
白桦子代家系幼林期生长表现及适应性分析
2015-01-11 06:33:08徐焕文李志新彭儒胜尚福强邹建军滕文华刘桂丰
浙江农林大学学报 2015年6期
刘 宇,徐焕文,李志新,姜 静,彭儒胜,尚福强,邹建军,滕文华,刘桂丰
(1.东北林业大学 林木遗传育种国家重点实验室,黑龙江 哈尔滨 150040;2.辽宁省杨树研究所,辽宁 盖州 115200;3.辽宁省森林经营研究所,辽宁 丹东 118002;4.吉林省林业科学研究院,吉林 长春130033)
白桦子代家系幼林期生长表现及适应性分析
刘 宇1,徐焕文1,李志新1,姜 静1,彭儒胜2,尚福强3,邹建军4,滕文华1,刘桂丰1
(1.东北林业大学 林木遗传育种国家重点实验室,黑龙江 哈尔滨 150040;2.辽宁省杨树研究所,辽宁 盖州 115200;3.辽宁省森林经营研究所,辽宁 丹东 118002;4.吉林省林业科学研究院,吉林 长春130033)
林木子代家系的多点试验是研究其适应性及稳定性的重要环节。以6个白桦Betula platyphylla三倍体家系、4个二倍体家系为材料,分别在尚志、辉南、凌海、丹东等4个试验点营造子代测定林,对3年生树高和地径等性状进行多地点联合分析。结果表明:树高在各试验点均达到差异显著水平(P<0.01);地径仅在丹东、凌海试验点各家系达到显著水平(P<0.05)。尚志试验点最优三倍体家系为314和319;辉南试验点为318和320;凌海试验点为314,316和320;丹东试验点为316,318和319。采用生产力指数法及基因型分组法将参试家系分为3种类型,即高产非稳产型家系,包括202,314和320;低产稳产型家系,包括201,318和319;低产非稳产型家系,包括203,204和315。各试验点入选的优良家系分别属于高产非稳产型家系及低产稳产型家系。研究结果不仅为白桦三倍体家系的早期推广提供参考,而且也为高世代三倍体制种园的改建提供重要依据。图3表6参24
林木育种学;白桦四倍体子代;多点试验;家系选择;遗传稳定性
植物多倍体是自然界中的普遍现象。自然界中大约35%的被子植物和70%的禾本科Gramineae植物均属于多倍体[1],植物染色体的多倍体化,有效推动了植物的进化,是新物种形成的重要途径之一[2]。植物的多倍体化往往也带来器官的巨大性,由于林木主要被利用的是营养器官,且多年生习性又保证品种一旦育成后就可以长期持续利用。因此,与其他植物相比,林木多倍体育种的潜力更大、作用更为突出。有人利用欧洲山杨Populus tremula四倍体与美洲山杨Populus tremuloides二倍体杂交,选育出了树冠狭窄、适应性强、生长快、抗锈病能力强的人工杂种三倍体新品种 ‘Astria’[3]。北京林业大学采用化学诱导技术获得2n花粉,再通过杂交育种的方法选育出了三倍体毛白杨Populus tomentosa优良新品种,其材积生长不仅较二倍体快[4],同时也表现出对不同环境高强的适应能力以及抗病能力[5]。由于林木三倍体优良新品种表现出速生性、抗逆性以及高度不育性的特点而越来越被重视,成为当今研究的热点之一。目前,已经在杨树Populus,橡胶树Hevea brasiliensis,漆树Toxicodendron vernicifluum,桑树Morus alba等多个树种中均选育出了优良三倍体新品种,并在生产上大量推广应用[6-13]。中国白桦Betula platyphylla倍性育种研究始于2004年,由本研究团队采用秋水仙素诱导白桦种子创制了白桦四倍体[14],并营建了初级白桦三倍体制种园。根据种子活力、生长性状初步选择了部分优良白桦四倍体母树[15-17]。在前期研究的基础上,针对营建的多点子代测定林进行稳定性和适应性分析,是白桦四倍体子代家系推广的重要环节。因此,研究团队2011年在东北三省的4个试验点开展了白桦三倍体家系的区域化试验,通过多地点联合分析确定各家系的稳定性和适应性,为今后白桦三倍体家系的推广提供依据,也为制种园的建园母树的改良提供参考。
1 材料与方法
1.1 试验材料与试验设计

表1 白桦杂交双亲及子代代码Table1 Birch hybrid parents and progeny codes
2009年在东北林业大学白桦强化种子园中,以定植于白桦强化种子园内的四倍体白桦Q65为母本, 6株二倍体杂种白桦(B.platyphylla×B.pendula)1-11,2-3,2-4,2-9,3-8和3-10等为父本,通过控制授粉获共获得6个四倍体家系的种子(314,315,316,318,319和320),同时采集园中4个二倍体家系(201,202,203和204)自由授粉的种子(表1)。2010年 4月末于塑料大棚中育苗,2011年早春分别在黑龙江省尚志市帽儿山实验林场、吉林省辉南县石道河林场、辽宁省凌海县金城良种场、丹东市五龙背新建村等4个试验点开展造林试验(表2),试验林按完全随机区组设计,20株小区,4次重复,株行距2 m×2 m。2012年秋季进行全林树高、地径及保存率调查。

表2 各试验点的地理气候条件Table2 Soil types and climatic factors of the test sites
1.2 数据处理
1.2.1 方差分析 运用Microsoft Excel进行数据处理,用SPSS.v16.0软件进行方差分析及多重比较。多点联合方差分析的线性模型为:xijk=μ+υj+ρjk+τi+(τυ)ij+εijk。其中:xijk第i个家系在第j个地点第k区组的平均观测值;μ为群体的平均值;υj为地点j的效应值;ρjk为地点内的区组效应(随机效应);τi为家系i的效应值;(τυ)ij为家系×地点的互作效应;εijk为随机误差。
各试验地点内采用双因素方差分析,线性模型为:xijk=μ+τi+ρk+εijk。其中:xijk为第i个家系在第k区组的平均观测值;ρk为区组效应(随机效应);εijk为随机误差。

1.2.2 隶属函数法 隶属函数值R(X)计算公式:R(Xi)=(Xi-Xmin)/(Xmax-Xmin),反隶属函数值R(X)计算公式:R(Xi)=1-(Xi-Xmin)/(Xmax-Xmin)。其中:Xi为指标测定值,Xmin,Xmax为所有参试材料某一指标的最小值和最大值。
1.2.3 遗传稳定性分析 ①回归分析。以各家系在各试验点的平均地径与每一个试验点所有家系的地径平均值(即环境指数)的线性回归系数(bi)估计遗传稳定性。当bi=1时,认为家系具有平均稳定性;当bi<1时,表示该家系高于平均稳定性;当bi>1时,表示家系低于平均稳定性[18-19]。计算公式为:
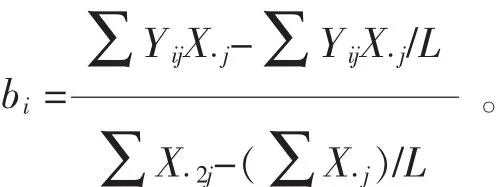
其中:bi表示第i家系回归系数;Yij表示第i家系在j个参试点某性状的平均值;X.j表示第j个参试点的环境指数;L表示各参试点个数。②Francis&Kannenberg基因型分组。以家系地径平均值为纵坐标,变异系数为横坐标绘制直方图,并以所有参试家系地径的总平均值和平均变异系数将其分为4组(图1)。第Ⅰ组属高产稳产型家系,第Ⅱ组是高产非稳产型家系,第Ⅲ组是低产稳产型家系,第Ⅳ组是低产非稳产型家系[20-21]。其中:家系地径平均值(Yi)与变异系数(CV)计算如下:


图1 家系稳定性分组Figure 1 Family stability group
其中:Yij表示第i家系在j个参试点地径的平均值;L表示各参试点个数。③生产力指数评价。生产力指数(IPQ)评价法:即生产力指数越大,表示该家系生长越快,反之越慢[22]。计算公式为:
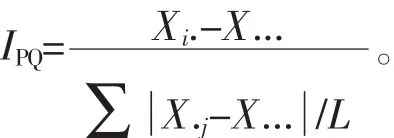
其中:Xi-X…为家系效应,是所有试验点i家系的地径平均值与所有试验点所有家系的平均值的离差;Σ∣X.j-X…∣为环境效应,是指各家系在j个参试点上的地径平均值与家系总平均值离差的绝对值除以参试点数的商。
2 结果与分析
2.1 参试家系树高和地径及保存率的多点联合方差分析
对4个试验点10个参试家系的树高、地径以及保存率进行联合方差分析表明(表3),在地点间各家系树高、地径、保存率差异均达到极显著水平(P<0.01);在家系间的树高、地径、保存率也达到差异显著(0.01<P<0.05)与极显著水平(P<0.01);树高与地径在家系与地点的交互作用间达到显著水平(0.01<P<0.05)。这一结果说明白桦家系在不同地点或不同的立地条件上生长表现与保存情况均不一致;基因型与环境之间存在互作。因此有必要开展区域性试验,为白桦三倍体优良家系的推广提供依据。

表3 参试家系生长性状及保存率的多点联合方差分析Table3 Analysis of variance among birch families for growth traits and preservation rate for the tested lines at different sites
2.2 各家系在不同试验地点的生长性状及保存率变异情况
采用双因素方差分析模型对各试验点10个家系的树高和地径进行方差分析(表4),结果显示:在树高生长方面,4个试验点的家系间差异均达到显著水平(0.01<P<0.05)或极显著水平(P<0.01);而地径生长只有凌海、丹东试验点的家系间差异达到极显著水平(P<0.01)。
进一步分析发现,在年均温较高的辽宁省丹东和凌海2个试验点无论是树高还是地径生长均显著高于其他试验点,家系树高均值分别为1.85 m和1.80 m,地径均值分别为3.05 m和2.91 cm,而在黑龙江尚志市的帽儿山试验点,年均温仅有2.4℃,树高和地径均值分别只有1.10 m和1.31 cm。总之,参试家系的生长呈现从北至南随着年均温的增加其生长量也增加的规律。
各参试试验点的保存率不尽相同(表4),辽宁省凌海试验点保存率最高,达到70%以上,而黑龙江尚志市的帽儿山试验点的保存率最低仅为31.13%,保存率在4个试验点的家系间差异均未达到显著水平(P>0.05),说明各家系在各地区保存情况基本一致。
10个家系在4个试验点的生长变异情况见表4。从树高来看,凌海试验点的变异系数最大,达到27.42%;其次是尚志试验点,其树高变异系数为24.34%;丹东试验点的变异系数最小,仅有17.79%。从地径来看,丹东试验点的地径变异系数最大,达到36.37%;其次是凌海,为34.78%;尚志试验点的地径变异系数最小,为23.23%。
4 个试验点的参试家系树高、地径多重比较表明:从整体来看,各试验点树高和地径排在前列的均为二倍体家系,有些三倍体家系虽然在生长表现方面逊色于二倍体家系,但各试验点表现较好的三倍体家系如318和319等与表现最好的二倍体家系差异尚未达到显著水平(表5)。这说明参试的三倍体家系虽然目前生长表现不是最优,但其较优良二倍体家系相差不大,今后通过更多优良四倍体母本与更多二倍体父本杂交,有望筛选获得优良三倍体家系以及适于生产优良三倍体子代的杂交双亲。
采用隶属函数法(表6)分别对4个试验点的参试家系生长优劣进行评价,若以隶属函数值大于0.45为入选优良家系, 则尚志试验点是203, 314, 319, 202和204家系; 辉南试验点是201,202,204,320和318家系;凌海试验点是202,320,314,203,204,201家系;丹东试验点大于该值的是204,202,203,318和316等家系。分析结果表明:参试家系的树高及地径综合评价在各个试验点并不完全一致,也说明不同家系在不同试验点的适应能力有所不同。

表4 各试验点参试家系生长性状及保存率比较Table4 Genetic parameters among birch families for growth traits and preservation rate for the tested lines at different sites
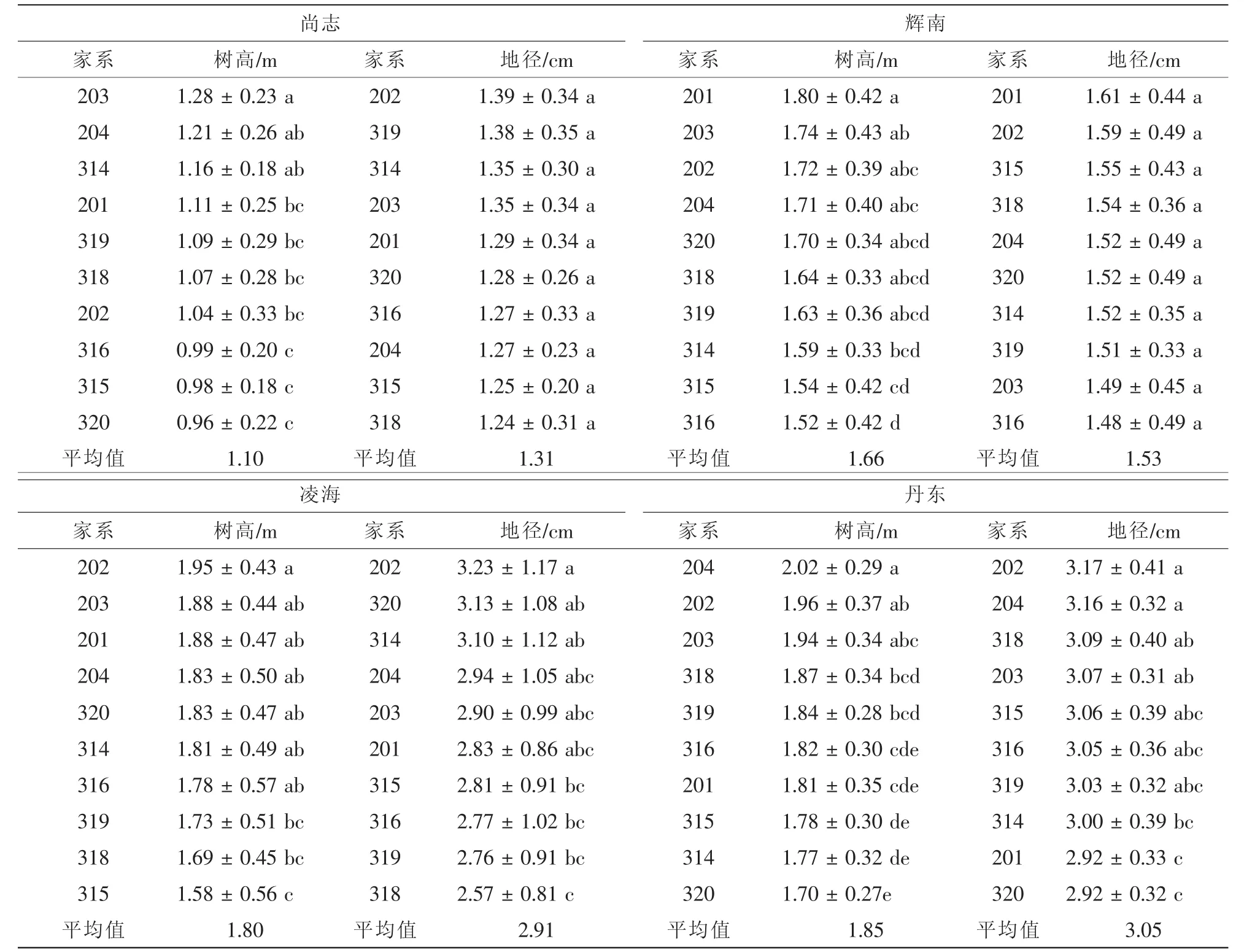
表5 各试验地点参试家系树高、地径多重比较Table5 Multiple comparisons of birch height and base diameter for the tested lines at different sites
2.3 生长稳定性的多点联合分析
由于地径性状在地点与家系间的交互作用达到了差异显著的水平(0.01<P<0.05),说明不同地点、不同家系存在较大差异,有必要开展生长稳定性分析。因此,分别采用生产力指数法以及基因型分组法对10个家系的遗传稳定性进行测定(图2和图3)。结果表明:除316家系外,上述2种方法区划结果基本一致。

表6 各试验地点参试家系隶属函数值Table6 Subordinate function values for the tested lines at different sites
据此,将参试家系分为3种类型,即基因型分组法的第Ⅱ类高产非稳产型家系,包括202,314和320家系,这些家系综合表现为具有较高的指标值,但遗传稳定性较差,随环境条件的变化而变化;第Ⅲ类为低产稳产型家系,包括201,318和319家系,这种类型的家系变异较小,具有良好的遗传稳定性,可以适应较为恶劣的环境,但其生产力水平较低;第Ⅳ类为低产非稳产型家系,包括203,204和315家系,这类家系突出表现为家系内变异较大,遗传稳定性较低,并且生产力也不高。上述2种方法分类结果基本一致,只有316家系被归为不同的类型,对该家系暂不进行生长稳定性的划分。
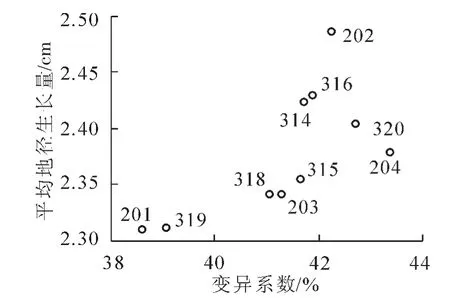
图2 基因型分组法对家系稳定性划分Figure 2 Gene grouping method for family stability group

图3 生产力指数与回归系数法对家系稳定性划分Figure 3 Productivity index and the regression coefficient for family stability group
3 结论与讨论
自开展白桦倍性育种研究以来,对白桦四倍体及其子代的研究已有诸多报道,但这些研究还仅仅限于种子性状及1年生苗期生长等方面[23,15-17]。本研究对早期获得的三倍体家系于2010年在东北三省的4个地点营建子代测定林,3年生时调查其树高、地径以及保存率,进行了遗传变异分析。结果表明:参试的10个家系(包括6个白桦三倍体全同胞家系、4个二倍体半同胞家系)树高、地径及保存率在地点间、家系间的差异均达到显著或极显著水平。丹东试验点的树高、地径生长最好,家系树高均值为1.85 m,地径均值为3.05 cm。丹东造林点年均温较高,平均年降水量大,适宜白桦的生长发育。
有研究指出,多倍体植物其发育时间要较同种二倍体植物迟缓[24]。本试验由于参试对象为3年生的林分,尚处于发育初期,所以可能导致部分试验点的三倍体家系间树高、地径性状差异不显著。而位于辽宁省的2个试验点生长差异性显著,这与试验地点生长期长短及年积温等条件相关。地处辽宁省的2个试验点由于年均温较高、生长期长、年降雨量大的气候特点,白桦的生长量较大,6个三倍体家系间的生长特性表现较充分;而地处吉林省、黑龙江省的试验点由于生长期短、年积温较低,白桦生长量较小,所以亲缘关系较近的6个三倍体家系间的特性差异尚未充分表达。
虽然参试家系在高生长和径向生长上表现最优的均是二倍体家系,三倍体家系表现稍逊于二倍体家系,但各试验点表现最好的三倍体家系均与该试验点表现最好的二倍体家系差异未达到显著水平,这说明白桦三倍体家系的选择是具有潜力的。研究结果显示,各试验点同一母本不同父本的三倍体子代家系间存在较大差异,这说明在母本相同的情况下,子代杂种的差异是由二倍体父本不同决定的。这与研究团队前期试验结果一致,即白桦三倍体的生长既依赖于染色体加倍效应,又与亲本的配合力水平有关[15]。由于本试验仅有1株白桦四倍体母树(Q65)达到了开花结实的年龄,无法进行多个母本与多个父本的杂交选配,因此,单一母本的遗传基础限制,可能也是三倍体家系生长表现略逊于二倍体家系的原因。
实现林木优良家系在一定区域范围内的推广,开展生长稳定性的多点联合分析是重要环节之一。生产力高,产量稳定的家系是推广的首选家系。对参试家系分别采用生产力指数与基因型分组法分类,10个家系在2种区划方法中分布基本一致有3种家系,316家系划分的不一致,建议随着林龄的增长再对该家系进行研究划分。
将各试验点选择的三倍体优良家系与生长稳定性分类结果进行比较发现:各试验点的三倍体优良家系其稳定性均属于第Ⅱ类和第Ⅲ类中,例如,属于高产非稳产型的314和320家系,也在尚志、辉南、凌海试验点入选的优良家系中;属于低产稳产型的318和319家系,也在尚志、辉南、丹东试验点入选的优良家系中。目前,由于生产单位对于白桦这一树种利用方式的多样化,我们的选育目标也随之具有多个方向。对于高产非稳产型家系的选育主要侧重于其在生长适宜地区能够体现出产量增长迅速,成材所需时间短,木材产量高等方面的优势;而对于低产稳产型家系的选育则侧重于其生长适应能力强,可克服环境不良的影响,生长所需时间长则其木材材质势必高于速生类型,因此可以满足对材质要求较高的需求。我们建议对高产非稳产型的314和320家系在参试造林点及环境条件相近的毗邻地区进行推广。由于试验分析的林龄只有3年生,对营建的子代测定林的材积分析也是我们的后续研究。随着白桦制种园中四倍体母树的陆续开花结实,研究团队在后续的白桦四倍体育种中正在加大对亲本的选配,采用交配设计方法进一步深入白桦四倍体子代的遗传研究。
[1] 郭启高,宋明,梁国鲁.植物多倍体诱导育种研究进展[J].生物学通报,2000,35(2):8-10.
GUO Qigao,SONG Ming,LIANG Guolu.Plant induced polyploidy breeding research progress[J].Bull Biol,2000, 35(2):8-10.
[2] 孙敏红,张蜀宁.多倍体育种在园艺作物中的应用[J].江苏农业科学,2004,32(1):68-72.
SUN Minhong,ZHANG Shuning.Polyploid breeding in application of horticultural crops[J].Jiangsu Agric Sci,2004, 32(1):68-72.
[3] WEISGERBER H,RAU H M,GARTNER E J,et al.25 years of forest tree breeding in Hesse[J].Allg Forstztg,1980, 26:665-712.
[4] 康向阳.三倍体毛白杨新品种选育[J].北京林业大学学报,2004,26(3):40,63.
KANG Xiangyang.Triploid Populus tomentosa new species breeding[J].J Beijing For Univ,2004,26(3):40,63.
[5] 高克姝.三倍体毛白杨新品种对比试验[J].山西林业科技,1999(1):5-9.
GAO Keshu.Contrast test of triploid Populus tomentosa new species[J].J Shanxi For Sci Technol,1999(1):5-9.
[6] EINSPAHR D W.Production and utilization of triploid hybrid aspen[J].Iowa State J Res,1984,58(4):401-409.
[7] ZHANG Zhenghai,KANG Xiangyang,ZHANG Pingdong,et al.Incidence and molecular markers of 2n pollen in Populus tomentosa Carr.[J].Euphytica,2007,154(1/2):145-152.
[8] ZHANG Zhenghai,KANG Xiangyang.Cytological characteristics of numerically unreduced pollen production in Populus tomentosa Carr.[J].Euphytica,2010,173(2):151-159.
[9] 李云,冯大领.木本植物多倍体育种研究进展[J].植物学通报,2005,22(3):375-382.
LI Yun,FENG Daling.Advances in research into polyploidy breeding of woody plants[J].Chin Bull Bot,2005,22(3):375-382.
[10] 康向阳.林木多倍体育种研究进展[J].北京林业大学学报,2003,25(4):70-74.
KANG Xiangyang.Advances in researches on polyploid breeding of forest trees[J].J Beijing For Univ,2003,25(4):70-74.
[11] 赖杭桂,陈霞,徐洪伟,等.橡胶树三倍体种质创制及生物学鉴定[J].热带作物学报,2013,34(6):1001-1006.
LAI Hanggui,CHEN Xia,XU Hongwei,et al.Creation and biological identification of triploid germplasm in rubbertree[J].Chin J Trop Crop,2013,34(6):1001-1006.
[12] 尚宗燕,张继祖,刘谦虎,等.漆树染色体观察及三倍体漆树的发现[J].西北植物学报,1985,5(3):187-191.
SHANG Zongyan,ZHANG Jizu,LIU Qianhu,et al.The observation on chromosome of Rhus verniciflua stokes and discovery of triploid lacquer tree[J].Acta Bot Boreali-Occident Sin,1985,5(3):187-191.
[13] 杨今后,杨新华.桑树人工三倍体育种的研究[J].蚕业科学,1989,15(2):65-70.
YANG Jinhou,YANG Xinhua.Breeding of artificial triploid in mulberry[J].Ser Sin,1989,15(2):65-70.
[14] 刘福妹,穆怀志,刘子嘉,等.秋水仙素处理不同家系白桦种子诱导四倍体的研究[J].北京林业大学学报, 2013,35(3):87-92.
LIU Fumei,MU Huaizhi,LIU Zijia,et al.Inducing tetraploid of Betula platyphylla with different generations of seeds by colchicine[J].J Beijing For Univ,2013,35(3):87-92.
[15] 林琳,穆怀志,姚启超,等.白桦不同杂交组合三倍体子代当年生苗木生长性状分析[J].北京林业大学学报, 2012,34(5):1-5.
LIN Lin,MU Huaizhi,YAO Qichao,et al.Growth traits of triploid progenies in one year old from different crosses of Betula platyphylla Suk.[J].J Beijing For Univ,2012,34(5):1-5.
[16] 徐焕文,刘宇,李雅婧,等.白桦三倍体制种园中各家系种子活力比较[J].西南林业大学学报,2013,33(5):34-39.
XU Huanwen,LIU Yu,LI Yajing,et al.Comparative analysis on seed vitality of different families produced in Betula pendula triploid seed orchard[J].J Southwest For Univ,2013.33(5):34-39.
[17] 刘宇,徐焕文,姜静,等.基于种子活力及苗期生长性状的白桦四倍体半同胞家系初选[J].北京林业大学学报, 2014,36(2):74-80.
LIU Yu,XU Huanwen,JIANG Jing,et al.Family selection of birch tetraploid half-sibling based on seed vigor and seedling growth traits[J].J Beijing For Univ,2014,36(2):74-80.
[18] 杨传平,杨书文,刘桂丰,等.长白落叶松生长性状的稳定性分析[J].东北林业大学学报,1991,19(育种专刊):32-37.
YANG Chuanping,YANG Shuwen,LIU Guifeng,et al.The stability analysis of the growth character of Larix olgensis[J].J Northeast For Univ,1991,19(Spiss):32-37.
[19] 王秋玉,杨书文,刘桂丰,等.红皮云杉遗传稳定性的研究及最佳种源选择[J].东北林业大学学报,1993,21(1):5-12.
WANG Qiuyu,YANG Shuwen,LIU Guifeng,et al.Astudy on the genetic stability of Picea korraiesis and the optimal provenance selection[J].J Northeast For Univ,1993,21(1):5-12.
[20] 吴幼媚,王鹏良,韦颖文,等.桉树无性系区域试验[J].林业科技开发,2008,22(2):64-67.
WU Youmei,WANG Pengliang,WEI Yingwen,et al.Regional tests of clones of Eucalyptus[J].China For Sci Technol,2008,22(2):64-67.
[21] 李培,华劲松,王清,等.几种产量分析法在芸豆多点试验中的应用[J].西昌学院学报:自然科学版,2011,25(3):9-12.
LI Pei,HUA Jinsong,WANG Qing,et al.Application of several yield analytical methods on kidney bean’s multilocation test[J].J Xichang Coll Nat Sci Ed,2011,25(3):9-12.
[22] 刘桂丰,杨书文,杨传平,等.樟子松种源试验的研究——遗传稳定性测定及最佳种源选择[J].东北林业大学学报,1991,19(2):17-23.
LIU Guifeng,YANG Shuwen,YANG Chuanping.et al.Studies on the provenances test of mongolian scots pine:the genetic stability determination and the optimal provenance selection[J].J Northeast For Univ,1991,19(2):17-23.
[23] MU Hhuaizhi,JIANG Jing,LI Huiyu,et al.Seed vigor,photosynthesis and early growth of saplings of different triploid Betula families[J].Dendrobiology,2012,68(2):11-20.
[24] 杨俊宝,彭正松.多倍体植物的表观遗传现象[J].遗传,2005,27(2):335-342.
YANG Junbao,PENG Zhengsong.Epigenetic phenomena of plant polyploids[J].Hereditas,2005,27(2):335-342.
Growth performance and adaptability of Betula platyphylla offspring in the period of young forest
LIU Yu1,XU Huanwen1,LI Zhixin1,JIANG Jing1,PENG Rusheng2,SHANG Fuqiang3,ZOU Jianjun4, TENG Wenhua1,LIU Guifeng1
(1.State Key Laboratory of Tree Genetics and Breeding,Northeast Forestry University,Harbin 150040,Heilongjiang, China;2.Poplar Institute of Liaoning Province,Gaizhou 115200,Liaoning,China;3.Liaoning Institute of Forest Management,Dandong 118002,Liaoning,China;4.Jilin Provincial Academy of Forestry Science,Changchun 130033, Jilin,China)
Multi-point testing of tree progeny,an important link in the study of tree suitability and stability,was used to determine the height and ground diameter of Betula platyphylla (birch)for six triploid families and four diploid families planted at four sites:Shangzhi,Huinan,Linghai,and Dandong.3 years ago a multi-point conjoint analysis was used for differences in height between sites.Results of the multi-point conjoint analysis of height and ground diameter showed that differences in height between the sites were highly significant(P<0.01).Also,ground diameter for the Dandong and Linghai sites was significantly different(P<0.05).Families 314 and 319 were excellent triploid families at the Shangzhi site;at Huinan they were 318 and 320;at Linghai:314,316 and 320;and at Dandong:316,318 and 319.Then,tested families were divided into three types according to their productivity index and genotype grouping methods.The first type included high yieldinstability families 202,314 and 320.201,318 and 319 belonged to the low yield-stability type.The low yieldinstability type contained families 203,204 and 315.Excellent families selected from all sites belonged to thefirst and second type.This study not only provided a reference for early popularization of birch triploid families, but also formed an important basis for alterations of high generation triploid seed orchards.[Ch,3 fig.6 tab.24 ref.]
forest tree breeding;birch(Betula platyphylla)tetraploid progeny;multi-point test;family selection;genetic stability
S722.3
A
2095-0756(2015)06-0853-08
浙 江 农 林 大 学 学 报,2015,32(6):853-860
Journal of Zhejiang A&F University
10.11833/j.issn.2095-0756.2015.06.005
2015-01-23;
2015-04-01
“十二五”国家科技支撑计划项目(2012BAD01B05)
刘宇,从事林木遗传育种研究。E-mail:nefuly@126.com。通信作者:刘桂丰,教授,博士生导师,从事林木遗传育种研究。E-mail:liuguifeng@163.com
猜你喜欢
中国水产(2024年3期)2024-04-11 07:36:00
农业科技通讯(2023年1期)2023-02-12 07:09:08
科学养鱼(2021年11期)2022-01-11 11:21:10
食品与生活(2019年12期)2019-05-15 15:01:41
生命科学研究(2018年1期)2018-05-29 01:12:56
现代园艺(2017年13期)2018-01-19 02:28:17
江苏农业科学(2017年16期)2017-10-27 12:48:11
现代农业科技(2017年8期)2017-06-10 09:32:18
中成药(2016年4期)2016-05-17 06:07:54
中国果业信息(2015年6期)2015-01-23 17:38:07
