
不同覆盖栽培年限雷竹林生物量分配格局研究
2015-01-17 03:54高贵宾潘雁红吴良如徐旻昱
江西农业大学学报 2015年4期
高贵宾,潘雁红,吴良如,钟 浩,袁 娜,徐旻昱
(1.国家林业局竹子研究开发中心,浙江 杭州 310012;2.余杭区林业水利局,浙江 余杭 311100)
不同覆盖栽培年限雷竹林生物量分配格局研究
高贵宾1,潘雁红1,吴良如1,钟 浩1,袁 娜2,徐旻昱2
(1.国家林业局竹子研究开发中心,浙江 杭州 310012;2.余杭区林业水利局,浙江 余杭 311100)
对不同覆盖栽培年限雷竹林生物量分配格局进行了研究,结果发现:随覆盖年限增加,新竹留养困难,歇园期间新竹留养不规律,导致竹林年龄结构不稳定,表现出1年、4年生竹子生物量下降,2年、3年生竹子生物量先升高后下降的总趋势,进而影响竹林年龄结构的人工可控性。竹林生物量在覆盖3年时有所升高,竹叶生物量的累积,应该是竹林仍能保持较高生产力的关键因素。竹叶生物量在竹林覆盖6年后开始有较大差异,到第12年已经跟覆盖0年、3年竹林达到显著差异水平,竹叶生物量的衰减成为长期覆盖竹林生产力衰退的重要限制因子。竹林覆盖到第3年,地下竹鞭根生物量才达到或接近饱和状态,说明竹林覆盖后虽然竹鞭有所上浮,但仍然有能力实现空间拓殖;继续覆盖,竹鞭上浮增大,生存空间缩小,竹鞭的萌发与生长受到限制,生物量逐渐衰减。竹鞭根是土壤养分主要吸收和运输器官,其生物量的衰减,势必会减弱竹林系统水分代谢及矿质营养等能力,进而影响竹林生长。研究认为,建立稳定的竹林年龄结构,提高留养新竹子质量,及时翻土深埋鞭,并结合可行的退化土壤生态修复技术,才能有效防止竹林生物量衰减,实现竹林可持续经营。
雷竹;覆盖栽培;生物量;分配格局
高贵宾,潘雁红,吴良如,等.不同覆盖栽培年限雷竹林生物量分配格局研究[J].江西农业大学学报,2015,37(4):663-669.
笋用雷竹林覆盖栽培经营取得了良好的经济效益,然而,在获得巨大经济效益的同时,竹林也表现出一系列问题,如水土流失、地力衰退、生产力下降、竹林开花等[1-3]。众多研究者围绕这些问题进行了研究,重点主要集中于竹林衰退特征、竹林复壮、竹林覆盖栽培技术改良、完善并提高经营等级等[4-7]方面。生物量是考察生态系统中种群生长状况的重要指标[8-10],目前关于覆盖栽培雷竹林生物量方面的研究主要有雷竹林地上生物量估算[11-12]、退化竹林林分结构及地上生物量特征[13]、覆盖竹林细根生物量及养分含量变化[14]等,研究相对较少,未形成系统性。基于此,作者对不同覆盖栽培年限雷竹林生物量分配格局进行了研究,通过比较不同时间序列人工集约经营对竹林生物量形成的干扰情况,探究竹林生物量分配格局对生存环境的适应性变化;分析不同时间尺度强度人工经营条件下竹林生物量变化趋势,试图揭示竹林生态系统结构与功能的动态变化对竹林生长造成的影响,以期为现代竹林人工经营和衰退竹林的生态修复技术研究提供理论依据。
1 材料与方法
1.1 材料
于2014年10月在浙江余杭径山镇覆盖栽培雷竹林开展生物量调查。分别选择立地条件和经营水平相近的不同覆盖栽培年限:覆盖0年、覆盖3年、累积覆盖6年、累积覆盖9年、累积覆盖12年竹林作为调查样地。每样地设置3个100 m2样方。各样地竹林覆盖之初,采取连续覆盖3年,歇园1年的方式,之后连续覆盖2年,歇园1年。覆盖物以麦糠为发酵层,覆盖厚度10 cm;以砻糠为保温层,厚度15 cm。各样地(除覆盖0年竹林)立竹度基本为16 000株/hm2,只留养1~4年生竹株。覆盖0年竹林为新造3年林,年龄结构数量比例为1年生竹∶2年生竹∶3年生竹为1∶1∶1,立竹度12 000株/hm2。覆盖3年以上竹林由于留笋养竹困难及歇园时竹子留养不规律等因素而难以保持稳定的年龄结构。整个竹林胸径分布在2.50~3.50 cm。
1.2 方法
立竹生物量调查:检尺各覆盖年限样地样方中不同年龄竹株胸径,分别随机选取样方中平均胸径大小的各年龄段、生长势良好的健康竹株3株作为标准竹,则每样地3个100 m2样方中每个年龄段共计有9株标准竹,每样地所有年龄段标准竹合计36株(覆盖0年竹林27株)。将标准竹齐地伐倒,称量杆、枝、叶鲜重;采挖其竹篼,清除其竹根,洗净、晾干,称鲜重。
竹鞭根生物量调查:在每种样地样方中,分别随机采挖3处1 m3小样方,则每样地共计有9个1 m3小样方,以10 cm为区间分层,将各土层所得竹鞭和鞭根洗净、晾干,称鲜重。
所得数据在Excel和SPSS17.0软件中进行作图和统计分析,其中覆盖竹林各指标数据相互间差异显著性比较,采用SNK检验方法(即q检验)。立竹生物量以鲜重计。
2 结果与分析
2.1 竹林立竹生物量分配格局
2.1.1 各覆盖年限不同年龄立竹不同部位生物量变化 由图1~图4可见:覆盖0年竹林竹子各部位生物量随年龄增大有总体下降趋势,说明新造林过程中,为保证竹子成活率,所选母竹径级较小,后随竹林不断更新,径级有所增加,竹子生长势逐渐增强。图中,覆盖3~12年竹林各部位生物量均为2年、3年生竹子总生物量大于1年、4年生竹子总量,说明竹林歇园期间大量留养了新竹,随老竹子砍伐及新竹子更新,而成为占有较大生物量的中间年龄竹子。从同一年龄竹子不同部位生物量随覆盖年限增长的变化来看,有1年、4年生竹子生物量下降,2年、3年生竹子生物量先升高后下降的总趋势,反映了竹林覆盖期间留笋养竹困难、覆盖后留养的新竹质量相对较差,随覆盖年限增长,竹林更新质量总体有所下降,但在竹林歇园期间又大量留养新竹,所以会产生2年、3年生竹子生物量先升高后下降的现象。说明随覆盖年限增加,竹林年龄结构不容易稳定,从而会影响竹林结构的人工可控性。
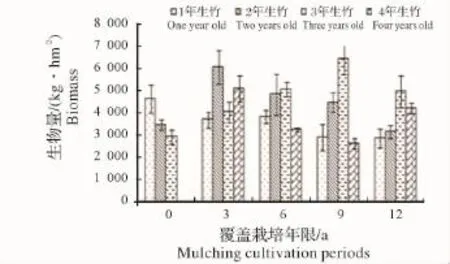
图1 竹竿生物量变化Fig.1 Variations of biomass of bamboo culm
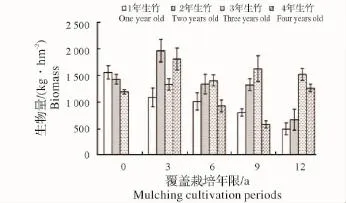
图2 竹枝生物量变化Fig.2 Variations of biomass of bamboo branch
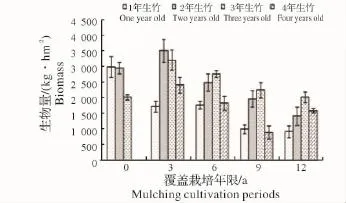
图3 竹叶生物量变化Fig.3 Variations of biomass of bamboo leaf
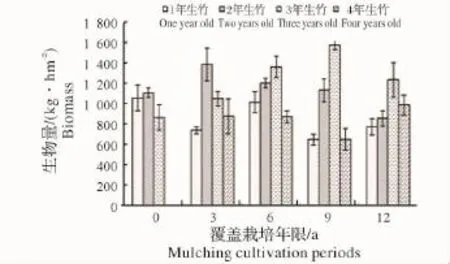
图4 竹篼生物量变化Fig.4 Variations of biomass of bamboo pocket
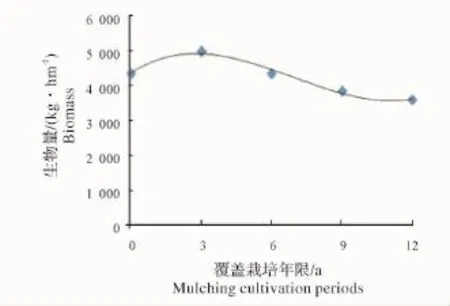
2.1.2 立竹不同部位生物量随覆盖年限的变化 如图5所示:竹秆生物量在覆盖3年时达最高点,之后随覆盖年限增长开始下降;竹枝、竹叶生物量在覆盖0年、3年平缓升高,之后开始下降;各覆盖年限竹林竹篼生物量变化不大。从图中看出,各部分生物量高低水平是竹秆>竹叶>竹枝>竹篼。整个竹林地上生物量随竹林覆盖年限增长的变化趋势(图6)显示,覆盖3年时生物量有所升高,之后不断下降。这一方面说明新造竹林3~4年后虽然开始覆盖,但整个竹林的生产力水平仍然较高,生物量不断累积,竹子各部分生物量的累积以竹秆最大,竹叶次之。因竹叶是整个竹林系统的有机营养制造者,在覆盖3年的竹林中,竹叶生物量的累积,应该是竹林仍能保持较高生产力的关键因素;另一方面说明随覆盖年限增长,竹林地上生物量发生衰减,其竹林生产力可持续性将受到限制。
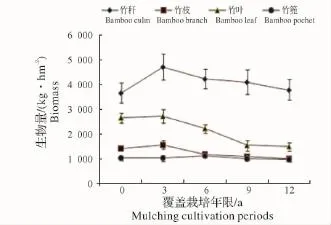
图5 竹子不同部位生物量随覆盖年限的变化Fig.5 Biomass variations of different parts of bamboo in different eriods of mulching
2.1.3 立竹生物量分配比较 表1可见:各覆盖年限竹林竹株不同部位生物量分配比率:竹秆>竹叶>竹枝>竹篼,竹秆、竹叶总比率均在70%以上,是构成竹林地上部分生物量的主要部分;竹子同一部位生物量随覆盖年限的增长,竹秆、竹篼分配比率有所增大,竹枝、竹叶有所减小。通过对各覆盖年限竹子不同部位及整个竹林竹株生物量进行SNK检验(即q检验)得出(表2):不同覆盖年限间竹秆、竹枝、竹篼生物量差异水平并不显著,竹叶在竹林覆盖6年后开始有差异,到第12年竹林竹叶生物量已经跟覆盖0年、3年竹林达到显著差异水平,这说明竹叶生物量的衰减是长期覆盖竹林生产力衰退的重要限制因子。整个竹林竹株生物量,覆盖3年跟覆盖0年(为3年新造林,竹林郁闭度较小,生物量总体水平较低)、6年达显著差异水平,跟覆盖9年、12年竹林达极显著差异水平,覆盖0年、6年竹林跟覆盖9年、12年竹林也达显著差异水平,说明虽然在竹林经营过程中采取了一些恢复技术,但竹林衰退仍然无法避免,其恢复技术有较大的提升空间。
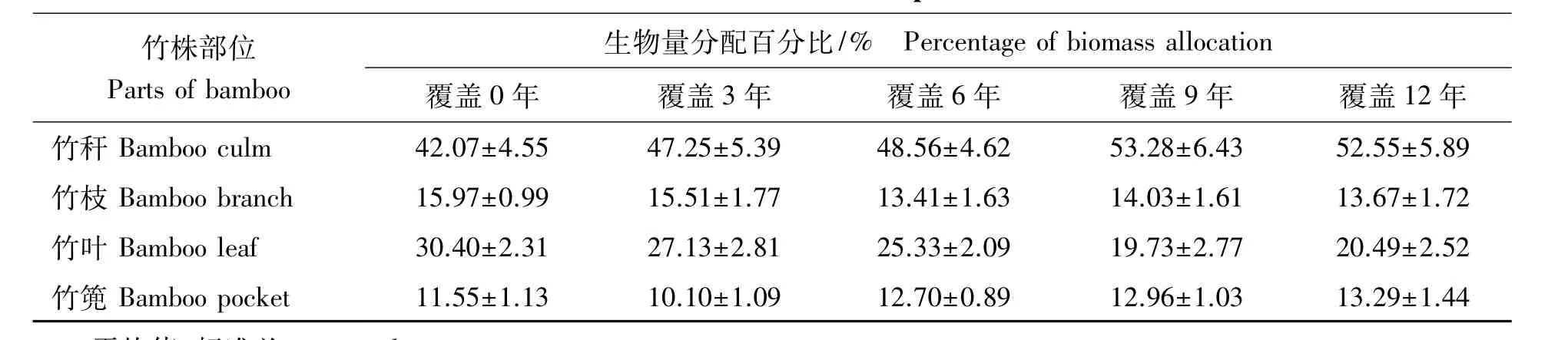
表1 竹子不同部位生物量分配Tab.1 Biomass allocation of different parts of bamboo
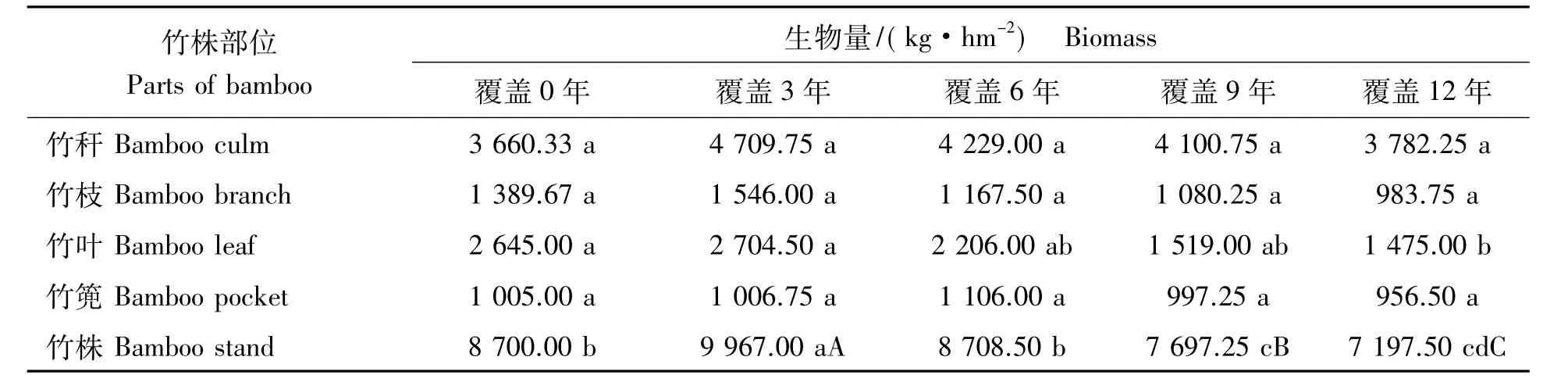
表2 竹子不同部位生物量差异Tab.2 Biomass differences of different parts of bamboo
2.2 竹林地下竹鞭根生物量分配格局
2.2.1 各覆盖年限不同土层地下竹鞭根生物量变化 如图7、图8所示:覆盖之初竹鞭根集中分布于20~40 cm土层,其中以30 cm土层分布相对较多,随覆盖年限增长,主要分布于10~30 cm土层,其中以20 cm土层分布相对较多,到第12年,30 cm土层以下已基本没有竹鞭根分布,说明覆盖年限越长,竹鞭根上浮程度越大。这与地表覆盖物清除不彻底及竹鞭根趋肥性有极大关系。各覆盖年限同一土层竹鞭根生物量在覆盖3年竹林均有较高累积,之后均有所下降,说明竹林覆盖到第3年,地下土层竹鞭根分布才达到或接近饱和状态,竹林覆盖后虽然竹鞭整体上浮,但仍然有能力实现空间拓殖,这可能也与竹林土壤退化较慢有关。
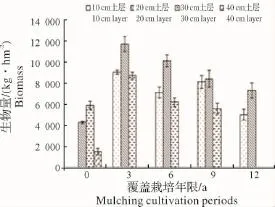
图7 不同土层竹鞭生物量变化Fig.7 Biomass variation of bamboo rhizome in different soil layer
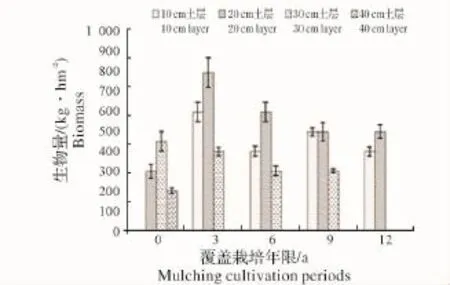
图8 不同土层鞭根生物量变化Fig.8 Biomass variation of bamboo root in different soil layer
2.2.2 地下竹鞭根生物量随覆盖年限的变化 竹林地下竹鞭及鞭根生物量随覆盖年限增长逐渐累积,第3年达顶峰,之后逐渐衰减(图9、图10),这与各覆盖年限地下鞭根总生物量随覆盖年限增长的变化趋势(图11)相吻合。虽然竹林经过了3年覆盖,但其生物量积累的持续性并没有被打断,这可以解释竹林为保证自身生存发展,对人工经营干扰存在某种程度的“拮抗”作用。竹鞭根是竹林生长所需水分及矿质元素等营养成分的主要吸收器官,其生物量的衰减,势必会减弱竹林系统水分代谢及矿质营养等能力,进而影响到整个竹林生长。
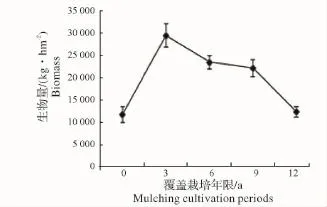
图9 竹鞭生物量随覆盖年限的变化Fig.9 Biomass variation of bamboo rhizome in different periods of mulching
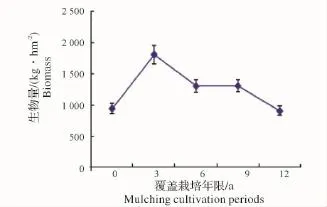
图10 鞭根生物量随覆盖年限的变化Fig.10 Biomass variation of bamboo root in different periods of mulching
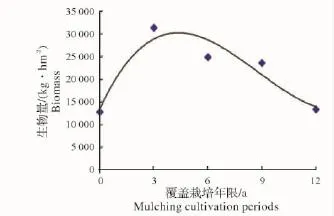
图11 地下鞭根生物量与覆盖年限的关系Fig.1 Relationship between rhizome,root biomass and mulching period
2.2.3 竹林地下竹鞭根生物量分配比较 由表3可见:各覆盖年限竹鞭及鞭根生物量分配比率均以竹鞭占绝对优势,各覆盖年限竹鞭生物量分配比率在92%~95%,变化不大,鞭根生物量则以覆盖0年占比率最高,说明竹林地下营养吸收器官在容量较大的土壤空间中,可以通过扩大鞭根生物量辐射范围,以更好的吸收和利用土壤养分。通过对各覆盖年限竹鞭根生物量进行SNK检验(即q检验)发现:覆盖3年竹林地下竹鞭根生物量与各覆盖年限竹林均达极显著差异水平(覆盖0年竹林地下竹鞭根生长尚未饱和,可另作他论)。到覆盖12年,竹林地下竹鞭根已经极大衰减,说明竹鞭上浮,其生存空间缩小,已经严重影响竹鞭的萌发与生长,此时的竹林只能翻土深埋鞭,才能防止竹鞭根生物量衰减。
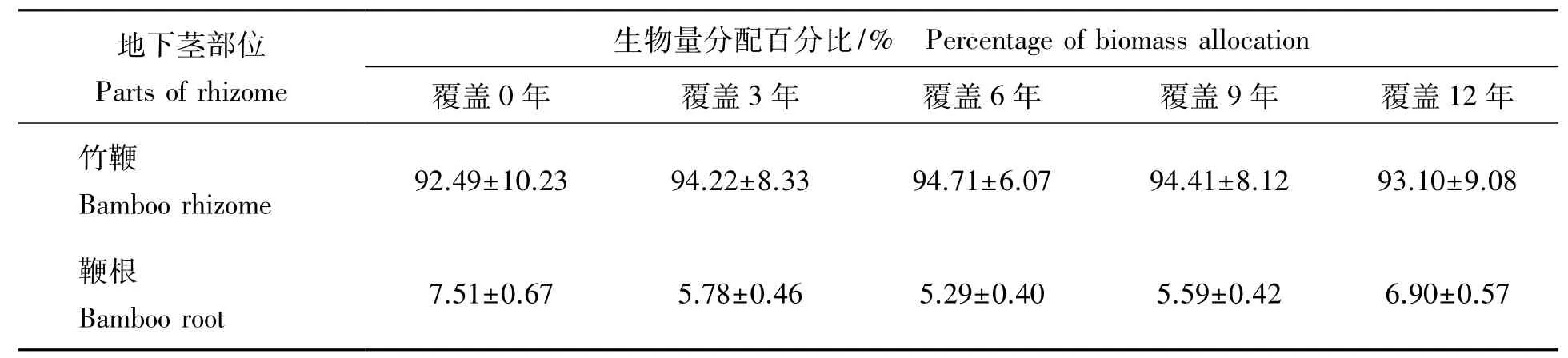
表3 竹林地下竹鞭根生物量分配Tab.3 Biomass allocation of rhizome and root of bamboo forests
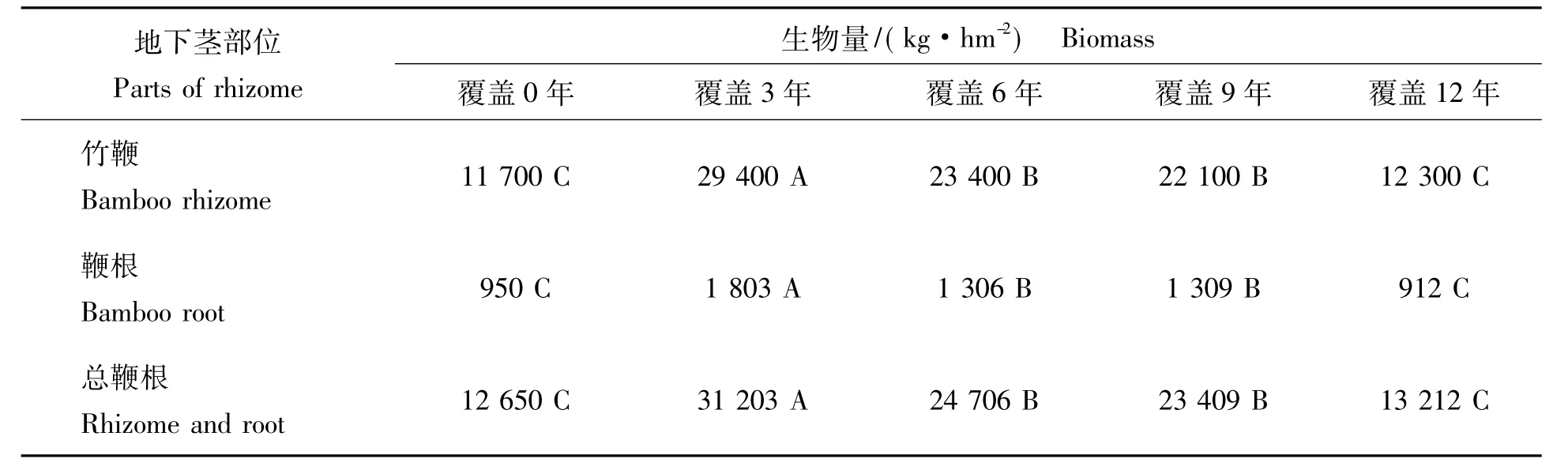
表4 竹林地下竹鞭根生物量差异Tab.4 Biomass differences of rhizome and root of bamboo forests
3 结论与讨论
本实验结果表明:随覆盖年限增加,竹笋留养困难,歇园期间竹笋留养不规律,使竹林年龄结构不容易稳定,表现出1年、4年生竹子生物量下降,2年、3年生竹子生物量先升高后下降的总趋势,进而影响竹林结构的人工可控性。竹林生物量在覆盖3年时有所升高,竹叶生物量的累积,应该是竹林仍能保持较高生产力的关键因素,之后发生衰减,竹林生产力可持续性受到限制。竹叶在竹林覆盖6年后开始有差异,到第12年竹林,竹叶生物量已经跟覆盖0年、3年竹林达到显著差异水平,竹叶生物量的衰减是长期覆盖竹林生产力衰退的重要限制因子。多重比较结果说明虽然在竹林经营过程中采取了一些恢复技术,但竹林衰退仍然无法避免,其恢复技术有较大的提升空间。根据研究结果,发现需解决的首要问题是建立稳定的竹林年龄结构,提高留养新竹质量,防止竹林生物量衰减。这就需要改变传统竹林覆盖经营模式,避免竹林歇园期间为单纯保持竹林立竹度而不规律多留养新竹的做法。所以我们基于正在进行的覆盖竹林与相连的未覆盖竹林生理整合作用的研究,提出一种新覆盖方法,常年覆盖,常年留养新竹,探究竹林生产力可持续性与经济效益的关系,并与传统竹林覆盖模式进行比较,最终建立一定面积的示范林,目前该研究正在进行中。
竹林覆盖到第3年,地下土层竹鞭根分布才达到或接近饱和状态,说明竹林覆盖后虽然竹鞭有所上浮,但仍然有能力实现空间拓殖,这可能也与竹林土壤退化较慢有关。长期覆盖经营,竹鞭上浮增大,生存空间缩小,竹鞭萌发与生长受到限制,竹鞭根生物量逐渐衰减。竹鞭根是土壤养分主要吸收器官,其生物量的衰减,势必会减弱竹林系统的水分代谢及矿质营养等能力,进而影响到整个竹林生长。此时的竹林需要翻土深埋鞭或加客土,才能防止因生存空间的局限性导致的竹鞭根生物量衰减。众多研究表明[15-17]:长期覆盖经营,竹林土壤发生严重退化。所以我们在翻土深埋鞭的同时,重点开展竹林土壤生态修复技术研究,用特制竹炭生物有机肥及竹屑发酵肥“还竹于竹”,配合施用生石灰,改善土壤环境,期望可以解决覆盖栽培雷竹林退化问题的技术瓶颈,实现竹林可持续经营。
参考文献:
[1]戎静,庄舜尧,杨浩.太湖源地区雷竹林氮磷径流输出与拦截控制[J].水土保持通报,2011,31(4):168-171.
[2]李子川.不同集约栽培年限下雷竹林土壤化学性质和铝形态研究[D].临安:浙江农林大学,2011.
[3]刘丽,陈双林,李艳红.基于林分结构和竹笋产量的有机材料覆盖雷竹林退化程度评价[J].浙江林学院学报,2010,27 (1):15-21.
[4]刘亚迪,范少辉,蔡春菊,等.地表覆盖栽培对雷竹林凋落物养分及其化学计量特征的影响[J].生态学报,2012,32 (22):6955-6963.
[5]张有珍,何钧潮,郑惠君.肥料种类及施肥深度对覆盖雷竹林的影响[J].浙江林业科技,2011,31(3):40-43.
[6]祝小祥,谢国雄,徐进,等.临安市雷竹林土壤肥力分析与培肥措施[J].中国农学通报,2013,29(28):72-76.
[7]可晓,陈双林,张小平,等.雷竹林存留有机覆盖物高效降解菌株分离及产酶条件优化[J].浙江农林大学学报,2012,29(2):244-250.
[8]谭勇,何东进,游巍斌,等.闽东滨海湿地入侵种互花米草表型可塑性及生物量分配对潮位梯度的响应[J].应用与环境生物学报,2014,20(5):863-868.
[9]毛伟,李玉霖,崔夺,等.沙质草地不同生活史植物的生物量分配对氮素和水分添加的响应[J].植物生态学报,2014,38(2):125-133.
[10]韩梅,杨利民,张永刚,等.中国东北样带羊草群落C3和C4植物功能群生物量及其对环境变化的响应[J].生态学报,2006,26(6):1825-1832.
[11]徐小军,周国模,杜华强,等.基于Landsat TM数据估算雷竹林地上生物量[J].林业科学,2011,47(9):1-6.
[12]董德进,周国模,杜华强,等.6种地形校正方法对雷竹林地上生物量遥感估算的影响[J].林业科学,2011,47(12):1-8.
[13]李艳红,陈双林,刘丽,等.林地有机材料覆盖退化雷竹林地上部分生物量研究[J].中国农学通报,2009,25(8):102-107.
[14]董建.林地覆盖措施对雷竹细根生物量及养分含量的影响[D].雅安:四川农业大学,2011.
[15]郭子武,俞文仙,陈双林,等.林地覆盖对雷竹林土壤微生物特征及其与土壤养分制约性关系的影响[J].生态学报,2013,33(18):5623-5630.
[16]徐祖祥,陈丁红,李良华,等.临安雷竹种植条件下土壤养分的变化[J].中国农学通报,2010,26(13):247-250.
[17]季海宝,孙晓,桂仁意,等.集约经营对雷竹林土壤与植株铝含量的影响[J].林业科学,2014,50(1):15-20.
A Study on Biomass Allocation Pattern of Phyllostachys praecox in Different Mulching Cultivation Periods
GAO Gui-bin1,PAN Yan-hong1,WU Liang-ru1,ZHONG Hao1,YUAN Na2,XU Min-yu2
(1.Bamboo Research and Development Center,State Forestry Bureau,Hangzhou 310012,China;2.Yuhang Forestry and Water Conservancy Bureau,Yuhang 311100,China)
The biomass allocation patterns of Phyllostachys praecox in different mulching cultivation periods were studied.The results showed that following the increase of mulching cultivation periods,the keeping of new bamboo became difficult,and with the irregular keeping of new bamboo in the paused mulching cultivation period,the age structure of bamboo forest became unstable,which exhibited an general trend that the biomass of one year old and four years old bamboo decreased,two years old and three years old bamboo firstly increased and then decreased,which further influenced the management controllability of bamboo age structure.Bamboo biomass increased in the 3-year mulched forest,the accumulation of bamboo leaf biomass might be the key factorfor the high productivity of bamboo forest.Bamboo leaf biomass had greater variation after mulching for 6 years,in the 12-year mulched bamboo forest,there were significant differences compared with the 0-year and 3-year mulched forests,the attenuation of bamboo leaf biomass became an important limiting factor to the declining of bamboo forest productivity.After mulching for 3-year,the biomass of bamboo rhizome just reached the saturation state.This showed that,although the bamboo rhizome floated upward after mulching cultivation,it could still had the ability to occupy soil space.If mulching cultivation continued,bamboo rhizome floated upward constantly,with living space reduced,the germination and growth of bamboo rhizome were limited,and then the biomass decreased gradually.Bamboo rhizome was the main organ for absorption and transportation of soil nutrient,the attenuation of its biomass would certainly weaken the abilities of water metabolism and mineral nutrition of bamboo forest system,which further influenced the growth of bamboo forest.It is concluded that,a stable bamboo age structure,improved quality of new bamboo,deep soil and timely burial of bamboo rhizome,combined with the practical technology of ecological restoration of degraded soil might be able to prevent the attenuation of bamboo biomass effectively and then realize the sustainable management of bamboo forest.
Phyllostachys praecox;mulching cultivation;biomass;allocation pattern
S795.9.02
A
1000-2286(2015)04-0663-07
2014-12-04
2015-01-27
中央科研院所所长基金(ZXJJ201203)和中央级公益性科研院所基本科研业务费专项资金(CAFYBB2014QA038)
高贵宾(1982—),男,助理研究员,主要从事竹类植物研究,E-mail:anshu998@163.com。
猜你喜欢
湖北林业科技(2021年4期)2021-09-07
林业科学(2020年12期)2021-01-26
少儿美术(2020年9期)2020-11-05
小哥白尼(趣味科学)(2020年1期)2020-06-16
陶山(2019年1期)2019-03-29
东坡赤壁诗词(2018年1期)2018-03-31
创新作文(1-2年级)(2017年5期)2017-12-07
世界竹藤通讯(2017年4期)2017-08-17
空中之家(2017年7期)2017-07-20
现代装饰(2017年12期)2017-05-22

- 江西农业大学学报的其它文章
- 莱芜猪种质资源特性研究
——脂肪沉积性状