
自主跑轮运动对小鼠空间学习记忆及Notch信号通路的影响
2015-01-28 19:20张宪亮徐波陆乐何标余锋季浏
体育学刊 2014年6期
张宪亮+徐波+陆乐+何标+余锋+季浏
摘 要:为了解自主跑轮运动对C57BL/6小鼠空间学习记忆能力及海马内Notch信号通路的影响,选用12只雄性5周龄C57BL/6小鼠,随机分为运动组(R组,n=6只)和对照组(C组,n=6只),运动组进行8周的自主跑轮运动,对照组安静饲养,第8周开始利用Morris水迷宫测试其学习记忆能力。运动结束后,断颈处死小鼠取双侧海马,用Western Blot检测小鼠大脑海马内NICD的蛋白表达水平,用RT-PCR检测小鼠大脑海马内的Jagged-1、Notch-1、PS-1、Hes-1的mRNA表达水平。结果显示:Morris水迷宫实验显示,运动组小鼠学习记忆能力明显强于对照组;Western Blot测试,运动组小鼠海马内NICD表达显著上调,RT-PCR测试,运动组小鼠海马内Jagged-1 mRNA、Notch-1 mRNA、PS-1 mRNA和Hes-1 mRNA表达水平较对照组显著上调。结果说明,8周自主跑轮运动激活Notch信号通路,提高了海马空间学习记忆能力。
关 键 词:运动生物化学;学习记忆;Notch信号通路;自主跑轮运动;小鼠
中图分类号:G804.7 文献标志码:A 文章编号:1006-7116(2014)06-0134-05
Effects of voluntary wheel exercise on spatial learning and memory and
Notch signal pathway of mice
ZHANG Xian-liang1,XU Bo1,LU Le2,HE Biao1,YU Feng1,JI Liu1
(1.Key Laboratory of Adolescent Health Assessment and Exercise Intervention Ministry of Education,
School of Physical Education and Health,East China Normal University,Shanghai 200241,China;
2.School of Physical Education,Shanghai Normal University,Shanghai 200234,China)
Abstract: In order to understand the effects of voluntary wheel exercise on spatial learning and memory and Notch signal pathway in hippocampus of C57BL/6 mice, the authors prepared 12 5-week old male C57BL/6 mice, randomly divided them into an exercise group (group R, n=6) and a control group (group C, n=6), let the mice in group R do a voluntary wheel exercise for 8 weeks, let the mice in group C live in a calm condition, starting from week 8, tested their learning and memory ability by utilizing Morris water maze, after the exercise was finished, cut the neck of the mice and took out hippocampus at both sides, measured the protein expression level of NICD in hippocampus of the mice by means of Western blot, measured the mRNA expression levels of Jagged-1, Notch-1, PS-1 and Hes-1 in hippocampus in the brains of the mice by means of RT-PCR, and revealed the following findings: the Morris water maze experiment showed that the learning and memory ability of the mice in group R was significantly stronger than that of the mice in group C; in Western blot test, the expression of NICD in hippocampus of the mice in group R increased significantly; in RT-PCR test, the mRNA expression levels of Jagged-1, Notch-1, PS-1 and Hes-1 in hippocampus of the mice in group R increased significantly as compared with those of the mice in group C. The said findings indicated that 8-week voluntary wheel exercise activated Notch signal pathway, enhanced the spatial learning and memory ability of hippocampus.endprint
Key words: sports biochemistry;learning and memory;Notch signal pathway;voluntary wheel exercise;mouse
运动作为中枢神经系统一种有效的刺激形式,在大脑结构和功能可塑性的调节中起重要作用,大量研究发现适宜的运动可以提高大脑学习记忆能力,但其机制是多方面的,也是不确切的。近来有研究发现,Notch信号通路在学习记忆能力的调节中起了关键作用[1]。Notch信号通路是一条传递细胞与细胞间信号的信号转导途径,在进化上高度保守,它精确调控着神经系统的发育分化及凋亡,并与突触可塑性、海马学习记忆能力等脑的高级功能密切相关。Ge等[2]发现敲除果蝇内的Notch基因导致其长期记忆能力下降,而过表达Notch基因野生型果蝇内可增强其长期记忆能力,提示Notch信号通路在果蝇的长期记忆能力中起了关键作用。Costa等[3]选用Notch+/-转基因鼠和野生型鼠进行实验,发现Notch+/-转基因鼠空间学习能力下降,而其他学习能力没有改变,同时选用RBP-J+/-转基因鼠和野生型鼠进行实验,发现RBP-J基因缺失同样造成了空间学习记忆能力的下降,说明,Notch信号通路在海马空间学习记忆能力的调节中起重要作用。而且Zhang等[4]发现Notch信号通路可能通过促进海马神经发生,提高突触可塑性,从而改善海马学习记忆能力。但是,运动对小鼠海马空间记忆能力的提高是否通过激活Notch信号通路呢?本实验通过8周的自主跑轮运动干预,研究运动对5周龄C57BL/6雄性小鼠海马Notch信号通路中Jagged-1、Notch-1、PS-1、Hes-1表达的影响,并探讨其在运动改善学习记忆能力中的作用。
1 研究对象与方法
1.1 实验动物及分组
5周龄健康雄性C57BL/6小鼠12只(由上海斯莱克公司提供),每笼1只,常规分笼饲养,随机分为跑轮运动组(R组)和对照组(C组),每组6只。环境室温:(22±5) ℃;动物饲养环境湿度:40%~60%;光照:自然昼夜交替。自由饮食、饮水。
1.2 运动方案
对照组放于不含跑轮的饲养笼内,安静饲养;跑轮运动组放于含有跑轮的饲养笼内,进行为期8周的跑轮运动,运动量的记录每天1次。第8周开始,两组小鼠均进行Morris水迷宫测试,上午10:00开始,为期 6 d,1~5 d为定位航行实验,第6天为空间探索实验。
1.3 取材
8周训练结束后,将小鼠断颈处死,取双侧海马组织,放入-80 ℃冰箱保存,待测PCR及Western blot指标。
1.4 Western blot检测海马内NICD蛋白表达水平
按每20 mg组织加入100~200 μL的Western裂解液(含PMSF)中混匀,对称置于瓷珠匀浆仪中匀浆后,置于冰上静置10 min,重复2~4次。于4 ℃下,12 000 g离心3~5 min,取上清,分装,置于-80 ℃冰箱待测。根据BCA蛋白浓度测定试剂盒步骤,测定蛋白含量。计算蛋白上样量,使用SDS-PAGE电泳将蛋白转至PVDF膜上。经2 h封闭后,分别加入一抗(NICD,1︰500,c-23307,santa),4 ℃下孵育过夜。次日加入二抗(HRP,IgG,1︰1 000,碧云天),室温孵育2 h。TBST摇床洗10 min,重复3次后,于暗室使用ECL发光显影,Alpha凝胶成像系统自动曝光,捕捉图像,ImageJ1.46对图像进行灰度值分析,将NICD蛋白与内参蛋白的平均密度之比作为NICD的相对表达水平。
1.5 RT-PCR检测BDNF、Jagged-1、Notch-1、PS-1、Hes-1的mRNA
取适量的海马组织,用Trizol法提取小鼠海马内总的RNA,逆转录得到稳定的cDNA,实时荧光定量PCR检测Jagged-1、Notch-1、PS-1、Hes-1mRNA的相对表达量,引物序列见表1。
1.6 统计分析
运用SPSS17.0进行统计学分析计算,数据处理使用T检验,P<0.05具有显著性差异,P<0.01具有极显著性差异。所有数据均用均数±标准差来表示。
2 实验结果及分析
运动组小鼠平均每天运动量为(7 418.47±
2 432.15) m。
2.1 对小鼠Morris水迷宫测试的影响
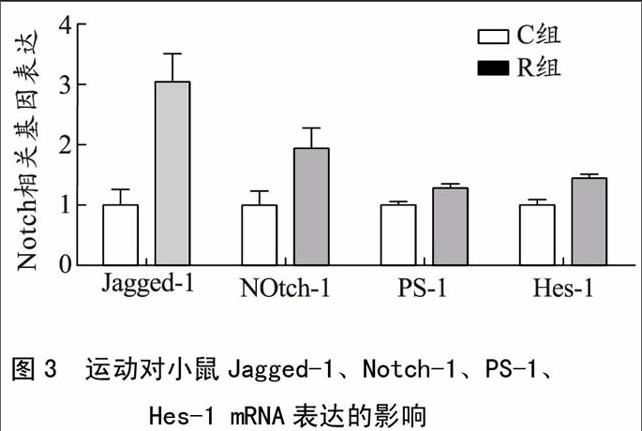
1)定位航行实验结果见图1。
图1 自主跑轮运动对小鼠Morris水迷宫实验
定位航行实验的影响
如图1所示:5 d定位航行实验中,两组小鼠的潜伏期呈下降趋势,运动组小鼠的潜伏期均低于对照组小鼠。其中,第2天和第3天运动组小鼠潜伏期显著低于对照组小鼠(P<0.05)。而两组小鼠的平台象限路程百分比呈上升趋势;而且运动组小鼠平台象限路程百分比高于对照组,其中两组小鼠第1天的平台象限路程百分比出现了显著性差异(P<0.05)。
2)空间探索实验结果。
在空间探索实验中,与对照组相比((22.81±5.28)%),运动组小鼠平台象限路程百分比((30.78±7.78)%)出现了显著性差异(P<0.05)。
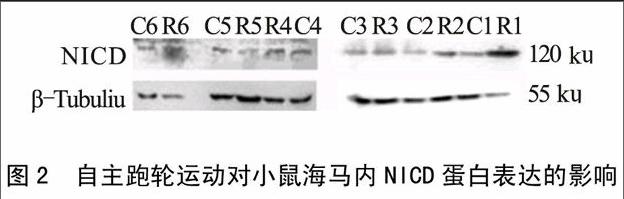
2.2 对小鼠海马内NICD蛋白表达的影响
如图2所示,与对照组(0.21±0.11)相比,运动组小鼠海马内NICD蛋白表达(0.75±0.36)显著上调(P<0.05)。
图2 自主跑轮运动对小鼠海马内NICD蛋白表达的影响
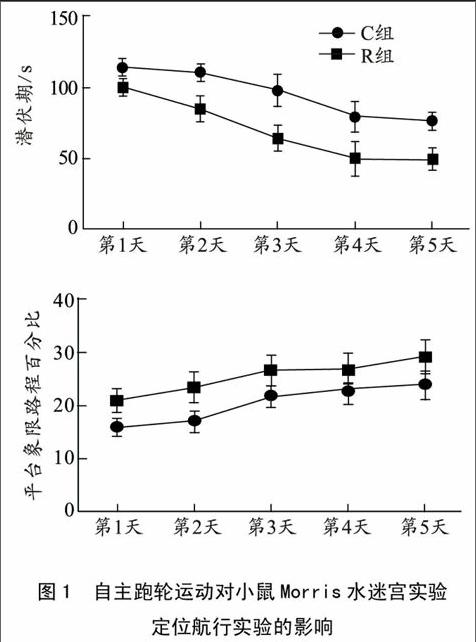
2.3 对小鼠海马内Notch信号通路相关基因mRNA的影响
如图3所示,与对照组相比,运动组Jagged-1 mRNA(P<0.01)、Notch-1 mRNA(P<0.05)、PS-1 mRNA(P<0.05)和Hes-1 mRNA(P<0.01)的表达水平均出现明显上调,其中,Jagged-1和Hes-1出现了非常显著性。endprint
图3 运动对小鼠Jagged-1、Notch-1、PS-1、
Hes-1 mRNA表达的影响
3 讨论
Morris水迷宫是一种强迫实验动物(一般指大鼠、小鼠)通过被动游泳的方式,学习寻找并记忆隐藏于水下平台的实验,用于大鼠或小鼠学习、记忆能力的检测。包括定位航行实验和空间探索实验。经实验发现,两组小鼠潜伏期均呈现逐渐下降的趋势、平台象限路程百分比均呈上升趋势,说明两组小鼠均经历一个学习的过程,逐渐记忆平台象限的位置。并且在第2、3天运动组小鼠潜伏期显著低于对照组小鼠,第1天运动组小鼠平台象限路程百分比显著高于对照组小鼠,说明运动组小鼠在定位航行实验中的表现要好于对照组小鼠。在空间探索实验中,运动组小鼠平台象限路程百分比显著高于对照小鼠,说明运动组小鼠记忆平台象限的表现要好于对照组小鼠,运动组小鼠呈现出较好的记忆能力。运动组小鼠在迷宫实验中更有目的性,学习记忆能力要好于对照组。提示,8周的自主跑轮运动提高了小鼠学习记忆能力。Uysal等[5]将22 d Wistar大鼠分为对照组和运动组,运动组进行8周的跑台训练,并对其进行Morris水迷宫测试,结果显示运动组小鼠提高了空间记忆能力。褚昕宇等[6]将10周龄C57BL/6小鼠分为对照组、应激组、运动组和应激运动组。通过Morris水迷宫实验检测其学习记忆能力,发现运动不仅可以提高正常小鼠学习记忆能力,还可以抑制应激造成的小鼠学习记忆能力的损害。这些研究与本实验的研究结果表现出高度的一致性,说明适宜的体育运动可以提高海马空间学习记忆能力。其原因可能是:长期适宜运动后,血液循环加快,大脑海马得到充足的养料及氧气,促使脑细胞发育良好;而且适宜的体育运动促使机体体温升高,增强了大脑海马内相关抗氧化酶的活性[7]。
Notch信号转导途径参与了学习记忆的神经过程。研究发现在敲除Notch基因的果蝇内,其长时程记忆受到损害[8]。对Notch基因突变的小鼠进行水迷宫实验,发现其空间记忆能力缺陷,并发现其突触可塑性受到抑制,而给予Notch配体Jagged-1后,学习记忆能力得以改善,说明Notch通路与海马学习记忆密切相关[9-10]。Notch信号通路主要由Notch受体、Notch配体、CSL转录因子以及靶分子等构成。Notch受体在神经干细胞上高度表达,某些诱导因子的作用下,干细胞上的Notch受体与邻近细胞上的Notch配体相互结合,经过肿瘤坏死因子α转化酶(tumor necrosis factor-α converting enzyme TACE)和γ-分泌酶(主要成分为PS-1)的酶切,产生NICD(notch intracellular domain,NICD)蛋白,并将有活性的NICD释放入胞浆转移到核内,与CSL蛋白结合,形成转录因子,激活Hes-1等靶基因,参与神经系统的发生,提高海马学习记忆能力[11]。
本实验发现,自主运动组小鼠NICD蛋白显著上调,Jagged-1、Notch-1、PS-1以及Hes-1mRNA表达显著增加。NICD是Notch信号通路激活的标记分子,Hes-1是NICD转移至核内后激活的靶基因,因此NICD蛋白的高表达和Hes-1基因的高表达说明运动后Notch信号通路被激活。Moritz等[12]通过免疫组织化学方法发现运动后增加了神经细胞的数量,同时发现在新增殖的细胞内同时伴随有NICD的阳性表达,说明运动激活了Notch信号通路,与本实验结果一致。另外,本实验同时发现运动组小鼠学习记忆能力显著高于对照组,说明运动提高学习记忆能力的同时激活了Notch 信号通路,提示Notch信号通路可能介导了运动诱导的学习记忆能力提高。
NICD是Jagged-1与Notch-1结合后在TACE酶和γ-分泌酶酶切下形成的,因此Jagged-1与Notch-1的表达水平与NICD密切相关。Tsivitse等[13]在骨骼肌重塑实验中发现,下坡跑上调了Notch-1与其配体的表达,激活Notch信号通路,介导了骨骼肌重塑。本实验中运动组小鼠Jagged-1和Notch-1基因表达水平显著高于对照组小鼠,说明运动组小鼠Notch信号通路中Jagged-1与Notch-1的结合更多,提示运动组小鼠NICD的表达上调可能与运动后Jagged-1和Notch-1的高表达有关,与Tsivitse等人研究一致。另有研究发现在敲除TrkB基因的小鼠内Jagged-1表达显著低于对照组小鼠,说明BDNF可以通过与其受体TrkB结合,增加Jagged-1的表达,从而激活Notch信号通路[14]。同时大量实验证实自主跑轮运动后海马神经元内的BDNF mRNA和TrkB mRNA的表达明显增加[15-16]。因此推测运动可能通过增加BDNF表达水平,提高Jagged-1表达,从而激活Notch信号通路。
于此同时,Notch受体与配体结合后,必须在S2、S3位点分别经过TACE与γ-分泌酶的酶切,才能形成NICD,从而调节Notch信号通路。PS-1是γ-分泌酶的主要成分,是Notch信号通路中的关键调节酶。Chen等[17]选用野生型鼠作为研究对象,测试其LTP,发现抑制γ-分泌酶的活性可导致其LTP下降,因此,γ-分泌酶在正常生理条件下可能起了一个积极的作用。本实验同时检测了PS-1的mRNA水平,发现运动组小鼠PS-1mRNA增加,说明运动激活Notch信号通路也可能与PS-1的表达上调有关。但是,关于γ-分泌酶研究主要集中于阿尔茨海默病(AD),它参与了AD的发病特征老年斑的核心物质Aβ的形成。研究发现抑制γ-分泌酶的活性,可以减少Aβ的形成,从而改善AD的学习记忆能力[18]。王芳[19]研究发现有氧跑台训练可以显著降低-D半乳糖注射造模AD大鼠海马内PS-1的基因表达,与本实验的研究并不一致。原因可能是所选老鼠模型不同,PS-1在野生型鼠中参与了Notch信号通路的激活,促进了海马神经发生,提高了突触可塑性,促进海马空间学习记忆能力;而PS-1在AD鼠内,主要介导了Aβ的形成,引起机体氧化应激、炎症反应、细胞凋亡等,降低海马空间学习记忆能力。因此关于运动对PS-1的影响仍不十分清楚,需进一步研究证实。endprint
参考文献:
[1] Ables J L,Breunig J J,Eisch A J,et al. Notch just development:notch signalling in the adult brain[J]. Nat Rev Neurosci,2011,12(5):269-283.
[2] Ge X,Hannan F,Xie Z,et al. Notch signaling in drosophila long-term memory formation[J]. Proc Natl Acad Sci U S A,2004,101(27):10172-10176.
[3] Costa R M,Honjo T,Silva A J. Learning and memory deficits in notch mutant mice[J]. Curr Biol,2003,13(15):1348-1354.
[4] Zhang X,Huang G,Liu H,et al. Folic acid enhances notch signaling,hippocampal neurogenesis,and cognitive function in a rat model of cerebral ischemia[J]. Nutr Neurosci,2012,15(2):55-61.
[5] Uysal N,Tugyan K,Kayatekin B M,et al. The effects of regular aerobic exercise in adolescent period on hippocampal neuron density,apoptosis and spatial memory [J]. Neurosci Lett,2005,383(3):241-245.
[6] 褚昕宇,季浏,王泽军. 跑转轮运动与限制性应激对成年小鼠海马神经发生以及空间学习记忆能力的影响[J]. 北京体育大学学报,2013,36(5):69-74.
[7] 徐波,黄涛,季浏. 跑台训练增强大鼠的学习记忆及其海马BDNF基因表达[J]. 北京体育大学学报,2011,34(4):51-54.
[8] Lasky J L,Wu H. Notch signaling,brain development,and human disease[J]. Pediatr Res,2005,57(5 Pt 2):104R-109R.
[9] Dahlhaus M,Hermans J M,Van Woerden L H,et al. Notch1 signaling in pyramidal neurons regulates synaptic connectivity and experience-dependent modifications of acuity in the visual cortex[J]. J Neurosci,2008,28(43):10794-10802.
[10] F?rster E,Bock H H,Herz J,et al. Emerging topics in reelin function[J]. Eur J Neurosci,2010,31(9):1511-1518.
[11] Ehm O,G?ritz C,Covic M,et al. RBPJkappa-dependent signaling is essential for long-term maintenance of neural stem cells in the adult hippocampus[J]. J Neurosci,2010,30(41):13794-13807.
[12] Moritz D Brandt,Antonia Maass,Gerd Kempermann,et al. Physical exercise increases Notch activity,proliferation and cell cycle exit of type-3 progenitor cells in adult hippocampal neurogenesis[J]. European Journal of Neuroscience,2010,32(8):1256-1264.
[13] Tsivitse S K,Peters M G,Stoy A L,et al. The effect of downhill running on Notch signaling in regenerating skeletal muscle[J]. Eur J Appl Physiol,2009,106(5):759-767.
[14] Dorfman M D,Kerr B,Garcia-Rudaz C,et al. Neurotrophins acting via TrkB receptors activate the jagged1-Notch2 cell-cell communication pathway to facilitate early ovarian development[J]. Endocrinology,2011,152(12):5005-5016.
[15] Vaynman S,Ying Z,Gomez pinilla F. Interplay between brain derived neurotrophic factor and signal transduction modulators in the regulation of the effects of exercise on Synaptic plasticity[J]. Neuroscienc,2003,122(3):647-657.
[16] Berchtold N C,Kesslak J P,Cotman C W,et al. Hippocampal brain derived neurotrophic factor gene regulation by exercise and the medial septum[J]. Neurosci Res,2002,68(5):511-521.
[17] Chen Y,Behnisch T. The role of γ-secretase in hippocampal synaptic transmission and activity-cdependent synaptic plasticity[J]. Neurosci Lett,2013,56(13):300151-300156.
[18] Townsend M,Qu Y,Gray A,et al. Oral treatment with a gamma-secretase inhibitor improves long-term potentiation in a mouse model of Alzheimers disease[J]. J Pharmacol Exp Ther,2010,333(1):110-119.
[19] 王芳. 有氧运动对AD大鼠学习记忆、Aβ相关基因表达及脂质过氧化的影响[D]. 上海:华东师范大学,2011:25.endprint
猜你喜欢
作文周刊·小学二年级版(2022年20期)2022-05-05
创新作文(小学版)(2019年10期)2019-09-25
作文周刊·小学一年级版(2016年27期)2017-06-03
小学生学习指导(低年级)(2017年5期)2017-05-04
海外文摘(2016年4期)2016-04-15
中国病理生理杂志(2015年8期)2015-12-21
医学研究杂志(2015年3期)2015-06-10
创业家(2015年1期)2015-02-27
遗传(2014年2期)2014-02-28
