
甘肃引黄灌区典型荒漠植物种群生态位特征
2015-02-04 08:45靳虎甲纪永福魏怀东周兰萍胡小柯
中国水土保持 2015年7期
靳虎甲,李 亚,纪永福,魏怀东,周兰萍,胡小柯,肖 斌
(1.甘肃省水利厅 水土保持局,甘肃 兰州730000;2.甘肃省荒漠化与风沙灾害防治省部共建国家重点实验室培育基地/甘肃省荒漠化防治重点实验室/甘肃省治沙研究所,甘肃 兰州730070)
(责任编辑 徐素霞)
1917年Grinnell 提出了生态位(niche)概念[1],生态位的研究便成为生态学的一个热点问题。生态位理论不断完善和发展,生态位的概念也先后经历了“空间的生态位”[2]、“功能性的生态位”[3]和“多维超体积生态位”[4]等认识阶段,目前对于生态位的理解趋向于“植物种在多维环境梯度空间的位置”[5]这一表述。在评价种间和种内关系,以及种群在群落中所处的地位时,生态位的研究已经成为一种重要的手段,从而使生态位的研究在生物多样性及其形成机制、群落演替与发展趋势、植物资源保护与利用等方面有广泛的应用前景[6]。
甘肃引黄灌区(景电灌区)是中国西北最主要的引黄灌区之一,横跨甘肃景泰、古浪两县,北倚腾格里沙漠,东临黄河,南靠长岭山,自20 世纪七八十年代开始实施景电一期、二期工程提灌黄河水灌溉,使大片固定沙地变为水浇地,并形成了新的绿洲[7]。总体而言,甘肃引黄灌区的生态环境十分脆弱,天然荒漠植被也存在不同程度的退化,一旦现有天然荒漠植被遭到破坏,必将严重威胁到灌区的农田。本研究选取甘肃引黄灌区天然荒漠植被为研究对象,运用生态位理论,通过对处于不同立地类型下不同恢复演替阶段的群落优势种群生态位进行研究,揭示该区荒漠植被的环境适应性和种间关系、种群动态的变化,阐明植被恢复过程的特征和规律。这一研究对该区退化生态系统功能的恢复和治理具有重要的理论意义和实践意义。
1 研究地区自然概况
研究区位于甘肃引黄灌区北部的腾格里沙漠东南缘,地理坐标为103°41' ~104°50'E、36°14' ~37°20'N,平均海拔1 650 m。该区气候干燥、干旱少雨、风沙大,年均降水量185 mm,集中在7—9月,年均蒸发量3 040 mm;年均气温8.5 ℃,雨热同期;年均风速为3.5 m/s,年大风日数为27.9 d,最大风速25 m/s,年平均沙尘暴日数为21.9 d,最多达47 d,大风多发生在3—5月份,主要以西北风为主。地表水资源较为贫乏,地下水资源矿化度高、水质差。灾害性天气主要有干旱、大风、霜冻、冰雹、干热风等。主要土壤类型为风沙土、灰钙土和荒漠灰钙土。自然分布的植物种类主要有油蒿(Artemisia ordosica)、甘蒙锦鸡儿(Caragana opulens)、猫头刺(Oxytropis aciphylla)、茵陈蒿(Artemisia capillaries)、沙米(Agriophyllum squarrosum)、画眉草(Eragrostis pilosa)、针茅(Stipa capillata)等。
2 样地设置与研究方法
2.1 样地设置
在野外分别选取流动沙丘、半固定沙丘、固定沙丘三种立地类型的典型样地各15 个,其中流动沙丘选取15 个20 m ×20 m 的样方,半固定沙丘和固定沙丘各选取15 个10 m×10 m 的样方,共选取45 个样方。使用全球定位系统(GPS)对调查地点定位,并记录各样地的海拔、地貌和土壤类型等环境因子(表1)。
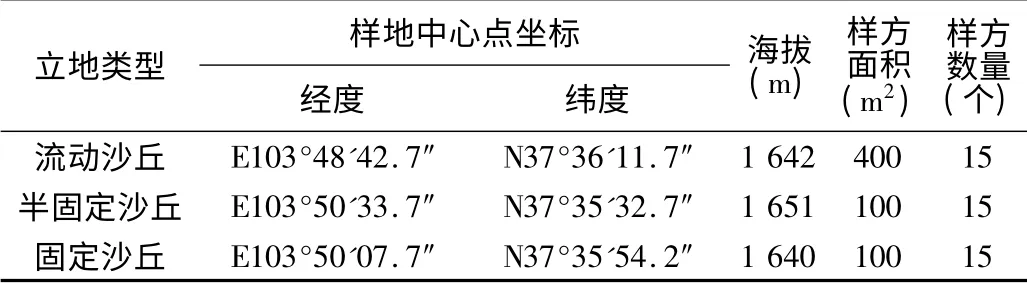
表1 不同立地类型样地信息
2.2 植被调查
植被调查于2011年9月进行,在每个样方内进行植被调查,逐株调查样方内所有植物的种类、数量(株/丛)、盖度和高度等指标。
2.3 数据分析
(1)重要值。在统计各样地植物高度、盖度、密度和频度的基础上,分别按灌木和草本植被计算样方内各物种的重要值,具体的计算公式为[8]

(2)生活型统计。本研究中,分别采用丹麦植物学家Raunkiaer 的生活型分类系统[9]和我国植物学家采用的《中国植被》生活型分类系统对植被类型进行了划分[10]。
(3)生态位宽度。本研究以所调查的样方作为资源状态,样方数目为资源梯度的数目,以重要值为数量指标进行生态位的计测。Levins 生态位宽度指数为[11]
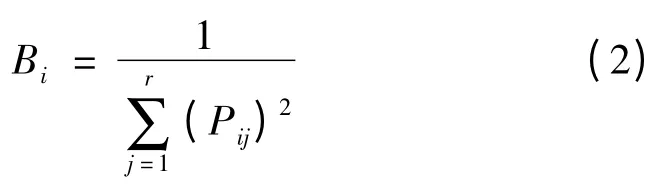
其中
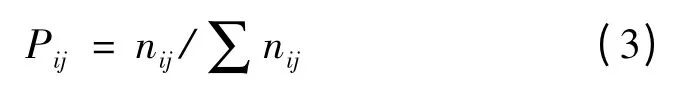
上二式中:Bi为种i 的生态位宽度;Pij代表种i 在第j个资源状态下的资源利用率;nij为种i 在资源梯度级j的数量特征值,本文中为种i 在第j 个样方的重要值;r为资源等级数,本文中为样方数。
(4)数据处理。数据分析处理和图形绘制均使用Microsoft 公司的表格处理软件Excel 完成。方差分析在SPSS13.0 数据处理系统中进行,多重比较用Duncan 新复极差法检验(显著性水平为0.05)。
3 结果与分析
3.1 不同群落的种群生态位宽度
在甘肃引黄灌区的腾格里沙漠南缘,不同立地类型的植物群落中各种群的生态位宽度及其在各群落中的生态位总宽度见表2。

表2 不同立地类型的植物群落中各种群的生态位宽度
由表2 可知:在流动沙丘群落中,砂蓝刺头和刺沙蓬的生态位宽度值较大,其值分别为0.846 6、0.819 9,白刺、沙米、雾冰藜、小画眉草的生态位宽度值也均超过了0.7,表现出了对该群落较强的适应性,而该群落中沙葱、针茅、芨芨草等一些多年生草本的生态位宽度值则比较低。在半固定沙丘群落中,白刺的生态位宽度值最大,已经达到了0.904 7,小画眉草、刺沙蓬、小花棘豆、油蒿、莳萝蒿的生态位宽度值均超过了0.8,而茵陈蒿、砾苔草、无芒隐子草、针茅的生态位宽度值在本群落里就比较低。在固定沙丘群落里,油蒿的生态位宽度值达到了0.973 8,表现出对该立地类型极强的适应性,蒺藜、刺沙蓬、猪毛蒿、砾苔草、猫头刺、针茅的生态位宽度值也均超过了0.9,其适应性也很强,但相对而言,柠条、白刺、沙芥等种群在该群落的生态位宽度值就比较低。就整个区域所有立地类型的生态位总宽度值而言,刺沙蓬的最大,达到1.507 8;油蒿其次,为1.336 8;砂蓝刺头、小画眉草、雾冰藜、蒺藜、白刺、猫头刺、猪毛蒿、小花棘豆的总生态位宽度值也较大,均超过1.0;相对而言,柠条、沙芥、籽蒿、虫实等种群的总生态位宽度值是比较低的。比较不同立地类型中各种群的生态位宽度值的平均值大小,固定沙丘>半固定沙丘>流动沙丘,这也从一个方面说明了不同立地类型上群落对环境的总体适应程度。
3.2 不同生活型种群生态位宽度
根据Raunkiaer 的生活型分类系统,本区域的植物可以分为一年生植物(Th)、地面芽植物(H)、地上芽植物(Ch)、高位芽植物(Ph)四种类型。而根据《中国植被》的生活型分类系统,本区域植物有一年生植物、多年生草本、半灌木与小灌木、灌木这四种类型。
在本区域按照Raunkiaer 生活型分类系统划分的各植被类型的生态位宽度变化如图1 所示。由图1 可知,对于一年生植物的生态位宽度值而言,半固定沙丘>流动沙丘>固定沙丘,但是三者之间的差异性未达到显著水平;地面芽植物的生态位宽度值在固定沙丘上达到0.534 7,显著高于其在半固定沙丘和流动沙丘的生态位宽度值;地上芽植物的生态位宽度值表现为固定沙丘>半固定沙丘>流动沙丘,且三者之间的差异性均达到了显著水平,其中固定沙丘为0.664 2,分别比半固定沙丘和流动沙丘增加了42.35% 和109.40%;高位芽植物的生态位宽度值在固定沙丘和半固定沙丘上分别达到0.605 5 和0.604 4,而在流动沙丘上只有0.485 4,前两者之间差异不显著,而前两者与流动沙丘间的差异性到达了显著水平。
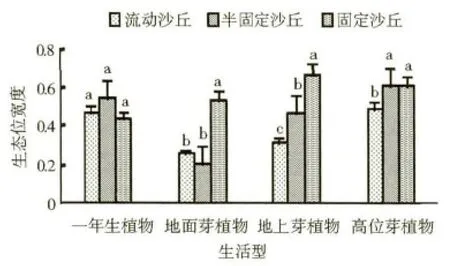
图1 不同立地类型采用Raunkiaer 生活型系统划分的植被类型生态位宽度(p <0.05)
按照《中国植被》的生活型分类系统对研究区的植物进行划分后再进行生态位宽度分析,结果见图2。由图2 可知,其变化特征与按Raunkiaer 生活型分类系统划分的植被类型的生态位宽度变化比较相似,但是具体分析结果略有不同:一年生植物在流动沙丘、半固定沙丘和固定沙丘的生态位宽度值分别为0.468 2、0.557 8、0.436 9,三者之间的差异性未达到显著水平;多年生草本的生态位宽度值在固定沙丘上为0.534 7,而在流动沙丘和半固定沙丘上只有0.256 4 和0.206 5,且固定沙丘与其他两种立地类型上的生态位宽度值差异性达到了显著水平;半灌木与小灌木的生态位宽度值表现为固定沙丘>半固定沙丘>流动沙丘,三者之间的差异性均达到了显著水平,半灌木与小灌木在固定沙丘的生态位宽度值高达0.943 8,是流动沙丘的2.97倍,是半固定沙丘的2.02 倍;灌木在半固定沙丘上的生态位宽度值是最大的,其次是固定沙丘,而流动沙丘最小,三者之间整体差异显著,但后两者之间的差异性未达到显著水平。
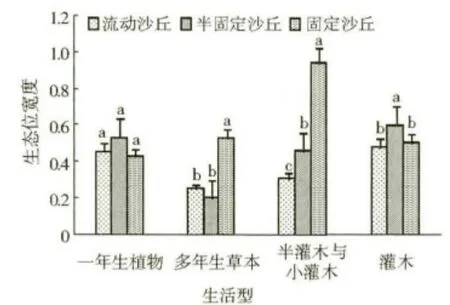
图2 不同立地类型采用《中国植被》生活型系统划分的植被类型生态位宽度(p <0.05)
按Raunkiaer 生活型分类系统划分的植被类型的生态位总宽度大小为地上芽植物>一年生植物>地面芽植物>高位芽植物,且这四种植被类型的生态位总宽度整体之间差异达到了显著水平,但地上芽植物、一年生植物、地面芽植物这三者之间的差异性未达到显著水平,且高位芽植物与地面芽植物的差异性也不显著,而高位芽植物与地上芽植物、一年生植物这两者之间的差异性是显著的(图3)。按《中国植被》的生活型分类系统研究本区植被,其生态位总宽度变化规律与按Raunkiaer 生活型分类系统划分的分析结果比较相似,具体表现为:半灌木与小灌木的生态位总宽度值最大(0.866 8),其次是一年生植物(0.763 3),再次是多年生草本(0.749 1),而灌木的生态位总宽度值最小(0.744 9)。就差异性而言,四种植被类型整体差异显著,但后三者之间的差异性却未达到显著水平(图4)。
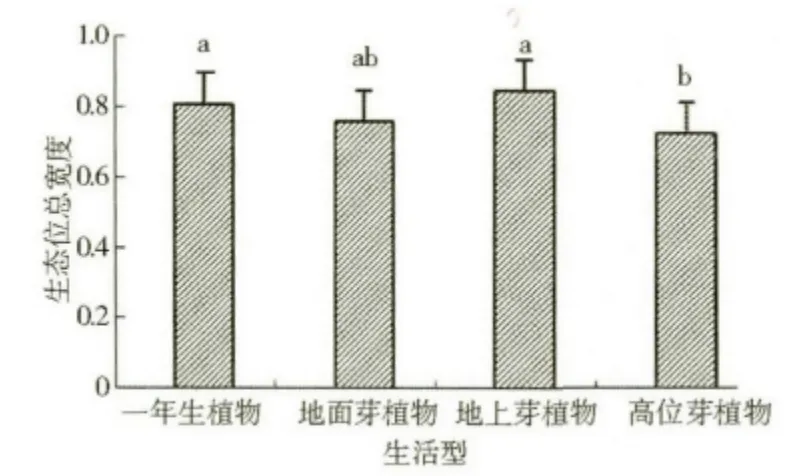
图3 采用Raunkiaer 生活型系统划分的植被类型的生态位总宽度(p <0.05)
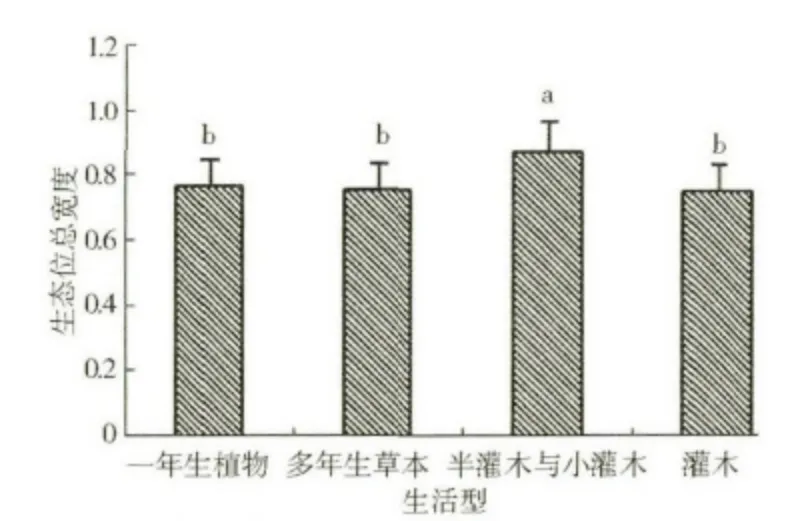
图4 采用《中国植被》生活型系统划分的植被类型的生态位总宽度(p <0.05)
4 结论与讨论
生态位反映了物种对群落内各种生态因子适应的综合结果,即生态位越宽的物种对资源利用越充分、分布越广泛[12]。具有较宽生态位的种群,其一定具有较强的生态适应性,因而生存机会多,分布范围也较广,更倾向于泛化种[13]。对甘肃引黄灌区北侧腾格里沙漠南缘荒漠区内不同立地类型的典型荒漠植物种群的生态位进行的研究,在一定程度上揭示了该区荒漠植被对资源的占有、利用情况及群落发展演替的趋势。
本研究选取的群落是从流动沙丘到半固定沙丘再到固定沙丘,而这样一种空间上的变化实际上从另一个侧面替代了植被在时间上的一种演替序列[14]。在这样一个演替过程中,每一个阶段的群落都有一个指示植物,如流动沙丘的先锋植物沙米[15],半固定沙丘上的白刺[16],固定沙丘上的油蒿、针茅、无芒隐子草等植物[17]。本研究表明,各种群在不同立地类型上的生态位宽度值存在差异,总体而言,在流动沙丘到半固定沙丘再到固定沙丘的演替过程中,一些一年生的流动沙丘指示性植物的生态位宽度值是逐渐减小的,而一些在半固定、固定沙丘上才适宜生长的半灌木与小灌木的生态位宽度值明显增大。不同立地类型上各种群生态位宽度值的平均值大小表现为固定沙丘>半固定沙丘>流动沙丘。沙米在这样一个演替序列中其生态位宽度呈现从大到小的变化趋势,白刺在从流动沙丘到半固定沙丘的演替过程中其生态位宽度是增加的,但到了固定沙丘其生态位宽度又明显减小,而无芒隐子草、针茅、油蒿的生态位宽度又呈逐渐增加的变化趋势。实际上群落的建立和演替过程其核心是物种的更替,物种更替是群落环境演变、物种的环境适应性、竞争作用等种间关系共同作用的结果和集中表现,这几方面的变化是互动的。在流动沙丘上,一些一年生的物种对空间的占用能力很强,有比较大的生态位宽度;而到了半固定沙丘阶段,由于白刺的竞争作用,使得这些先锋植物的生态位宽度出现不同程度的减小;在固定沙丘上,沙地环境有所稳定,加上土壤理化性质的改良,无芒隐子草、针茅等多年生草本与油蒿能比较好地占据该立地环境的资源空间,它们的生态位宽度明显增加。
按Raunkiaer 生活型分类系统划分的植被类型的生态位宽度表现为:一年生植物不同立地类型间的生态位宽度值差异不显著;地面芽植物的生态位宽度值固定沙丘显著大于流动沙丘和半固定沙丘;地上芽植物的生态位宽度值固定沙丘>半固定沙丘>流动沙丘,三种立地类型间差异更加明显;高位芽植物在固定沙丘与半固定沙丘间的差异很小,但与流动沙丘差异比较大。按《中国植被》的生活型分类系统划分的一年生植物、多年生草本、半灌木与小灌木、灌木的生态位宽度值在三种不同立地类型间的大小变化规律,与按Raunkiaer 生活型分类系统划分的植被类型比较相似。群落的生活型提供了群落对特定环境因子的反应、空间利用和种间竞争关系等方面的信息[18]。本研究中地上芽植物与一年生植物的生态位总宽度值比较高,说明这两类植被类型对于本区的环境条件比较适应。地上芽植物占优势反映该地区具有较长的严寒季节,而一年生植物的生态位宽度较大说明一年生植物在本区中占有的优势明显。拥有丰富的一年生植物本身是全球荒漠地区常见的一个特点[19],这也与该区域干旱的气候特征相吻合[20]。
[1]Grinnell J.The niche-relationships of the California Thrasher[J].The Auk,1917,34(4):427-433.
[2]Elton C.Animal Ecology[M].London:Sidgewick and Jackson,1927:63-68.
[3]Berendse F.Interspecific competition and niche differentiation between Plantago lanceolata and Anthoxanthum odoratum in a natural hayfield[J].Journal of Ecology,1983,71(2):379-390.
[4]Altenberg L D M L.Statistical inference on measures of niche overlap[J].Ecology,1985,66(4):1204-1210.
[5]余世孝,奥罗西L.物种多维生态位宽度测度[J].生态学报,1994,14(1):32-39.
[6]程中秋,张克斌,常进,等.宁夏盐池不同封育措施下的植物生态位研究[J].生态环境学报,2010,19(7):1537-1542.
[7]刘瑞,靳虎甲,马全林,等.甘肃景电灌区不同栽植年限枸杞生物量分配特征[J].生态学杂志,2012,31(10):2493-2500.
[8]张林静,岳明,赵桂仿.生态位不同计测方法在绿洲荒漠过渡带上的应用比较[J].生态学杂志,2002,21(4):71-75.
[9]Raunkiaer C.The Life Forms of Plants and Statistical Plant Geography[M].New York:Oxford University Press,1932:2-104.
[10]中国生态系统研究网络科学委员会.陆地生态系统生物观测规范[M].北京:中国环境科学出版社,2007:46-75.
[11]Levins R.Evolution in Changing Environments:Some Theoretical Explorations[M].Princeton:Princeton University Press,1968:61-62.
[12]张峰,上官铁梁.山西翅果油树群落优势种群生态位分析[J].西北植物学报,2004,24(1):70-74.
[13]李斌,李素清,张金屯.云顶山亚高山草甸优势种群生态位研究[J].草业学报,2010,19(1):6-13.
[14]张继义,赵哈林,张铜会,等.科尔沁沙地植物群落恢复演替系列种群生态位动态特征[J].生态学报,2003,23(12):2741-2746.
[15]马全林,张德奎,陈芳,等.流动沙丘先锋植物沙米的种子特征研究[J].种子,2008,27(11):72-76.
[16]靳虎甲,马全林,何明珠,等.石羊河下游白刺灌丛演替过程中群落结构及数量特征[J].生态学报,2013,33(7):2248-2259.
[17]刘瑞,李毅,刘世增,等.干旱区油蒿种群种内竞争研究[J].中国农学通报,2013,29(10):51-57.
[18]郭泉水,江洪,王兵,等.中国主要森林群落植物生活型谱的数量分类及空间分布格局的研究[J].生态学报,1999,19(4):573-574.
[19]李雪华,李晓兰,蒋德明,等.干旱半干旱荒漠地区一年生植物研究综述[J].生态学杂志,2006,25(7):851-856.
[20]赵丽娅,李兆华,李锋瑞,等.科尔沁沙地植被恢复演替进程中群落土壤种子库研究[J].生态学报,2005,25(12):3204-3211.
猜你喜欢
当代水产(2022年8期)2022-09-20
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
河南科学(2020年3期)2020-06-02
诗潮(2019年10期)2019-11-19
中央民族大学学报(自然科学版)(2018年4期)2018-03-19
成才之路(2017年21期)2017-07-27
安徽农学通报(2017年9期)2017-05-19
现代农业科技(2016年22期)2017-03-24
诗潮(2017年2期)2017-03-16
