风速(m/s)
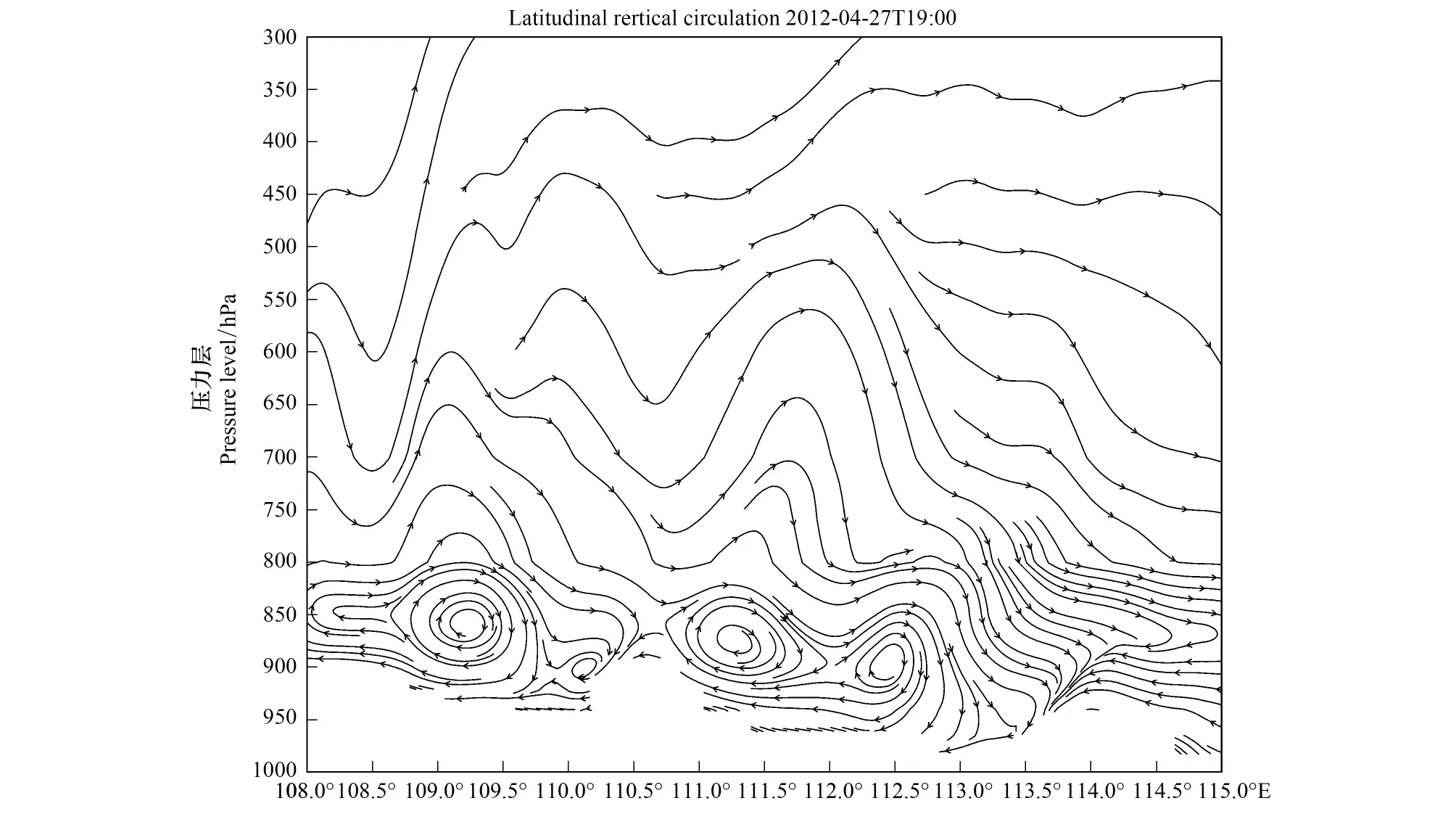
图17 2012年4月27 日 1900(UTC)沿27.325°N纬向垂直环流 Fig.17 Latitudinal vertical circulation along 27.325°N at 1900 UTC on 27 April 2012矢量线,ω扩大50倍;ω: 垂直风速(m/s)
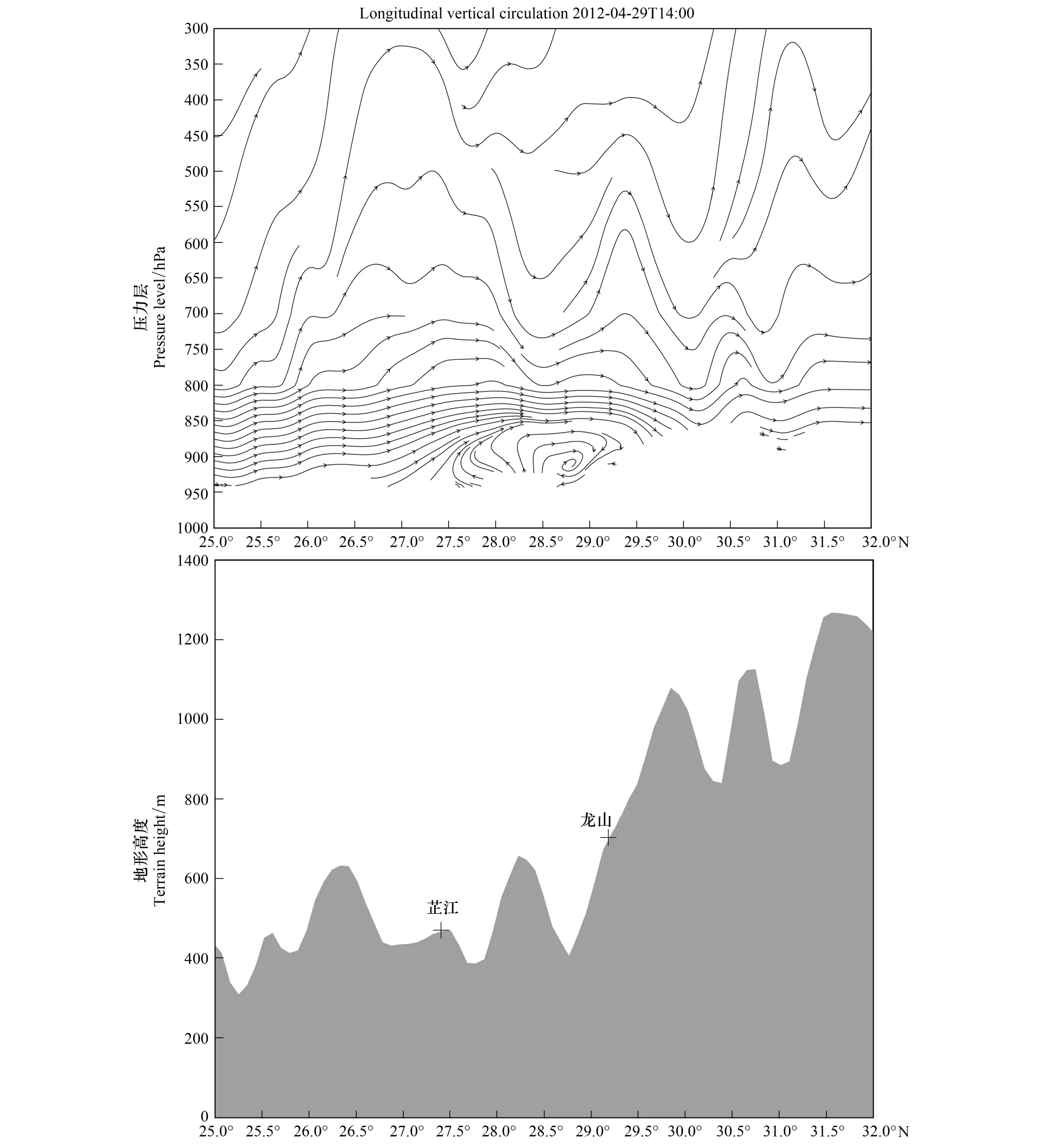
图18 沿109.55°E经向地形(阴影部分)和2012年4月29日1400(UTC)沿109.55°E经向垂直环流Fig.18 Topography (bottom, shaded) along 109.55°E, and Longitudinal vertical circulation along 109.55°E at 1400 UTC on 29 April 2012 矢量线,ω扩大50倍;ω: 垂直风速(m/s)

图19 2012年5月8日1100(UTC)沿109.55°E经向垂直环流Fig.19 Longitudinal vertical circulation along 109.5°E at 1100 UTC on 8 May 2012 矢量线,ω扩大50倍;ω: 垂直风速(m/s)
3 结论与讨论
本文通过对2012年湖南稻区白背飞虱前期迁入虫源地的轨迹分析,对特殊地形条件影响白背飞虱区域性降落的精细化分析,在中小尺度上阐释了白背飞虱迁入事件天气学背景和降落机制,为白背飞虱前期迁入预测和因地制宜的治理提供一定的理论依据。
3.1 湖南稻区白背飞虱前期迁入主要中小尺度虫源地为越南北部、广西西南部
本文使用WRF模式做了2012年湖南省白背飞虱前期迁入种群中小尺度虫源地及降落机制的个例分析。WRF模式有更为先进的数值计算和资料同化技和更为完善的物理过程(尤其是对流和中尺度降水过程),可以更好的模拟天气系统和降水落区[19],显著提高时空分辨率。同时,在轨迹计算程序中加入了昆虫飞行行为参数,进一步提高了轨迹计算的精度与准确性。结果表明,湘西稻区虫源主要来自越南北部、广西西南部;湘南、湘东稻区虫源主要来自广东南部,部分来自广西东南部及海南。
越南位于中南半岛东部,面积约330,000 km2,其中耕地面积占9,350,000 hm2,水稻面积约7,400,000hm2。越南水稻种植主要集中在湄公河平原与红河平原,其中湄公河平原水稻面积和产量占全国面积、产量的50%以上[20]。每年可收获2—3季水稻,北方种植冬春稻和晚稻,以河内为代表,冬春稻在12月和第二年的1月播种,晚稻在6 月中旬播种,南方一年三季稻[21,22];西南毗邻越南的广西与我国海南省隔海相望,是我国南部重要的双季稻种植区,常年4月下旬广西北部湾稻区水稻处于分蘖—拔节期;广东中南稻作区位于广州以南的中南部沿海平原和丘陵区,稻田面积约占全省的46%,是广东省稻谷的最主要产区。 广东西南稻作区位于雷州半岛,每年3月,以雷州半岛为代表的广东南部稻区早稻部分秧苗已经移栽,白背飞虱陆续出现零星迁入[23]。该稻区春播最早,秋收较迟,以双季稻为主。2012年3月上旬,我国南部沿海稻区已出现迁入峰,且迁入量大,迁入次数频繁,为后期稻飞虱的迁出提供了虫源保证。
湖南属亚热带季风湿润性气候,多雨期与夏季风的进退密切相关。850 hPa风场分析表明,湖南白背飞虱前期迁入事件与此期偏南风的盛行和越南北部、广西南部、广东南部以及海南稻区的水稻栽培与生育期有关。因此,通过对越南北部、广西西南部、广州西南部以及中国海南等虫源地的虫情演化和迁出气象条件的分析判断,可为湖南省白背飞虱发生的异地预测与田间防控提供重要依据。
3.2 湖南稻区白背飞虱迁入量与降落落点分布因地形差异明显
山区地形对稻飞虱的“迁飞—降落”过程有着极大的影响[5],携载稻飞虱的气流对地形障碍的响应可由3个因素决定:气流的稳定性、气流速度、山体地形特征[24]。山脉的阻滞使空气被迫抬升并形成绕流和穿谷流以及不同地形的不同加热作用和摩擦效应产生的中尺度环流对山区局地强对流天气有明显强化作用[25];同时,当气流通过山谷时,地形阻塞作用与地面摩擦力及夜间形成的山风使迎风坡的山麓形成垂直涡旋和强劲的峡谷风。山谷间峡谷风的这种“狭管效应”和迎风坡上垂直涡动的“转子效应”使得乘风而来的稻飞虱在很短时间内形成大规模集聚并集中降落。本研究结果表明,湖南地势复杂,河川多呈南北走向,山脉以东西走向为主。从地形上看,南岭山脉横亘在湖南和两广的边界上,其西段诸岭与呈东北—西南走向的雪峰山和武陵山构成“川”字型分布,湘中呈平坝小盆地及湘西南为喇叭口式地形。这种地理特点造就了湖南白背飞虱虫源地的分布差异,而且湘西较大的灯诱峰次主要位于“湘桂走廊”的东端谷地或者是山体的迎风坡,即白背飞虱前期迁入种群在湖南的主要落点基本沿湘西武陵山区至湘南雪峰山区一带稻区分布。这是由于每年4—5月盛行的西南气流通过“湘桂走廊”把广西西南部甚至越南北部的虫源输送到湘西,同时,少部分南向风和东南风可把广东南部与海南稻区的虫源输送到湘东南与湘中。春季西南气流逐步加强,湘东南由于位置偏东,加之南岭高山的阻挡,该区迁入峰次与迁入量较湘西要少得多。
从更大尺度上看,与湘西构成大范围同期突增区的地域还覆盖了整个云贵高原的山地稻区,包括滇东南、黔南黔东南、桂西北、川东南、渝南渝东南和鄂西南。胡国文等[26]的研究表明,贵州东南部和川、黔、湘交界地区(即大娄山东侧)迁入期比我国东部同纬度地区早得多且迁入量亦大得多,是由于该地处于大娄山和雪峰山之间的槽形地带,春季从广西西部北上的西南气流通常携虫群沿云贵高原的东南缘向北伸展到该区。而四川盆地四面环山,由于南部的云贵高原的阻挡作用,使底层西南气流不能进入盆地,因而春季西南气流多见时,四川盆地并不能经常见虫。陈德茂[27]对贵州稻飞虱的研究发现,包括铜仁、黔东南自治州大部分县在内的黔东为白背飞虱、褐飞虱常发区,海拔大多在300—800m,稻飞虱一旦爆发多成片受害。又由于本区地处云贵高原向湘鄂丘陵过渡的边缘,苗岭山脉和雪峰山之间的槽带成了稻飞虱迁飞的走廊,以致该区旬迁入量、灯下诱捕数量远大于省内其他区域。王泽乐等[28]对重庆“两迁”害虫的发生特点与迁入途径进行了研究,结果表明大娄山脉和武陵山脉在渝南和渝东南形成的峡谷地带为“两迁”害虫迁入重庆的重要通道;其中,来自湘桂的“两迁”害虫虫源经武陵山通道迁入重庆(秀山为第一站),来自云贵的虫源则通过大娄山通道入侵重庆(綦江为桥头堡)。一般而言,空中虫群北迁过程中若遇锋面天气,大部分降虫区位于雨区;若遇副高天气,大部分降虫区位于下沉气流区[29]。通过对湖南省前期迁入白背飞虱种群降落机制的研究可知,山区地貌格局导致湘西低温屏障、下沉气流与降雨频繁发生,造成该区迁入峰次多、迁入量大。暖湿气流容易在中尺度地形迎风坡造成气旋性幅合而形成降水[30],同时,地形也可起改变降水落区和强度的作用[31]。
地形对降水及降虫的影响需要更多的试验研究,通过WRF模式的数值试验,设置不同山地地形高度来模拟24 h降水的变化,并分析垂直速度、涡散度、湿位涡、水汽通量等动力场的变化, 阐明地形对降水的影响及白背飞虱的降落机制。此外,本文只是2012年的个例分析,尚需要通过更多个例的剖析,从中寻求稻飞虱迁飞降落的统计规律。
致谢:全国农业技术推广服务中心病虫测报处提供了灯下诱捕资料和田间系统调查资料;湖南省永州市植保站提供了永州市各县区灯下虫量数据;NCEP /NCAR提供了全球再分析资料;国家气象信息中心提供了逐时降水资料,在此一并致谢。
[1] 青先国. 湖南水稻生产发展的对策与关键技术. 中国稻米, 2013, 19(1): 7- 9.
[2] 黄志农, 张玉烛, 刘勇. 湖南水稻三大害虫致灾原因与防治策略. 作物研究, 2006, 20(4): 315- 317.
[3] 王金辉. 湖南省稻飞虱发生为害特点及综合治理实践. 植保技术与推广, 1998, 18(3): 12- 14.
[4] 全国白背飞虱科研协作组. 白背飞虱迁飞规律的初步研究. 中国农业科学, 1981, (5): 25- 31.
[5] 胡国文, 朱敏, 唐健, 潘群威, 仁泽君, 杨克明. 武陵山区稻飞虱常年大发生的特点及原因剖析. 西南农业学报, 1995, 8(2): 53- 60.
[6] 胡高, 包云轩, 王建强, 翟保平. 褐飞虱的降落机制. 生态学报, 2007, 27(12): 5068- 5075.
[7] 林之光. 地形降水气候学. 北京: 科学出版社, 1995: 407- 407.
[8] http://www.mmm.ucar.edu/wrf/OnLineTutorial/Basics/GEOGRID/ter_data.htm.
[9] Wang W, Bruyère C, Duda M, Dudhia J, Gill D, Lin H, Michalakes J, Rizvi S, Zhang X. ARW: Version 3 Modeling System User′s Guide. NCAR, 2011: 354- 354.
[10] Skamarock W C, Klemp J B, Dudhia J, Gill D O, Barker D M, Duda M G, Huang X Y, Wang W, Powers J G. A description of the Advanced Research WRF Version 3. NCAR Technical Note. NCAR/TN-475. 2008: 125- 125.
[11] http://nfgis.nsdi.gov.cn.
[12] http://rda.ucar.edu.
[13] http://old-cdc.cma.gov.cn/.
[14] 朱艳秋, 廖洞贤. 三维空气质点轨迹的计算. 应用气象学报, 1992, 3(3): 328- 333.
[15] Riley J R, Reynolds D R, Smith A D, Rosenberg L J, Cheng X N, Zhang X X, Xu G M, Cheng J Y, Bao A D, Zhai B P, Wang H K. Observations on the autumn migration ofNilaparvatalugens(Homoptera: Delphacidae) and other pests in East Central China. Bulletin of Entomological Research, 1994, 84(3): 389- 402.
[16] 刘芹轩, 吕万明, 张桂芬. 白背飞虱的生物学和生态学研究. 中国农业科学, 1982, 15(3): 59- 66.
[17] 邓望喜. 褐飞虱及白背飞虱空中迁飞规律的研究. 植物保护学报, 1981, 8(2): 73- 82.
[18] 罗举, 汪远昆, 张孝羲, 翟保平. 白背飞虱的迁飞生物学: 起飞与迁出. 应用昆虫学报, 2011, 48(5): 1202- 1212.
[19] 孙健, 赵平. 用WRF与MM5模拟1998年三次暴雨过程的对比分析. 气象学报, 2003, 61(6): 692- 701.
[20] 吕荣华, 周行, 梁朝旭, 卢焜光. 越南水稻的栽培概况. 广西农业科学, 2004, 35(2):102- 103.
[21] 曾娟, 关瑞峰, 吕荣华, 陈应志, 辛德育, 王标. 越南水稻重大病虫害发生概况及其与中国相关性浅析. 世界农业, 2013, (7): 123- 129.
[22] 王政, 齐国君, 吕利华, 胡芊, 袁维熙. 广东白背飞虱早期迁入种群的虫源地分析. 应用昆虫学报, 2011, 48 (5): 1253- 1259.
[23] 竺可桢. 东南季风与中国之雨量. 地理学报, 1934, (1): 1- 27.
[24] Whiteman C D. Mountain Meteorology: Fundamentals and Applications. USA: Oxford University Press, 2000: 355- 355.
[25] 章名立. 地形对暴雨的影响 // 暴雨文集. 长春: 吉林人民出版社, 1978: 58- 64.
[26] 胡国文, 王毓才, 谢明霞. 我国西南稻区白背飞虱,褐稻虱的迁飞和发生特点. 植物保护学报, 1982, 9(3):179- 186.
[27] 陈德茂.贵州稻飞虱研究(1981—1983年). 贵州农业科学, 1984, (5): 24- 28, 11- 11.
[28] 王泽乐, 王梓英, 刘祥贵, 何永坤, 刘怀, 周天云, 郭继萱, 宿巧燕, 车兴壁. 重庆市稻飞虱、稻纵卷叶螟2009年重发生的特点及原因. 西南师范大学学报: 自然科学版, 2011, 36(1): 83- 87.
[29] 谈涵秋, 毛瑞曾, 程极益, 姚禾芬. 褐飞虱远距离迁飞中的降落和垂直气流、降雨的关系. 南京农学院学报, 1984, (2): 18- 24.
[30] 廖菲, 洪延超, 郑国光. 地形对降水的影响研究概述. 气象科技, 2007, 35(3): 309- 316.
[31] Smith R B. The influence of mountains on the atmosphere. Advances in Geophysics, 1979, 21(3): 87- 230.
Mesoscale source areas and landing mechanisms of the early immigrant white-backed planthopperSogatellafurcifera(Horváth) in Hunan Province, China
WU Qiulin1, HU Gao1, LU Minghong2, WANG Biao3, ZHU Xiuxiu3, SU Fang4, ZHANG Zhonggang5, ZHAI Baoping1,*
1DepartmentofEntomology,NanjingAgriculturalUniversity,KeyLaboratoryofIntegratedManagementofCropDiseaseandInsectPests,MinistryofEducation,MinistryofAgricultureofChina,Nanjing210095,China2NationalAgriculturalTechnicalExtensionandServiceCenter,Beijing100125,China3HunanPlantProtectionStation,Changsha410005,China4HongjiangPlantProtectionStation,Hongjiang418200,China5ZhijiangPlantProtectionStation,Zhijiang419100,China
Recently, continuous outbreaks of the white-backed planthopper (WBPH)Sogatellafurcifera(Horváth) have caused heavy rice yield losses in China. Hunan Province is a hotspot in principal migration pathways, and consequently, the main infestation area for the rice planthopper species; therefore, significant economic damage caused by WBPH outbreaks in this province will directly threaten food security in China. Therefore, a better understanding of the source areas and landing mechanisms of immigrant planthoppers in a mesoscale region such as Hunan Province would be essential for an accurate and detailed offsite forecast based on the hopper occurrence status in the source area. A trajectory analysis method based on atmospheric conditions has been developed for the quantitative simulation of population movements and distributions over a range of temporal and spatial scales. Meanwhile, the Weather Research and Forecasting (WRF) model can not only improve the precision and accuracy of trajectory calculation by providing a meteorological background with high spatial and temporal resolution, but also describe and interpret the WBPH migratory mesoscale process by analyzing atmospheric parameters with its advanced numerical technique. In this study, weather datasets on peak days of light-trap catches of the early immigrant WBPHs in Hunan Province during late April and early May of 2012 were selected to conduct a three-dimensional trajectory analysis and a multifactor spatio-temporal dynamic analysis to simulate the migration processes with the WRF model. The following results were derived from the analysis: (1) From April to May 2012, immigrant populations of WBPH in western Hunan were mainly from northern Vietnam and southwestern Guangxi and partly from southeastern Guangxi and southern Guangdong. Source areas of WBPHs that immigrated to southeastern or central Hunan were traced back to southeastern Guangxi and southern Guangdong and partly to Hainan. In late April and early May, source areas of migrant WBPHs were relatively different between western and eastern Hunan. A prevailing southwest monsoon and obstruction by the Nanling Mountains ensured that fewer early immigrant WBPHs landed in southeastern Hunan than in southwestern Hunan. (2) The source populations of WBPH were carried by the southwest low level jet and then clustered and forced to land by downdrafts, a low temperature barrier, and precipitation, which was caused by a frontal cyclone, shear line, or strong updraft, with the application of a three-dimensional trajectory analysis and meteorological modeling. (3) The landing areas of the early immigrant WBPHs were primarily along the Wuling mountainous region in western Hunan, the Xuefeng mountainous region in southern Hunan, and the eastern end of the Xiang-Gui Corridor. Terrain-forced flows (winds modified or channeled by the underlying complex terrain) are associated with mountainous terrain. Thus, terrain was believed to be a main factor that forced massive numbers of WBPHs to land and immigrate frequently into western Hunan. Most of the immigration events and insect convergence occurred in valleys and on the windward side of mountains owing to the vortex effect caused by mountain blocking and friction, mountain breezes, and ravine winds in a valley.
Sogatellafurcifera; mesoscale source areas; landing mechanisms; WRF model; trajectory analysis
国家973项目(2010CB126200);农业公益性行业科研专项(200903051)
2014- 03- 11;
日期:2015- 04- 20
10.5846/stxb201403110415
*通讯作者Corresponding author.E-mail: bpzhai@njau.edu.cn
吴秋琳,胡高,陆明红,王标,朱秀秀,粟芳,张仲刚,翟保平.湖南白背飞虱前期迁入种群中小尺度虫源地及降落机制.生态学报,2015,35(22):7397- 7417.
Wu Q L, Hu G, Lu M H, Wang B, Zhu X X, Su F, Zhang Z G, Zhai B P.Mesoscale source areas and landing mechanisms of the early immigrant white-backed planthopperSogatellafurcifera(Horváth) in Hunan Province, China.Acta Ecologica Sinica,2015,35(22):7397- 7417.
猜你喜欢
作物学报(2022年11期)2022-08-31
湘潮(上半月)(2021年10期)2021-12-02
海峡科学(2021年6期)2021-08-04
园艺与种苗(2021年6期)2021-07-19
文萃报·周五版(2020年3期)2020-02-14
南风(2019年14期)2019-08-26
中华民居(2018年5期)2018-11-20
岷峨诗稿(2017年4期)2017-04-20
环境昆虫学报(2016年6期)2016-12-27
湖南安全与防灾(2015年10期)2015-03-03
