
白蛾黑基啮小蜂种群数量变化规律的初步研究
2015-02-03 07:37郭同斌宋明辉王虎诚郭加忠
江苏林业科技 2015年2期
关键词:变化规律
吴 静,郭同斌*,宋明辉,杜 伟,王虎诚,郭加忠
(1.徐州市森林病虫防治检疫站,江苏 徐州 221009;2.高邮市京杭运河管理处,江苏 高邮 225600)
白蛾黑基啮小蜂种群数量变化规律的初步研究
吴静1,郭同斌1*,宋明辉1,杜伟1,王虎诚1,郭加忠2
(1.徐州市森林病虫防治检疫站,江苏徐州221009;2.高邮市京杭运河管理处,江苏高邮225600)
摘要:选择徐州市林果场、云龙区和新沂市等受杨小舟蛾危害较重的不同类型杨树林分的未放蜂区(CK)和放蜂区,分别测定白蛾黑基啮小蜂在自然和人工释放2种状态下的种群数量变化。结果表明,该蜂林间自然种群数量以越冬代最高,对杨小舟蛾越冬蛹的寄生率高达(22.2±5.8) %,极显著高于第1~4代蛹(3.4%~6.4%) ;于越冬代和第2代害虫蛹期人工释放该蜂,释放后当代小蜂种群数量明显上升,可显著提高释放当代(越冬代和第2代)害虫蛹的寄生率,并对释放后各代害虫蛹具有一定的持续寄生效果;各代小蜂室温下的羽化进度均与相应世代寄主蛹发育进度相吻合。
关键词:白蛾黑基啮小蜂;杨小舟蛾;种群数量;变化规律
白蛾黑基啮小蜂(Tetrastichus nigricoxae Yang)属膜翅目(Hymenoptera)姬小蜂科(Eulophidae),是杨树人工林食叶害虫——杨小舟蛾(Micromelalopha troglodyta Graeser)蛹期重要的群集性内寄生蜂[1]。目前国内学者对该寄生蜂的生物学、行为学及其繁殖利用技术进行了详细研究[1-5],但有关其种群数量变化规律尚未见报道。为此,笔者对该蜂在自然与林间人工释放2种状态下的种群数量变化规律进行了初步研究,并对室内成蜂羽化与林间寄主化蛹的进度进行了观测,进一步探讨寄生蜂羽化进度与其相对应世代寄主蛹发育进度间的相关性,为利用该蜂生物防治杨树害虫提供依据。
1 材料与方法
1.1自然种群数量调查
由于白蛾黑基啮小蜂除成蜂外,卵、幼虫和蛹3个虫态均在寄主蛹壳内发育,调查其林间种群数量较为困难,因此以调查寄主蛹的被寄生率来代表其林间种群的发生数量。2009年11月至2010年12月,分别选择徐州市林果场杨树林带、徐州市云龙区六堡村古黄河杨树成片林和小韩庄村杨树林网及新沂市草桥镇浪青河岸杨树林带共4种受杨小舟蛾危害较重的不同类型杨树林分,作为白蛾黑基啮小蜂对杨小舟蛾各代蛹自然寄生率调查的固定样地,每块样地按不同区位或方位划分成2个调查小区。调查样地作为放蜂区的对照(CK),不进行放蜂处理,并远离放蜂区300 m以上[该蜂生物学研究结果表明,成蜂飞行扩散距离为1~3 m[2],因此放蜂区人工所释放的小蜂不能飞到上述自然状态的调查样地,因而自然状态(CK区)和人工释放状态的调查地小蜂种群间不存在相互影响]。上述调查样地杨树品种均为Ⅰ-69杨,林龄6~10 a,样地面积分别为1.3,16.7,2.7,6.7 hm2。
于2009年11月杨小舟蛾越冬蛹期,在上述CK区每块调查样地的每个小区内,分别采集杨小舟蛾蛹40~200头(由于第1代蛹数量极少,每个小区采集数量不低于40头,其他各代100~200头,每代共采集样本8个)。将采集的越冬蛹装入铁纱笼内(防止老鼠等其他动物危害),置于表土层,并覆盖稻草越冬保存。翌年4月15日前取出单头,置于室内指形管中,逐日调查羽化为白蛾黑基啮小蜂(出蜂)的寄生蛹数(即可统计出越冬代成蜂羽化出蜂的发育进度,详见1.3节)和同期羽化为日本追寄蝇(Exorista japonica)等其他寄生性天敌的寄生蛹数及羽化为杨小舟蛾成虫(出蛾)的未寄生蛹数。小蜂羽化结束后,对未出蜂也未出蛾的越冬蛹进行解剖,统计未出蜂的寄生蛹数,计算各样本杨小舟蛾越冬蛹的被寄生率(白蛾黑基啮小蜂)和天敌总寄生率(啮小蜂+日本追寄蝇等其他天敌),计算方法参照文献[5]。2010年第1~4代化蛹末期,采集各代杨小舟蛾蛹(无需越冬保存),置于室内室温条件下发育至出蜂(同时统计2~5代成蜂羽化进度。由于第1代小蜂寄生于尚未羽化的杨小舟蛾越冬蛹[5],其数量极少而难以采集,因此对自然和1.2节人工释放状态的第1代小蜂种群数量均未进行调查),统计1~4代寄主蛹的被寄生率和天敌总寄生率,调查统计方法同上。
1.2人工释放后种群数量调查
1.2.1杨小舟蛾越冬蛹期释放2009年10月上、中旬,在云龙六堡和新沂草桥选取2块杨树林,释放白蛾黑基啮小蜂防治杨小舟蛾越冬蛹,放蜂量为30 枚/hm2(每枚柞蚕蛹出蜂1 000~1 500头)。分别于2009年11月和2010年5~9月,采集杨小舟蛾越冬蛹和第1~4代蛹,调查白蛾黑基啮小蜂对越冬蛹的寄生率、天敌总寄生率和对第1~4代蛹的持续寄生效果,采样数量与调查统计方法同上。选择远离上述放蜂区300 m以外的杨树林分,作为未放蜂区(CK)调查该蜂的自然寄生率(详见1.1节)。
1.2.2杨小舟蛾第2代蛹期释放2010年6月,在徐州市林果场选取杨树林带,于下旬释放白蛾黑基啮小蜂防治杨小舟蛾第2代蛹,释放量为75枚/hm2柞蚕蛹(淹没式释放,约相当于林间害虫蛹数量的3倍),同时采用上述方法设立对照区(CK)。分别采集放蜂区和对照区的第2~4代蛹,调查白蛾黑基啮小蜂对第2代蛹的寄生率、天敌总寄生率和同期对照区寄生率及对第3~4代蛹的持续寄生效果,采样数量与调查统计方法同上。
1.3室内成蜂羽化与林间寄主化蛹进度观测
将1.1节各代自然种群数量调查的每块样地所采集2个小区样本中的1个和1.2.1节越冬代放蜂后各代蛹寄生效果调查所采样本,置于室内室温条件下培育至出蜂。逐日观察调查各代寄主蛹被啮小蜂寄生的蛹数,统计在室温条件下越冬代和第2~5代白蛾黑基啮小蜂成蜂羽化进度。2010年5月下旬起,在徐州市林果场杨树林带选取样株15株,在样株地表以干基为中心,铺设直径为2 m的塑料布,布上覆盖麦草。每日6:00~8:00逐日收集、统计塑料布内当日下树化蛹的老熟幼虫和蛹数量,记录天气情况,直至最后1代害虫化蛹结束,统计2~4代幼虫的化蛹进度,分析该寄生蜂各代成蜂羽化与寄主化蛹进度的相关性。
采用SAS统计软件多重比较t检验法检验不同世代寄生率指标间的差异显著性[6]。
2 结果与分析
2.1自然种群数量变化
2009~2010年白蛾黑基啮小蜂对杨小舟蛾各代蛹的自然寄生率和包含日本追寄蝇等其他天敌的
总寄生率观测结果见图1。由图1可知,各代蛹的自然寄生率以越冬代最高,达(22.2±5.8) %,极显著高于其他各代(P<0.01) ;第1~4代蛹的自然寄生率为3.4%~6.4%,相互之间均无显著差异;各代蛹的总寄生率也以越冬代最高,但相互之间均无显著差异。因此,白蛾黑基啮小蜂的林间自然种群数量以越冬代最高,第1~5代种群数量极显著低于越冬代。
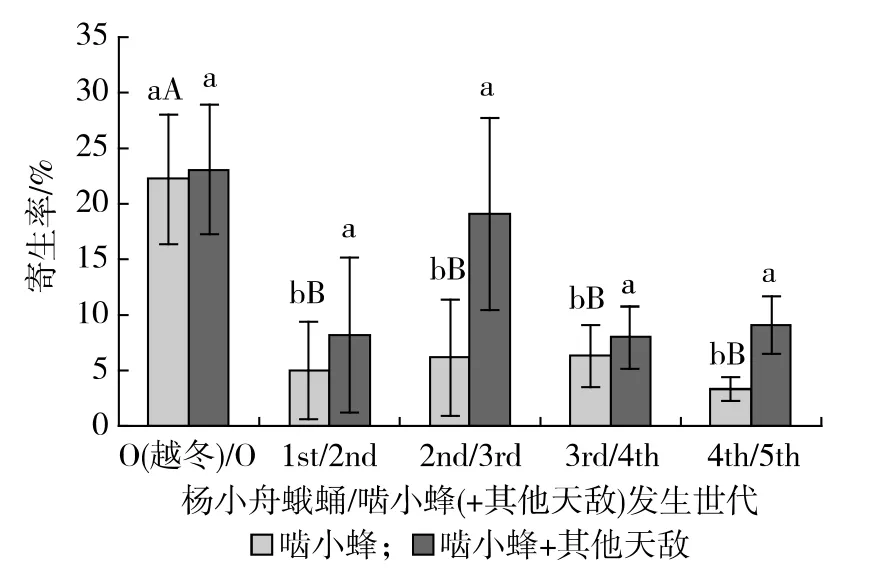
图1 杨小舟蛾各代蛹的林间自然寄生率
2.2人工释放后种群数量变化
2.2.1杨小舟蛾越冬蛹期释放2009年于云龙六堡和新沂草桥2地杨小舟蛾越冬蛹期释放后,白蛾黑基啮小蜂对越冬蛹和第1~4代蛹的寄生率和天敌总寄生率的观测结果见图2。由图2-a可知,在云龙六堡杨树成片林释放白蛾黑基啮小蜂后,杨小舟蛾越冬蛹的寄生率高达(40.3±5.6) %,极显著高于第1代和第3~4代蛹(P<0.01),显著高于第2代蛹(P<0.05) ;第2代蛹寄生率为(29.3± 11.7) %,也极显著高于第1代和第3~4代蛹(P<0.01) ;第1代和第3~4代蛹的寄生率无显著差异。天敌的总寄生率也以越冬蛹和第2代蛹最高(41.3%~46.6%),极显著高于其他各代。因此,杨树成片林内杨小舟蛾越冬蛹期释放后,白蛾黑基啮小蜂越冬代种群数量显著上升,对寄主越冬蛹具有显著的寄生效果;第1,2代种群数量显著下降,对第1代蛹的持续寄生作用显著下降;但第3代种群数量又显著上升,对第2代蛹具有显著的持续寄生效果;第4~5代种群数量较低,对3~4代蛹的持续寄生效果较差。

图2 杨小舟蛾越冬蛹期释放后各代蛹的寄生率
新沂草桥杨树林带释放后,杨小舟蛾越冬蛹的寄生率高达(33.7±6.6) %,也极显著高于第2~4代蛹(P<0.01) ;第2,3代蛹寄生率(3.2%~5.6%)无显著差异;第4代蛹的寄生率为(13.5± 1.8) %,显著高于第2~3代(P<0.05)。杨小舟蛾越冬蛹天敌的总寄生率(34.5%)也极显著高于其他各代(P<0.01),第4代蛹总寄生率(18.2%)显著高于第2~3代(P<0.05) (见图2-b)。因此,杨树林带内杨小舟蛾越冬蛹期释放后,白蛾黑基啮小蜂越冬代种群数量也显著上升,但第3~4代种群数量显著下降(第1代寄主蛹数量较少,未调查),而第5代种群数量明显上升,对第4代寄主蛹具有一定的持续寄生效果。
2.2.2杨小舟蛾第2代蛹期释放2010年杨小舟蛾第2代蛹期释放后,白蛾黑基啮小蜂对释放当代(第2代)寄主蛹的寄生率高为(44.4±5.4) %,极显著高于第3~4代(P<0.01),天敌总寄生率为(50.8±3.8) %,也极显著高于第3~4代(P<
0.01) (见图3) ;第2代寄生蛹的寄生率和天敌总寄生率均极显著高于同期CK寄生率,表明白蛾黑基啮小蜂释放后对当代寄主蛹具有显著的寄生效果; 第3代蛹的寄生率和总寄生率与同期CK均无显著差异,表明放蜂后对第3代蛹的持续寄生效果不佳;而第4代蛹的持续寄生率和总寄生率分别为(13.5 ±8.2) %和(17.7±11.6) %,其中持续寄生率与同期CK间存在显著差异(P<0.05),表明放蜂后对第4代蛹的持续寄生作用显著(见图3)。因此,于杨小舟蛾第2代蛹期释放,释放当代(第3代)林间小蜂的种群数量显著上升,但释放后下1代(第4 代)种群数量显著下降,释放后下2代(第5代)虽较第4代种群数量显著上升,但仍显著低于第3代小蜂的种群数量。
2.3室内成蜂羽化与林间寄主化蛹进度
2010年白蛾黑基啮小蜂在室温下的累计羽化进度和杨小舟蛾林间累计化蛹进度观测结果见图4。

图3 杨小舟蛾第2代蛹期释放后对第2~4代蛹的寄生率
按照张孝羲[7]对昆虫发育进度预测中始盛期、高峰期和盛末期的划分标准(累计发育进度分别为16%,50%和84%),由图4计算出的白蛾黑基啮小蜂羽化和杨小舟蛾化蛹的始盛期、高峰期、盛末期及2~4代小蜂羽化与其相应世代寄主化蛹的期距见表1。
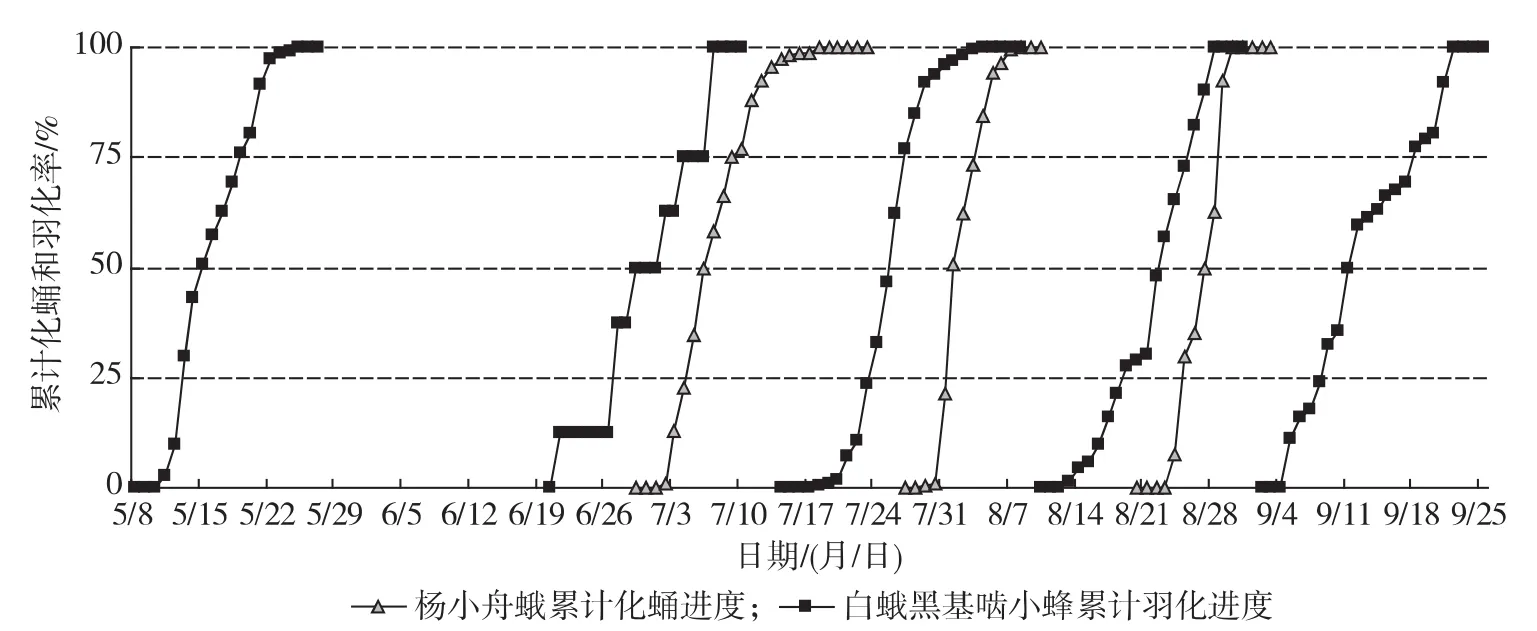
图4 白蛾黑基啮小蜂累计羽化进度和杨小舟蛾累计化蛹进度

表1 白蛾黑基啮小蜂羽化和杨小舟蛾化蛹高峰期测定结果
由表1可知,白蛾黑基啮小蜂越冬代成蜂羽化盛期为5月中旬;第2~5代成蜂羽化盛期分别为6月下旬、7月上旬、7月下旬、8月中、下旬和9月上、中旬。受2010年早春持续低温影响[8],越冬代和第1代成蜂羽化盛期推迟,第1代寄主蛹的发生也受气候影响而相应推迟,但随着气温回升,第2~5代成蜂羽化盛期与对应世代寄主蛹的发生期基本吻合(见图4)。由图4还可以看出,第2~4代小蜂羽化
期均早于相应寄主世代(第2~4代杨小舟蛾)蛹的发生期,发生的期距分别为(6.0±1.0),(7.7± 1.2),(5.3±2.5) d,3个期距之间均无显著差异,但以第3代期距最长。
3 讨论
郭同斌等研究认为,白蛾黑基啮小蜂属追踪寄生杨小舟蛾各代蛹的专性寄生蜂,越冬代成蜂羽化后是靠寄生尚未羽化的杨小舟蛾越冬蛹来完成第1代小蜂的世代发育,而此时越冬代小蜂与杨小舟蛾越冬代成虫几乎同时羽化,而且杨树人工纯林内存在其他转主寄主的可能性较小,可供越冬代小蜂寄生的寄主越冬蛹的数量极低[5],加之杨小舟蛾第1代是种群数量积累阶段,可供2代小蜂寄生的1代寄主蛹的数量也较低[9],由此可以推断第1,2代啮小蜂自然种群数量均明显下降,对第1代寄主蛹的寄生率也明显降低。本研究证实该寄生蜂在杨树人工纯林内对杨小舟蛾越冬蛹的平均自然寄生率高达22.2%,极显著高于第1~4代蛹,表明该蜂第1~5代自然种群数量极显著低于越冬代,从而证实了上述推断。各代小蜂室温下的羽化进度与相应世代寄主蛹发育进度相吻合,前者较后者提前5.3~7.7 d,其中第3代期距最长,这可能与第3代寄主蛹和小蜂的发育正处于夏季高温期,寄主蛹发育历期明显缩短,寄生蜂也需相应提前羽化而寻找寄主蛹寄生的生物学特性有关。上述研究结果表明,小蜂相应寄主世代提前羽化,为小蜂羽化后搜寻寄主蛹并产卵寄生提供了有效时间,与郭同斌等的报道相一致,进一步证实了该蜂属追踪寄生各代寄主蛹的“专性寄生蜂”的研究结论[5]。
研究证实,于越冬代和第2代害虫蛹期人工释放白蛾黑基啮小蜂,可显著增加释放当代(越冬代和第2代)害虫蛹的寄生率,并对释放后各代害虫蛹具有一定的持续寄生效果,但不同世代持续寄生效果不一样:越冬代释放后云龙和新沂2地分别以3代小蜂对2代蛹、5代小蜂对4代蛹的持续寄生效果较好,第2代蛹期释放后5代小蜂对4代蛹持续寄生作用明显,其余世代的持续寄生效果均不佳。上述试验结果差异可能与试验林分结构差异和第2~4代寄主种群数量较高等有关。杨小舟蛾生物学研究结果[9]表明,第1代是害虫种群数量积累阶段,第2代为种群数量增殖阶段,第3~4代为种群数量急剧上升的成灾阶段,因此第2~4代杨小舟蛾林间种群数量较越冬代和第1代显著上升。第2~4代蛹数量显著上升可能为造成该寄生蜂第1~5代自然寄生率和包括其他天敌在内的总寄生率显著下降的主要原因,这有待进一步研究。
根据上述自然与人工放蜂后白蛾黑基啮小蜂种群数量变化规律,可选择越冬代和第2代寄主化蛹盛期释放该蜂生物防治杨小舟蛾。一方面,寄主害虫种群处于基数和增长的初期,为发挥天敌防治作用的关键期;另一方面,上述2个世代释放后可显著降低越冬代和第2代寄主种群数量,并对放蜂后各代具有一定的持续寄生作用,可充分发挥寄生蜂控制效果。在防治生产上,本研究的防治试验区经连续2 a持续释放,杨小舟蛾基本保持了有虫无灾的低虫口状态,从而初步证实杨小舟蛾越冬代和第2代为释放该蜂防治杨树害虫的有利防治时期。
参考文献:
[1]郭同斌,颜学武.黑棒啮小蜂种团(膜翅目姬小蜂科)寄生蜂研究进展[J].南京林业大学学报:自然科学版,2011,35(6) : 127-133.
[2]颜学武,郭同斌,蒋继宏,等.白蛾黑基啮小蜂的生物学特性[J].南京林业大学学报:自然科学版,2008,32(6) :29-33.
[3]颜学武,郭同斌,蒋继宏,等.白蛾黑基啮小蜂寄主接受行为的研究[J].南京林业大学学报:自然科学版,2009,33 (5) : 121-125.
[4]杜伟,郭同斌,颜学武,等.白蛾黑基啮小蜂的发育起点温度和有效积温[J].中国生物防治,2009,25(4) :368-369.
[5]郭同斌,王虎诚,徐克勤,等.白蛾黑基啮小蜂的人工繁殖及其对杨小舟蛾的防治效果[J].南京农业大学学报:自然科学版,2010,33(5) : 81-86.
[6]高惠璇,李东风,耿直,等.SAS系统与基础统计分析[M].北京:北京大学出版社,1995:117-138.
[7]张孝羲.昆虫生态及预测预报[M].北京:农业出版社,1985: 303-317.
[8]杨中林,熊大斌,郭同斌,等.杨小舟蛾成虫和蛹自然发育进度研究[J].江苏林业科技,2011,38(3) : 1-5.
[9]郭同斌,王振营,梁波,等.杨小舟蛾的生物学特性[J].南京林业大学学报,2000,24(5) :56-60.
Preliminary study on the change rhythm of population quantity of Tetrastichus nigricoxae parasitizing Micromelalopha troglodyta pupae
WU Jing1,GUO Tong-bin1*,SONG Ming-hui1,DU Wei1,WANG Hu-cheng1,GUO Jia-zhong2
(1.Forest Pest Management and Quarantine Station of Xuzhou,Xuzhou 221009,China;
2.Gaoyou Administration Office of Grand Canal,Gaoyou 225600,China)
Abstract:Experiments were made to determine population quantity of Tetrastichus nigricoxae Yang under nature and artificial releasing in the non-releasing and releasing areas of poplar plantation damaged by Micromelalopha troglodyta within Xuzhou Forestry Centre,Yunlong District and Xinyi City.The results showed that the population quantity of overwinter parasitoids was highest.The parasitism rate of overwinter M.troglodyta pupae was (22.2±5.8) %,significantly higher than those of first to fourth pupae (3.4%~6.4%).The population quantity of T.nigricoxae and parasitism rate of overwinter and second pupae were increased after releasing in overwinter and second generations of M.troglodyta.The continuous control effects of the parasitoids were found to next pest’s generations.The accumulative eclosion progress of all generations of adult T.nigricoxae corresponded with M.troglodyta pupation.
Key words:Tetrastichus nigricoxae Yang; Micromelalopha troglodyta Graeser; Population quantity; Change rhythm
通信作者:*郭同斌(1967-),男,博士,研究员级高级工程师。E-mail: gtb1101@163.com。
作者简介:吴静(1969-),女,工程师,从事林业有害生物防治技术推广工作。
基金项目:江苏省“333工程”科研项目“人工繁殖与释放白蛾黑基啮小蜂等杨树害虫生物防治新技术”(BRA2010027)
收稿日期:2015-03-15;修回日期:2015-03-19
文章编号:1001-7380(2015) 02-0001-05
中图分类号:S763; Q968
文献标识码:Adoi:10.3969/j.issn.1001-7380.2015.02.001
猜你喜欢
现代农业科技(2016年23期)2017-04-06
西部资源(2017年1期)2017-03-27
农民致富之友(2016年22期)2017-02-07
中国实用医药(2016年28期)2016-12-07
科教导刊·电子版(2016年22期)2016-11-02
中国医药科学(2016年7期)2016-10-11
中国民族民间医药·下半月(2014年11期)2014-12-10
农业科技与装备(2014年6期)2014-11-28
湖北农业科学(2014年6期)2014-07-02
