
Acupuncture inhibits Notch1 and Hes1 protein expression in the basal ganglia of rats with cerebral hemorrhage
2015-01-18 05:49WeiZouQiuxinChenXiaoweiSunQingbinChiHongyuKuangXuepingYuXiaohongDai
中国神经再生研究(英文版) 2015年3期
Wei Zou, Qiu-xin Chen, Xiao-wei Sun Qing-bin Chi Hong-yu Kuang, Xue-ping Yu Xiao-hong Dai
1 Third Department of Acupuncture, First Affliated Hospital, Heilongjiang University of Chinese Medicine, Harbin, Heilongjiang Province, China
2 Heilongjiang University of Chinese Medicine, Harbin, Heilongjiang Province, China
3 Department of Endocrinology, First Affliated Hospital, Harbin Medical University, Harbin, Heilongjiang Province, China
Acupuncture inhibits Notch1 and Hes1 protein expression in the basal ganglia of rats with cerebral hemorrhage
Wei Zou1,*, Qiu-xin Chen2, Xiao-wei Sun1, Qing-bin Chi1, Hong-yu Kuang3, Xue-ping Yu1, Xiao-hong Dai1
1 Third Department of Acupuncture, First Affliated Hospital, Heilongjiang University of Chinese Medicine, Harbin, Heilongjiang Province, China
2 Heilongjiang University of Chinese Medicine, Harbin, Heilongjiang Province, China
3 Department of Endocrinology, First Affliated Hospital, Harbin Medical University, Harbin, Heilongjiang Province, China
Notch pathway activation maintains neural stem cells in a proliferating state and increases nerve repair capacity. To date, studies have rarely focused on changes or damage to signal transduction pathways during cerebral hemorrhage. Here, we examined the effect of acupuncture in a rat model of cerebral hemorrhage. We examined four groups: in the control group, rats received no treatment. In the model group, cerebral hemorrhage models were established by infusing non-heparinized blood into the brain. In the acupuncture group, modeled rats hadBaihui(DU20) andQubin(GB7) acupoints treated once a day for 30 minutes. In the DAPT group, modeled rats had 0.15 µg/mL DAPT solution (10 mL) infused into the brain. Immunohistochemistry and western blot results showed that acupuncture effectively inhibits Notch1 and Hes1 protein expression in rat basal ganglia. These inhibitory effects were identical to DAPT, a Notch signaling pathway inhibitor. Our results suggest that acupuncture has a neuroprotective effect on cerebral hemorrhage by inhibiting Notch-Hes signaling pathway transduction in rat basal ganglia after cerebral hemorrhage.
nerve regeneration; acupuncture; cerebral hemorrhage; immunohistochemistry; western blot assay; Notch1; Hes1; rats; DAPT; neural stem cells; NSFC grant; neural regeneration
Funding:This study was supported by the National Natural Science Foundation of China, No. 81273824, 30772840; Ministry of Education Doctoral Fund in China, No. 20102327110003; the Natural Science Foundation of Heilongjiang Province in China, No. ZD201204; and Special funds for Technological Innovation Research of Harbin, China, No. 2012RFXXS062
Zou W, Chen QX, Sun XW, Chi QB, Kuang HY, Yu XP, Dai XH (2015) Acupuncture inhibits Notch1 and Hes1 protein expression in the basal ganglia of rats with cerebral hemorrhage. Neural Regen Res10(3)∶457-462.
Introduction
Cerebral hemorrhage is non-traumatic, and caused by rupture of blood vessels in brain parenchyma. The most common causes are hypertension, cerebral arteriosclerosis, and intracranial vascular malformations. Cerebral hemorrhage is often induced by exertion and emotion, and most patients show sudden onset during activity. A large number of studies have shown that cerebral hemorrhage causes neuronal apoptosis (Chamnanvanakij et al., 2002; Riggs et al., 2005; Gao et al., 2009), which aggravates the brain damage and causes contralateral limb dysfunction. Therefore, to reduce the degree of brain injury and promote functional recovery, it is important to inhibit neuronal apoptosis (Guan et al., 2013). Previous studies have shown that acupuncture reduces damage by effectively promoting growth, repair, and restoration of the nervous system on the injury side (Feng et al., 2013; Li et al., 2013; Nam et al., 2013), although the underlying mechanism is not fully understood.
The Notch signaling pathway is an evolutionarily conserved signal transduction pathway involved in regulating cell proliferation, differentiation, and apoptosis in almost all tissues and organs (Monahan et al., 2009; Guo et al., 2010; Yalcin-Ozuysal et al., 2010; Fernandez-Valdivia et al., 2011; Gianni-Barrera et al., 2011). Despite this, few studies have focused on the Notch signaling pathway in the central nervous system. Recently, it has been shown that related gene and protein expression in the Notch signal transduction pathway is present in the central nervous system (Yagi et al., 2012). Notch1, a Notch homologue, is a highly conserved cell membrane receptor protein, which can directly regulate gene transcription and inhibit or promote cell differentiation, proliferation, and apoptosis (Miele, 2006). Notch1 protein is widespread in embryonic neural tissue, but decreases rapidly after birth until it is no longer detected (Zhang et al., 2012). However, Notch1 protein expression is detected in adult neural stem cells from the subventricular zone of the lateral ventricles, the retina, and hippocampal dentate gyrus (Zhang et al., 2012).
The main role of the Notch signaling pathway is inproliferation and differentiation of stem cells, facilitation of inhibitory cell differentiation signalsvia“bypass inhibition”, and following signal transmission, initiation of family protein binding between cell membrane Notch receptors and its ligands, Delta/Serrate/Lag-2 (DSL) and CBF-1/Suppressor of hairless/Lag (CSL), on neighboring cells. During central nervous system development, Notch-Hes signaling inhibits neural stem cell differentiation into neuronal and glial cells by maintaining them in an undifferentiated state with self-renewing capacities. Binding of the Notch receptor to its ligand, activates the Notch signaling pathway and inhibits neuronal differentiation, improves the internal environment, promotes endogenous neural stem cell proliferation, and enhances nerve repair (Chapouton et al., 2010; Falk et al., 2012).In vitroexperiments show that during cerebral ischemic injury, the Notch signal transduction pathway upregulates endogenous Notch1 receptor expression (Guo et al., 2013).
To date, studies have seldom focused on changes in the Notch signaling pathway during cerebral hemorrhagic injury, and there are no consistent conclusions on the effects. Thus, in this study, we used a rat model of cerebral hemorrhage to investigate the effect of acupuncture on Notch1 and Hes1 expression in brain tissue. Our aim was to determine if acupuncture promotes restoration of brain injury after cerebral hemorrhage by increasing Notch1 and Hes1 expression, which may provide a theoretical foundation for acupuncture treatment during cerebral hemorrhage.
Materials and Methods
Animals
Healthy specific-pathogen-free male Wistar rats (n= 156), aged 50 days and weighing 350 ± 20 g were supplied by the Harbin Veterinary Research Institute in China (Certifcation No. SCXK (Hei) 20100027). All rats were housed at 22 ± 2°C with humidity of 50 ± 5% and noise < 60 dB.
Establishment of a cerebral hemorrhage model
In accordance with a previous method (Rosenberg et al., 1990), rats were intraperitoneally anesthetized with 10% chloral hydrate (350 mg/kg) (Royalton, Dalian, Liaoning Province, China), and fxed in the prone position to a stereotaxic apparatus. Rats were positioned with the anterior and posterior fontanelles in the same horizontal plane (determined by the upper incisor tooth hook plane being 2.4 mm lower than the plane between the ears). The median scalp was shaved and sterilized, and a median incision approximately 1 cm long was made. The periosteum was stripped using a bone stripper, and the anterior fontanelle and coronal suture exposed. A round hole (1.0 mm diameter) was drilled using a dental drill (3.5 mm right and 0.2 mm posterior to bregma) until the dural surface was reached. The tail was disinfected with alcohol and 3 cm cut from the caudal end. Blood (50 µL) was obtained using a microinjector fxed on the stereotaxic apparatus, which entered vertically approximately 6 mm along the hole. Non-heparinized blood (50 µL) was infused into the caudate putamen at 25 µL/min. The needle was maintained in place for 5 minutes, and simultaneously, gauze was used to bandage the tail wound. Finally, the skull wound was blocked with zinc phosphate cement and the scalp sutured.
Screening rat models
In accordance with Berderson’s score (Berderson et al., 1986), the specifc method for assessing cerebral hemorrhage models was: rats were lifted by their tails, until they were 10 cm higher than the desktop; the paw of a normal rat will be straight.

Berderson score:
After model establishment, the rats had regained consciousness, according to appropriate signs and symptoms, rats with intracranial hematoma were considered successful models of cerebral hemorrhage.
Experimental groups
The rats were randomly divided into control (n= 12), model (n= 48), acupuncture (n= 48), and N-[N-(3,5-difluorophenacetyl-L-alanyl)]-S-phenylglycine t-butyl ester (DAPT) (Notch inhibitor) (n= 48) groups. The model, acupuncture and DAPT groups were equally subdivided into four subgroups according to different time points (1, 3, 7, and 14 days) after model establishment. The control group was only observed at day 1. Rats in the control group did not receive any treatment, and were directly decapitated and samples collected. In the model group, samples were collected at 1, 3, 7, and 14 days after model establishment. In the acupuncture group, rats received acupuncture 12 hours after model establishment, and once every 24 hours thereafter. The acupuncture method was as follows: rats were fxed on the acupuncture board. TheBaihui(DU20) (specifc location: head between the middle ears) andQubin(GB7) (specific location: leading edge of ear root) acupoints were treated on the affected side using a 1.3 inch stainless steel needle (Huatuo, Suzhou, Jiangsu Province, China). The needling depth was 1.0 inch, and the needle was maintained in place for 30 minutes. During this time, the needle was twisted three times for 5 minutes each, at a speed of 200 cycles per minute. Hippocampal specimens were collected at 1, 3, 7, and 14 days after acupuncture. In the DAPT group, a round hole (3.6 mm diameter) was drilled using a dental drill (1.4 mm right and 0.8 mm posterior to bregma) 30 minutes before model establishment. Within 5 minutes, 10 µL DAPT solution (0.15 µg/µL) was injected using a microsyringe (Hamilton, Bonaduz, Switzerland)viathe lateral ventricle. The needle was maintained in place for 5 minutes. Hippocampal specimens were obtained at 1, 3, 7, and 14 days after modeling.
Specimen collection
Rats were intraperitoneally anesthetized using an overdose of chloral hydrate at appropriate time points after surgery.The chest was opened rapidly and the heart exposed. Catheterization from the left ventricle into the aortic root was performed. A small incision was made in the right atrial appendage for an exit hole. The ventricle was perfused using approximately 200 mL each of 4°C saline and 4% paraformaldehyde/0.1 M phosphate buffer. Samples were dehydrated, paraffn embedded, and then sliced into 6 µm-thick sections for hematoxylin-eosin staining and immunohistochemistry. Some rats were directly decapitated after anesthesia overdose, and basal ganglia tissue immediately stored in liquid nitrogen at -80°C for western blot assay.

Table 1 Notch1 protein expression (obtained from gray values) in the basal ganglia of rats with intracerebral hemorrhage
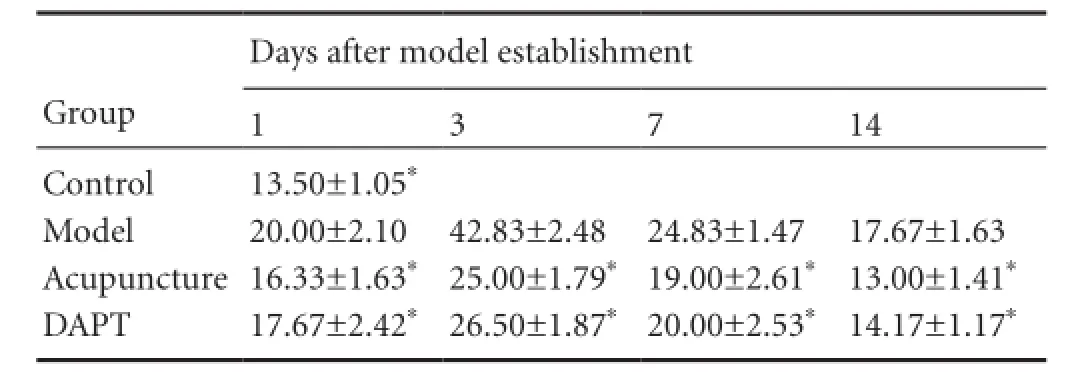
Table 3 Hes1 immunoreactivity (cell count, × 400 magnifcation) in the basal ganglia of rats with cerebral hemorrhage
Immunohistochemistry
Notch1 and Hes1 expression in the basal ganglia was determined by immunohistochemistry. After paraffn embedding and dehydration, sections were placed in 3% fresh H2O2solution at 25°C for 10 minutes to block endogenous peroxidase, and then washed in 0.01 M PBS three times for 2 minutes each. Sections were immersed in 0.01 M citrate buffer (pH 6) and heated twice in the microwave for 5 minutes each time, with 10 minute intervals. After naturally cooling to 25°C, sections were incubated with primary antibody (rabbit Notch1 or Hes1 polyclonal antibodies; 1:200; NeoMarkers, Fremont, CA, USA) overnight at 4°C, and washed with 0.01 M PBS three times for 2 minutes each. Sections were then incubated with biotinylated secondary antibody (horseradishperoxidase-goat anti-rabbit IgG; NeoMarkers) at 37°C for 30 minutes, washed with 0.01 M PBS three times for 2 minutes each, visualized with 3,3′-diaminobenzidine (Golden Bridge Biotechnology Co., Ltd., Beijing, China), counterstained with hematoxylin, mounted with neutral gum, and then observed under an optical microscope (Olympus, Tokyo, Japan).
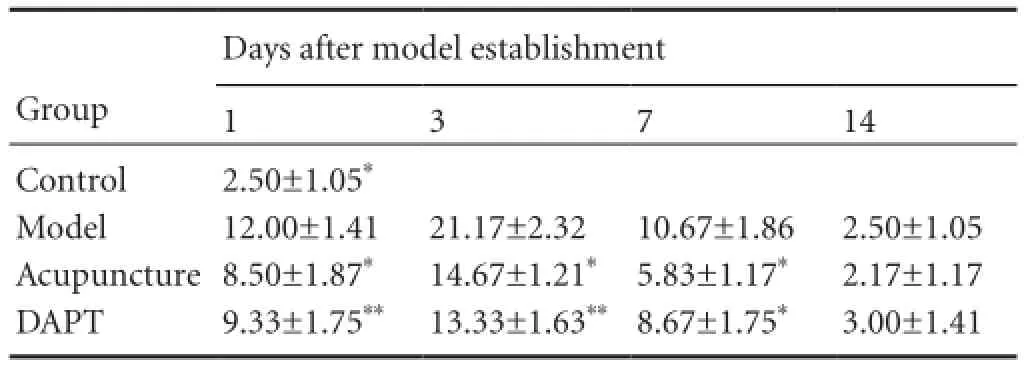
Table 2 Notch1 immunoreactivity (cell count, × 400 magnifcation) in the basal ganglia of rats with cerebral hemorrhage
Image analysis
Data were analyzed using Motic Med 6.0 pathological image analysis system (Motic Med Industrial Group Co., Ltd., Xiamen, Fujian Province, China). Each section was randomly observed and counted at 400× magnifcation. Total positive cell number was calculated.
western blot assay
Basal ganglia tissue was extracted using the RIPA lysis buffer protein BCA kit (Biomiga, San Diego, CA, USA) to determine total protein concentration. Total protein loaded from each sample was 50 µg/well. Samples were electrophoresed and transferred to membranes. Film was blocked with 5% skim milk in phosphate Tween buffer at 4°C overnight. Samples were incubated with primary antibody (rabbit Notch1, polyclonal antibody; 1:1,000; Epitomics, Burlingame, CA, USA) for 16—24 hours at 4°C. Membranes were then washed with Tris-buffered saline with Tween 20, and incubated in secondary antibody (goat anti-rabbit IgG; 1:1,500; Epitomics) for 1 hour at room temperature. Gray values for specifc bands were measured using Image J software (NIH, Bethesda, MD, USA).
Statistical analysis
Statistical analysis was performed by the frst author using SPSS 18.0 software (SPSS, Chicago, IL, USA), with values expressed as the mean ± SD. Inter-group data were compared by one-way analysis of variance and the least significant difference test. Values ofP< 0.05 were considered statistically signifcant.
Results
Notch1 protein expression in rat basal ganglia tissue

Figure 1 Notch1 protein expression in the basal ganglia of rats with intracerebral hemorrhage (western blot assay).

Figure 2 Notch1 immunoreactivity in the basal ganglia of rats with cerebral hemorrhage and acupuncture treatment (immunohistochemical staining, × 400 magnifcation).
In each group, Notch1 protein expression was visible in the basal ganglia at 1, 3, 7, and 14 days after modeling. Compared with the control group, Notch1 protein expression was signifcantly higher in the model group at 1, 3, and 7 days (P<0.01). Furthermore, compared with the model group, Notch1 protein expression was signifcantly lower in the DAPT group at each time point, particularly at 3 days (P< 0.01) although signifcant differences were still detected at 7 and 14 days (P< 0.05). At all time points, Notch1 protein expression was signifcantly lower in the acupuncture group than in the model group (P< 0.01), and was inhibited in the DAPT and acupuncture groups (P> 0.05; Table 1, Figure 1).
Notch1 immunoreactivity in rat basal ganglia tissue

Figure 3 Hes1 immunoreactivity in the basal ganglia of rats with cerebral hemorrhage (immunohistochemical staining, × 400).
Notch1-immunoreactive cells were widely distributed in the cytoplasm and nucleus. Weak Notch1 immunoreactivity was observed in the control group. Notch1-immunoreactive cell number was signifcantly higher in the model group than in the control group at various time points (P< 0.05), and signifcantly lower in the DAPT group than the model group at 1, 3 (P< 0.01), and 7 (P< 0.05) days. Notch1-immunoreactive cell number was lower in the acupuncture group than in the model group at 1, 3, and 7 days (P< 0.05). No signifcant difference was detected between the acupuncture and DAPT groups (P> 0.05; Table 2, Figure 2).
Hes1 immunoreactivity in rat basal ganglia tissue
Hes1-immunoreactive cells were mainly expressed in vascular endothelial and glial cells. Hes1 immunoreactivity was observed in the control group. At various time points, Hes1-immunoreactive cell number was signifcantly increased in the model group compared with the control group (P< 0.05). Moreover, Hes1-immunoreactive cell number was significantly lower in the acupuncture group than the model group at all time points (P< 0.05). Hes1 immunoreactivity was suppressed in the DAPT group, with significant differences between DAPT and model groups at 3, 7, and 14 days (P<0.05). Hes1 immunoreactivity was inhibited in both the acupuncture and DAPT groups, but no signifcant difference was detected at any time point (P> 0.05; Table 3, Figure 3).
discussion
Neural stem cells can self-renew and undergo multi-directional differentiation. Under certain conditions, they differentiate into neuronal or glial cells. The Notch signaling pathway is thought to be a main factor for embryonic neural stem cell growth regulation, playing a “side restrain”role (Artavanis-Tsakonas et al., 1999; Struhl, 1999; Shi et al., 2008). Specifically, the Notch signaling pathway indirectly maintains neural stem cell differentiation and self-renewal by inhibiting neuronal and glial cell differentiation (Androutsellis-Theotokis et al., 2006). Using chicken embryos, researchers have shown Notch’s “side restrain”, with the activated Notch signaling system inhibiting neuronal differentiation. Notch signaling pathway activation causes restraint of neural stem cell differentiation, and instead they proliferate. (Wakamatsu et al., 2000).
Our western blot results showed that Notch1 expression increased signifcantly at 1 day after modeling, peaked at 3 days, reduced at 7 days, and then obviously decreased and was close to normal levels at 14 days. Notch1 expressionwas visibly lower in the acupuncture and DAPT groups than the model group at various time points. DAPT is an inhibitor of the Notch signaling pathway.In vivoandin vitroexperiments show that DAPT application blocks the Notch signaling pathway and causes G1/S phase cell cycle delay in human embryonic stem cells, inducing neuronal cell differentiation and significantly reducing the time required for neural maturation (Borghese et al., 2010). In the present study, we found no signifcant difference between the acupuncture and DAPT groups, indicating that acupuncture plays a similar role to DAPT, and inhibits Notch1 and Hes1 expression and indirectly maintains neural stem cell self-renewal. Our immunohistochemical staining results are consistent with the observed western blot trends at 1 and 3 days. Notch1 immunoreactivity in the acupuncture group was signifcantly weaker than the model group at 7 days, and also weaker than the DAPT group, suggesting that acupuncture inhibits Notch signaling pathway expression. The Notch-Hes signaling pathway effect on neural stem cells, as well as our western blot and immunohistochemistry results, suggests that acupuncture may inhibit neuronal differentiation of neural stem cells by inhibiting the Notch-Hes signaling pathway and maintaining neural stem cell proliferation. This assumption needs further research, but offers new research directions for neural stem cell regeneration and repair.
Acknowledgments:We are very grateful to all the staff of Harbin Medical University in China for their help.
Author contributions:WZ designed research and XWS and QBC performed research. QXC wrote the manuscript. HYK gave technical or material support. XPY and XHD analyzed data. All authors approved the final version of the paper .
Conficts of interest:None declared.
Androutsellis-Theotokis A, Leker RR, Soldner F, Hoeppner DJ, Ravin R, Poser SW, Rueger MA, Bac SK, Kittappa R, McKay RD (2006) Notchsinalling regulates stem cell numbers in vitro and in vivo. Nature 442:823-826.
Artavanis-Tsakonas S, Rand MD, Lake RJ (1999) Notch signaling:cell fate control and signal integration in development. Science 284:770-776.
Bederson JB, Pitts LH, Tsuji M, Nishimura MC, Davis RL, Bartkowski H (1986) Rat middle cerebral artery occlusion: evaluation of the model and development of neurologic examination. Stroke 17:472-476.
Borghese L, Dolezalova D, Opitz T, Haupt S, Leinhaas A, Steinfarz B, Koch P, Edenhofer F, Hampl A, Brustle O (2010) Inhibition of Notch signaling in human embryonic stem cell-deribed neural stem cells delays G1/S phase transition and accelerates neuronal differentiation in vitro and invivo. Stem Cells 28:955-964.
Chamnanvanakij S, Margraf LR, Burns D, Perlman JM (2002) Apoptosis and white matter injury in preterm infants. Pediatr Dev Pathol 5:184-189.
Chapouton P, Skupien P, Hesl B, Coolen M, Moore JC, Madelaine R, Kremmer E, Faus-Kessler T, Blader P, Lawson ND, Bally-Cuif L (2010) Notch activity levels control the balance between quiescence and recruitment of adult neural stem cells. J Neurosci 30:7961-7974.
Falk R, Falk A, Dyson MR, Melidoni AN, Parthibam K, Young JL, Roake W, McCafferty J (2012) Generation of anti-notch antibodies and their application in blocking notch signalling in neural stem cells. Methods 58:69-78.
Feng X, Yang S, Liu J, Huang J, Peng J, Lin J, Tao J, Chen J (2013) Electroacupuncture ameliorates cognitive impairment through inhibition of NF-kappaB-mediated neuronal cell apoptosis in cerebral ischemia- reperfusion injured rats. Mol Med Rep 7:1516-1522.
Fernandez-Valdivia R, Takeuchi H, Samarghandi A, Lopez M, Leonardi J, Haltiwanger RS, Jafar-Nejad H (2011) Regulation of mammalian Notch signaling and embryonic development by the protein O-glucosyltransferase Rumi. Development 138:1925-1934.
Gao C, Liu W, Sun ZD, Zhao SG, Liu XZ (2009) Atorvastatin ameliorates cerebral vasospasm and early brain injury after subarachnoid hemorrhage and inhibits caspase-dependent apoptosis pathway. BMC Neurosci doi: 10.1186/1471-2202-10-7.
Garcia A, Kandel JJ (2012) Notch: a key regulator of tumor angiogenesis and metastasis. Histol Histopathol 27:151-156.
Gianni-Barrera R, Trani M, Reginato S, Banf A (2011) To sprout or to split? VEGF, Notch and vascular morphogenesis. Biochem Soc Trans 39:1644-1648.
Guan XL, Hou LC, Yang H, Wang FX, He M (2013) Glutamate in rat brain hemorrhage in brain water content and apoptosis changes and basic fbroblast growth factor. Zhongfeng yu Shenjing Jibing Zazhi 30:325-328.
Guo D, Li C, Teng Q, Sun Z, Li Y, Zhang C (2013) Notch1 overexpression promotes cell growth and tumor angiogenesis in myeloma. Neoplasma 60:33-40.
Guo Y, Zhao NJ, Lin L, Zhang MF, Zheng ZH (2010) Notch1 gene on human U251 glioma cell proliferation and cycle. Zhongguo Bingli Shengli Zazhi 26:1115-1119.
Imayoshi I, Shimogori T, Ohtsuka T, Kageyama R (2008) Hes genes and neurogenin regulate non-neural versus neural fate specifcation in the dorsal telencephalic midline. Development 135:2531-2541.
Li X, Wang Q (2013) Acupuncture therapy for stroke patients. Int Rev Neurobiol 111:159-179.
Miele L (2006) Notch signaling. Clin Cancer Res 12:1074-1079.
Monahan P, Rybak S, Raetzman LT (2009) The Notch target gene Hes1 regulates cell cycle inhibitor expression in the developing pituitary. Endocrinology 150:4386-4394.
Nam MH, Ahn KS, Choi SH (2013) Acupuncture stimulation induces neurogenesis in adult brain. Int Rev Neurobiol 111:67-90.
Nickoloff BJ, Osborne BA, Miele L (2003) Notch signaling as a therapeutictarget in cancer: a new approach to the development of cell fate modifying agents. Oncogene 22:6598-6608.
Riggs JE, Libell DP, Brooks CE, Hobbs GR (2005) Impact of institution of a stroke program upon referral bias at a rural academic medical center. J Rural Health 21:269-271.
Rosenberg GA, Mun-Bryce S, Wesley M, Kornfeld M (1990) Collagenase-induced intracerebral hemorrhage in rats. Stroke 21:801-807.
Shi Y, Sun G, Zhao C, Stewart R (2008) Neural stem cell self-renewal. Crit Rev Oncol Hematol 65:43-53.
Struhl G (1999) Greenwald l Presenilin is required for activity and nuclear access of Notch in Drosophila. Nature 398:522-524.
Sun Y, Nadal-Vicens M, Misono S, Lin MZ, Zubiaga A, Hua X, Fan G, Greenberg ME (2001) Neurogenin promotes neurogenesis and inhibits glial differentiation by independent mechanisms. Cell 104:365-376.
Wakamatsu Y, Maynard TM, Weston JA (2000) Fate determination of neural crest cells by Notch-mediated lateral inhibition and asymmetrical cell division during gangliogenesis. Development 127:2811-2821.
Wang Z, Li Y, Banerjee S, Kong D, Ahmad A, Nogueira V, Hay N, Sarkar FH (2010) Down-regulation of Notch-1 and Jagged-1 inhibits prostate cancer cell growth migration and invasion, and induces apoptosis via inactivation of Akt-mTOR and NF-κB signaling pathways. J Cell Biochem 109:726-736.
Yagi H, Saito T, Yanagisawa M, Yu RK, Kato K (2012) Lewis X-carrying N-glycans regulate the proliferation of mouse embryonic neural stem cells via the Notch signaling pathway. J Biol Chem 287:24356-24364.
Yalcin-Ozuysal O, Fiche M, Guitierrez M, Wagner KU, Raffoul W, Brisken C (2010) Antagonistic roles of Notch and p63 in controlling mammary epithelial cell fates. Cell Death Differ 17:1600-1612.
Zhang XM, Huang GW, Liu H, Chang H, Wilson JX (2012) Folic acid enhances Notch signaling, hippocampal neurogenesis, and cognitive function in a rat model of cerebral ischemia. Nutr Neurosci 15:55-60.
Copyedited by James R, Wysong S, Wang J, Qiu Y, Li CH, Song LP, Zhao M
*
10.4103/1673-5374.153696
http://www.nrronline.org/
Accepted: 2014-11-12

- 中国神经再生研究(英文版)的其它文章
- RAFting the rapids of axon regeneration signaling
- TAM receptors: two pathways to regulate adult neurogenesis
- Synapsing with NG2 cells (polydendrocytes), unappreciated barrier to axon regeneration?
- Targeting the body to protect the brain: inducing neuroprotection with remotely-applied near infrared light
- Novel advancements in threedimensional neural tissue engineering and regenerative medicine
- Functional regeneration of the brain: white matter matters