
多年封育对半干旱区草场植物多样性的影响
2015-01-03 06:30李俊琴
水土保持研究 2015年6期
李俊琴
(山西省水利水电勘测设计研究院,太原030024)
我国草原资源丰富,总面积达3.92亿hm2,但近年来随着一些人类不合理活动(如放牧、樵采、过度开垦、掠夺经营等)的加剧以及自然环境的恶化,使得我国草地生态系统退化严重,目前草地退化已成为我国生态环境恶化的突出问题之一。草地生态系统的退化是指其在演化过程中,其结构特征和能量流动等物质循环功能的恶化过程,即生物群落及其赖以生存环境的恶化过程[1]。由于不合理的放牧是引起草原退化的首要原因[2-3],我国实行了许多草地恢复的措施,其中对草地进行封育因其成本低、技术要求低和易推广的优势而得到广泛推广[4],这也是区域生态系统恢复和重建的一项重要举措,并已在世界各国得到广泛采用[5]。
人工封育是一种有效的快速植被恢复的措施,尤其是对北方农牧交错带这种降水量较少且年际变化较大的地方来说,这种效果尤为显著。围栏封育主要是通过人为的减少牲畜对草地生态系统的影响,使生态系统在自身的弹性下得以恢复和重建[6-9],它能够改善退化草场的群落组成,增加生态系统的物种多样性,并显著提高了植物生物量,同时对退化草场的结构和功能恢复起到一定的促进作用[10-12]。但封育时间和方式的不同,可能导致草地生态系统演替方向发生恶性变化,更有甚者可能造成草原的再次退化。本文以宁夏盐池县人工封育区为研究对象,通过研究多年封育过程中植物群落的多样性变化,探讨封育对草地植被群落和生物多样性的影响,并分析植被群落结构的变化,为制定合理的围栏封育措施提供依据,并为干旱、半干旱区封育措施下退化生态系统的植被恢复提供理论依据和技术支撑。
1 研究区概况
盐池县位于宁夏回族自治区东部长城沿线,北纬37°04′—38°10′、东经106°30′—107°41′,地处毛乌素沙漠南缘,黄土高原北部,地势南高北低,地形主要为剥蚀的准平原,境内地势南高北低,海拔1 295~1 951 m,属于典型的农牧交错地带。该县常年干旱少雨,风大沙多,年均气温为8.1℃,极端最高温和极端最低温为34.9℃和-24.2℃,年均无霜期为165d,年降水量仅为250~350mm,80%的降雨量集中在5—9月份,且由南向北、由东南向西北递减,但年蒸发量多达2 100mm,属于典型的中温带大陆性气候。土壤类型主要是灰钙土,此外还有黑垆土、风沙土、黄土、少量的盐土、白浆土等。植被在区系上属于欧亚草原区亚洲中部亚区—中部草原区的过渡带,是中国中部草原区的过渡地带。常见植物种类以旱生和中旱生类型为主,旱生类型主要分布于干草原区,中旱生类型则主要分布于草甸区。这些在气候、地理等自然条件方面的过渡性使得该县自然资源存在多样性和脆弱性等特点。
2 研究方法
2.1 外业调查
本试验结合当地荒漠化土地利用类型及主要的荒漠化治理工程种类,将研究区域设在位于毛乌素沙地西南缘,距盐池县城东北方向约20km处的人工封育区。该封育区自2002年以来用铁丝网作围栏对草场进行封育,以完全排除野生动物和家畜的采食。该试验在两个大样地中进行,样地相距较近,地理坐标为37°50′45″N,107°24′04″E,海拔1 366m,样地大小为100m×100m。本试验自2003年开始对样地内植物进行调查,调查时间选在每年的植物生长高峰期(7—8月份)进行,在每个样地内随机布设50个1m×1m样方,并对每个样方进行植物群落种类及各特征值的调查,调查包括植物名称、种数、株数、盖度、高度、地上生物量等。
2.2 数据处理方法
通过Excel 2010创建研究区草地植被数据库,得到并计算植物物种重要值矩阵,以及各个物种多样性指数。
2.2.1 重要值测度 不同学者对于重要值的表达有着不同见解,本文采用真正生态学意义上的重要值公式[13]:
重要值=(相对多度+相对频度+相对盖度+相对高度+相对生物量)/5
2.2.2 物种多样性测度 α多样性是评价群落内物种多样性的指标,一般将反映群落中物种丰富度和个体在各物种中分布均匀程度的指标称为a多样性[14],本研究选取下列α多样性指标来指示研究区内物种多样性性水平,并与环境因子做相关性分析。其计算公式为:
(1)物种丰富度计算:
S=样地或样方内出现的植物种数。
(2)Shannon-Wiener指数:

(3)Simpson多样性指数:

(4)群落均匀度指数:
E=H/lnS
式中:S——调查样方内植物种数;Ni——调查样方中第i种物种的重要值;N——调查样方内所有植物种的重要值和;Pi——第i个植物种的相对重要值;Pi=Ni/N;H——Shannon-Wiener指数。
3 结果与分析
3.1 物种重要值分析
通过用Excel 2010对研究区多年植被调查数据处理得出主要物种的重要值(表1),各年降雨量见图1。从表1可以看出自2003年开始至2014年只有3种植物每年都出现,分别是苦豆子(Sophoraalopecuroides)、黑 沙 蒿 (Artemisiaordosica)和 刺 沙 蓬(Salsolaruthenica),苦豆子和刺沙蓬的变化趋势基本一致。其中苦豆子刚开始时重要值非常低(0.065),随着封育时间的延长,它的重要值逐渐增加,在2006年达到最高(0.253),然后逐年降低,在2013年最低(0.037)。这是因为苦豆子根系比较发达,垂直根向下延深可达2~2.5m,且不定根无分层性,均匀扩散蔓延很广,其利用水分的能力比其他草本植物更强。由图1可知在刚开始的几年里,封育区内降水量较少,植物普遍生长状况不好,而苦豆子根系发达,利用水分能力强,所以重要值逐渐增长,但随着2007年降雨量增多,其它植物,特别是一年生草本植物的生长,这抑制了苦豆子的生长,所以其重要值也逐渐减少。而黑沙蒿重要值最大值出现在2003年,为0.371,是封育区内的优势物种,之后随着封育时间的延长其重要值逐渐降低,在2012年最低,为0.142,其优势种地位显著降低。其它物种的重要值也发生了显著的变化,许多植物甚至逐渐消失。而且从表1可以看出多年生草本重要值方面所占的比重随着封育时间的增长出现先减少后波动增加的趋势,而一年生草本则表现出先增长,然后维持相对稳定状态的变化规律。而半灌木所占的比例也逐年减低。这些都表明封育改变了研究区内原有的群落组成,使得原有优势种优势地位逐渐降低,其他物种的重要值波动变化,群落结构发生了明显的变化。
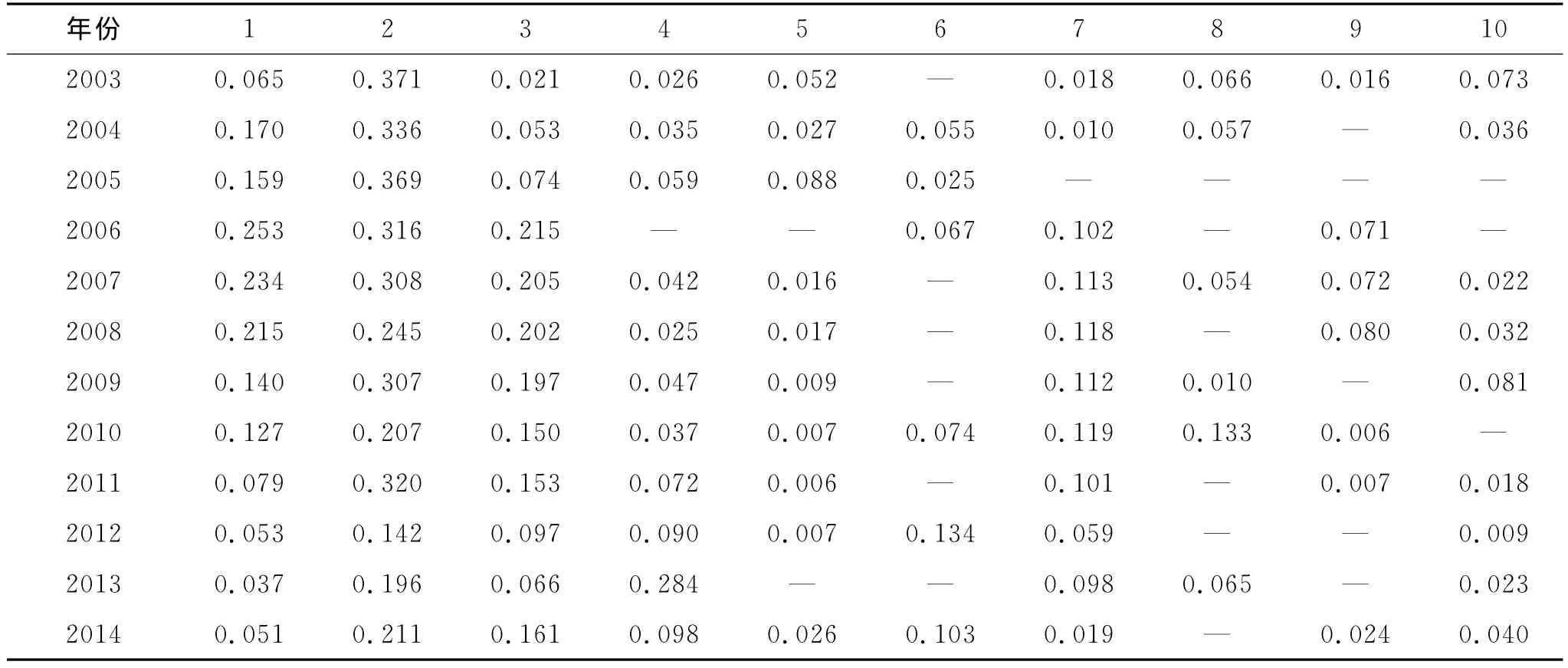
表1 各年主要植被重要值
3.2 物种多样性分析
由表2看出在封育刚开始时植被盖度和生物量都显著增高,随后又逐年波动降低。其中植被盖度在封育刚开始2~3a内快速生长,随后在封育4a时增长速度变慢,在封育第5年,也就是在2006年达到最高值,为67.3%,随后又逐渐波动降低,在封育9a时其值已低于封育三年时的盖度值,之后也是逐年减少,最终在封育第12年,即在2013年出现低峰值,为45.1%。而生物量的变化同植被盖度变化又略有不同,它在封育刚开始时逐渐增高,在封育第6年(即2007年)达到最高,为375.4kg/hm2,但随后又逐年逐渐波动降低,最终也是在封育第12年出现低峰值,为179.4kg/hm2。这些都说明封育在刚开始时确实有利于封育区内植物的恢复,但是随着封育的进行,植被恢复的速度逐渐减慢,并在封育的第5年左右出现植被开始逐步退化的现象,这说明封育时间与植被恢复并不是简单的线性关系,长时间的完全封育并不利于封育草场植被的恢复。表1中2011年和2014年植被盖度和生物量都比前一年有所提高,这主要是由于这两年降雨量(详情见图2)都比前一年有较大的提高导致的,并不能影响多年封育不利于草原植被恢复这一结论。

表2 不同封育年限植被盖度及生物量
不同年份植物各个多样性指数变化规律见图1,图2是自2003年开始盐池县降雨量变化。从图1中可以看出在封育刚开始的几年里植物的SW,SP多样性指数以及丰富度指数逐渐减少,这主要是由于随着围栏封育措施的实施,避免了牲畜对草场的破坏,并使人类活动逐渐减少,使封育区内个别物种得到大量的生长,如刺沙蓬和苦豆子,在刚开始封育的几年,它们的重要值也有了显著的提高(详情见表1),说明这两个物种在封育区内不断扩展,占据了大量的资源空间,这在一定程度上抑制了其他植物的生长,使得一些特化种生存空间变小直至消失,再加上这几年降雨较少,使得整个研究区植被生长状况普遍不好,这些综合因素导致各个多样性指数在这几年逐渐下降,丰富度指数也逐渐降低。而均匀度指数变化趋势则正好相反,因为许多特化种的消失,使得整个封育区内各个植物占有的资源空间更加均匀,植被的均匀度指数逐渐升高。之后随着封育时间的增长,许多优势种的生态地位逐渐减低,导致SW,SP多样性指数以及丰富度指数都比刚开始封育的几年有明显提高,封育区内重新出现了许多的特化种,如丝叶山苦荬(Ixerischinensis)和草木樨状黄芪(Astragalusmelilotoides),见表1。随着封育的继续进行,许多优势种的生态地位逐渐减低,各个多样性指数呈现周期性变化,由图1可以看出低峰和高峰出现的频率均约为4a/次,尽管个别年限(如2011年)还会出现一定的波动,但这也主要是由于降雨量等天气因素引起的。因此认为长期的完全封育会使植物的各个多样性指数发生周期性的波动,这并不利于草场植被的恢复以及管理,所以改善封育政策势在必行。

图1 不同年份植物多样性指数变化规律

图2 不同封育年限降雨量变化
4 结论与讨论
许多研究表明草场恢复会使得群落物种组成发生重大变化,并且导致物种多样性发生改变[15-17],本研究结果发现,多年生草本、一年生草本以及半灌木重要值比例随着封育的进行逐渐发生变化,其中半灌木所占比例越来越小,多年生草本所占比重随着封育时间的增长表现出先减少后波动增加的趋势,而一年生草本则表现出先增长,然后维持相对稳定状态的规律。这些都表明封育改变了封育区内原有的群落组成,使得原有优势种(黑沙蒿)地位逐渐降低,群落结构发生了明显的变化。
本研究结果显示,封育在刚开始时确实有利于封育区内植被的恢复,但是随着封育的进行,植被恢复的速度逐渐减慢。这是因为在围封条件下刚开始的几年土壤含水量得到大幅度提高[18-19],这使土壤存在的大量植物种子具备了萌发的条件,植被得到大量恢复,但是随着封育时间的加长,在近地表形成生物结皮,并出现土壤近地表板结的现象,降水渗透率和利用率减小,植物群落随着封育年限的增加呈现衰退趋势[20]。在封育5a后封育区内植被盖度和生物量都逐渐降低,说明植被出现退化的现象,这表明封育时间与植被恢复并不是简单的线性关系,长时间的完全封育并不利于封育草场植被的恢复,合理的封育也应该是在这时对封育区进行适当的人为干预,如进行刈割或者季节性的放牧等,这可以使草地生态系统的能量流动和物质循环保持良性状态,进而保持草地生态系统平衡,将更加有利于封育区内的植被恢复。
在群落生态学中物种多样性指数、丰富度指数和均匀度指数一直都是研究的重点内容,不同学者对围封与三者之间相互影响关系研究得出的结论并不相同[21-23]。本研究显示,封育时间与植被恢复并不是简单的线性关系,物种多样性随封育时间处于一种周期性波动状态,这和蒋德明等[24]通过对科尔沁沙地封育措施下植被以及土壤特性的研究得出的结论一致,并且这种波动也受降水量影响,当降水量发生变化时,各个多样性指数也会发生相应的变化,但这种影响效应在时间上具有滞后性[25]。同时本文通过对比各个年限植物多样性指数变化规律,得出封育区SW,SP多样性指数随封育年限波动规律与物种丰富度的变化基本一致,都表现为封育刚开始的几年逐渐降低,随着封育时间的增长,会比刚开始封育的几年有较大的提高,之后呈现一定的规律性波动,低峰和高峰出现的频率均约为4a/次。尽管个别年限会有一定的偏差,但这也主要是由于降雨量等天气因素的变化引起的,并不能影响这一结论,因此可认为物种丰富度能够在一定程度上代表物种多样性指标。
[1] 李博.中国北方草地退化及其防治对策[J].中国农业科学,1997,30(6):1-9.
[2] 中国科学院内蒙古宁夏自然资源综合考察队.内蒙古植被[M].北京:科学出版社,1985.
[3] 中国植被编辑委员会.中国植被[M].北京:科学出版社,1980:1080-1094.
[4] 高凯,朱铁霞,韩国栋.围封年限对内蒙古羊草—针茅典型草原植物功能群及其多样性的影响[J].草业学报,2013,22(6):39-45.
[5] 闫玉春,唐海萍,辛晓平,等.围封对草地的影响研究进展[J].生态学报,2009,29(9):5039-5046.
[6] Middleton N J,Thomas D S G.Worid Atlas of Desertification[M].2nd edition London:Edward Amold,1998.
[7] Meissner R A,Facelli J M.Effects of sheep exclusion on the soil seed bank and annual vegetation in chenopods shrublands of South Australia[J].Journal of Arid Environments,1999,42:117-128.
[8] 李永宏.内蒙古典型草原地带退化草原的恢复动态[J].生物多样性,1995,3((3):125-130.
[9] 杨晓晖,张克斌,侯瑞萍.封育措施对半干旱草场植被群落特征及地上生物量的影响[J].生态环境,2005,14(5):730-734.
[10] 陈四清,崔骁勇,周广胜,等.内蒙古锡林河流域大针茅草原土壤呼吸和凋落物分解的CO2排放速率研究[J].植物学报,1999,41(6):645-650.
[11] 左万庆,王玉辉,王风玉,等.围栏封育措施对退化羊草草原植物群落特征影响研究[J].草业学报,2009,18(3):12-19.
[12] 邹雨坤,张静妮,杨殿林,等.不同利用方式下羊草草原土壤生态系统微生物群落结构的PLFA分析[J].草业学报,2011,20(4):27-33.
[13] 王育松,上官铁梁.关于重要值计算方法的若干问题[J].山西大学学报:自然科学版,2010,33(2):312-316.
[14] 杨持.生态学试验与实习[M].北京:高等教育出版社,2003.
[15] 赵忠,王安禄,马海生,等.青藏高原东缘草地生态系统动态定位监测与可持续发展要素研究:Ⅱ.高寒草甸草地生态系统植物群落结构特征及物种多样性分析[J].草业科学,2002,19(6):9-13.
[16] 董全民,马玉寿,李青云,等.牦牛放牧率对小嵩草高寒草甸暖季草场植物群落组成和植物多样性的影响[J].西北植物学报,2005,25((1):94-102.
[17] 程中秋,张克斌,刘建,等.宁夏盐池荒漠草原区天然草地植物生态位研究[J].水土保持研究,2011,18(3):36-40.
[18] 马建军,姚宏,冯朝阳,等内蒙古典型草原区3种不同草地利用模式下植物功能群及多样性的变化[J].植物生态学报,2012,36(1):1-9.
[19] 单贵莲,初晓辉,田青松,等.典型草原恢复演替过程中土壤形状动态变化研究[J].草业学报,2012,21(4):1-9.
[20] 张克斌,卢晓杰,李瑞.北方农牧交错带沙地生物结皮研究[J].干旱区资源与环境,2008,(4):147-151.
[21] Meissner R A,Facelli J M.Effect of sheep exclusion on the soil seed bank and annual vegetation in chenopod shrub lands of South Australia [J].Journal of Arid Environment,1999,42:117-128.
[22] 周华坤,周立,赵新全,等.围栏封育对轻牧和重牧金露梅灌丛的影响[J].草地学报,2007,12(2):140-144.
[23] 刘建,张克斌,程中秋,等.围栏封育对沙化草地植被及土壤特性的影响[J].水土保持研究,2011,31(4):180-184.
[24] 蒋德明,苗仁辉,押田敏雄,等.封育对科尔沁沙地植被恢复和土壤特性的影响[J].生态环境学报,2013,22(1):40-46.
[25] 刘小丹,李瑞,张克斌,等.半干旱区人工封育草场植物群落物种多样性与复杂性研究:以宁夏盐池为例[J].生态环境学报,2014,23(7):1093-1101.
猜你喜欢
天津农林科技(2022年2期)2022-04-19
河南畜牧兽医(2022年3期)2022-04-13
昆明医科大学学报(2022年2期)2022-03-29
食品安全导刊(2021年20期)2021-08-30
绿色科技(2021年10期)2021-06-23
黑龙江工程学院学报(2020年5期)2020-10-21
河南科学(2020年3期)2020-06-02
水土保持研究(2019年6期)2019-10-19
草业学报(2019年2期)2019-02-25
草业科学(2015年9期)2015-12-11
