
麦秀林区青海云杉种群发育特征及其格局研究
2015-01-02 10:19高发明杨启青才让丹周周卫芬仁青多杰周国荣韩富忠田呈明
西北林学院学报 2015年4期
高发明,杨启青,才让丹周,周卫芬,仁青多杰,周国荣,韩富忠,田呈明*
(1.北京林业大学 林学院,北京100083;2.青海省林业厅 森林病虫害防治检疫站,青海 西宁810000;3.青海省黄南州麦秀林场,青海 黄南811300)
生物群落内各物种在空间和时间上的配置和分布所形成的种群分布格局,不仅依赖种群自身的生物学和生态学特征,并在很大程度上受群落种类组成和结构以及生境条件变化的综合影响[1-2],也是物种间生物学特性、种群之间、种群与环境之间相互作用的结果[3-4]。
云杉林作为青藏高原东部高山、亚高山地带的主要森林植被类型之一,具有独特的森林景观[5],并对稳定区域森林生态系统、遏制草地扩展与生态退化方面有极其重要的作用,被喻为三江源区生态屏障中的关键“守护者”[6]。目前国内学者对青海云杉林的群落特征、种群结构与动态等进行了研究[7-12],但对三江源地区的青海云杉,尤其是种群发育特征和分布格局方面尚未见报道。因此,开展青海云杉种群发育特征与分布格局研究,分析影响其种群发育与格局的重要因素或过程,对天然云杉林寄生害的有效防控,云杉天然林的保育、恢复与经营决策等具有重要价值。
1 材料与方法
1.1 研究区概况
麦秀林区的地理坐标为101°46′-102°04′E,35°8′-35°21′N,属高山峡谷地貌,地形呈南高北低走势。气候属于温带半干旱区域,境内年平均气温2.8℃,极端最高气温29℃,极端最低气温-31℃,气温极差60℃;年降水量477.4~511.9mm,6-9月占全年降水量的70.9%,无绝对无霜期,风大,空气干燥,年蒸发量远大于年降水量[13]。植被总的表现为青藏高原类型,森林类型主要以青海云杉纯林、紫果云杉纯林、云杉与山杨及桦木混交林、云杉与祁连圆柏混交林为主,灌木以高山柳、杜鹃、忍冬、锦鸡儿等为主,林下植被主要以菊科、莎草科、禾本科、蓼科、十字花科和唇形科植物为主[14]。
1.2 样方设置
2012年5-7月,在青海省黄南州麦秀林区,对分布于阴坡、半阴坡青海云杉种群进行实地调查,调查中样方面积和数量的确定参照周红敏[15]等的方法。按照云杉种群分布区的海拔等间隔(150m)划分为低、中、高3个海拔高度范围,每个海拔范围设置为30m×30m的调查样方6个,共计18个,分别编号为1、2、3、…、17、18,对乔木植物进行每木调查,具体包括胸径、树高等;同时记录各样方的海拔、坡度、坡向、坡位等生境指标(表1)。

表1 青海云杉种群的样方概况Table 1 Survey of sample plot of Picea crassifolia population
1.3 不同生长发育阶段的划分
利用种群年龄结构分析种群动态,采用刘建泉[7]等的方法,树木胸径以2cm为间距的径阶划分,将青海云杉种群划分为4个年龄段:Ⅰ幼苗、幼树阶段,年龄1~15a,树高≤1.5m;Ⅱ小树阶段,年龄16~50a,胸径2~10cm;Ⅲ中树阶段,年龄51~100a,胸径12~20cm;Ⅳ大树阶段,年龄>100a,胸径≥22cm。在样方内,统计各龄期青海云杉的株数,并分别对青海云杉种群的4个发育阶段进行空间分布格局类型检验,编制青海云杉静态生命表及存活曲线图,定量描述青海云杉种群动态[16]。

式中,Vn表示种群从n到n+1级的个体数量变化动态,Vpi表示整个种群结构的数量变化动态指数,式中Sn与Sn+1分别为第n与第n+1年龄级种群个体数,max(…)示取括符中数列级大值,-1≤Vn≤1,Vn、Vpi分别取正、负、零值时,可以反映出种群相邻年龄级间个体数量增长、衰退、稳定的结构动态关系。
当考虑未来的外部干扰时:

式中,min(…)表示取括号中的数列最小值。K为种群年龄级数量。
1.4 数据处理
分布格局类型常采用方差/均值比率法、负二项式法进行判别,聚集强度采用负二项式参数(K)、平均拥挤度(m*)、聚块性指数(m*/m)和Green指数(GI)[17]进行测定。
1)方差均值比(扩散系数DI)

式中,S2表示样本的方差,表示样本的平均数。若DI=l时,种群属随机分布;若DI>1时,为集群分布;若DI<1时,为均匀分布。采用t检验来确定S2/的实测值与1的差异程度,比较t与t0.05(n-1)确定其差异显著性。

式中,n表示抽样单位总数。
Morisita指数:I=1为随机分布;I<1为均匀分布;I>1为集群分布。
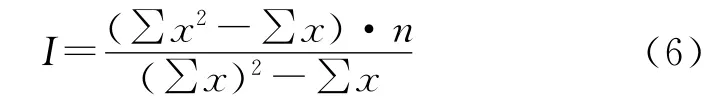
该方法的显著性检验可用χ2检验:
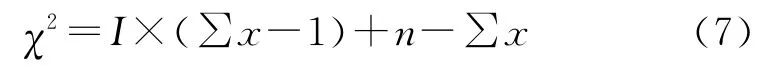
比较χ2与χ2(n-1)确定其差异显著性。
2)负二项参数K
K是聚集程度的测度参数,其计算公式

K值在最大聚集程度时趋向于0,K值越小,聚集度越大,如果K值趋于无穷大(一般为8以上),则逼近随机分布。
3)平均拥挤度(m*)与聚块性指数(m*/m)

式中,m*表示平均拥挤度,m表示总体平均值。
m*>1为聚集分布;m*=1为随机分布;m*<1为均匀分布。
聚块性指数m*/m:当m*/m>1时,为聚集分布;m*/m=1时,为随机分布;m*/m<1时,为均匀分布。
4)Green指数(GI)

GI>0为集群分布,GI<0为均匀分布。
2 结果与分析
2.1 青海云杉种群静态生命表及存活曲线
长生命周期的多年生植物可应用静态生命表来分析种群的动态[18]。青海云杉种群的静态生命表反映出其种群中幼苗、幼树个体数量最少,仅占6.6%;小树阶段个体较丰富,其他阶段种群结构较为合理(表2)。云杉种群内幼苗、幼树阶段个体的死亡率最高,导致幼龄数量严重不足,尽管小树存活率较高,但小树阶段之后死亡率升高,存活曲线趋向于Deevey-ⅳ型,其种群结构存在很大的波动性(图1)。

表2 青海云杉静态生命表Table 2 Static life table of P.crassifolia

图1 青海云杉种群存活曲线Fig.1 The survivorship curve of P.crassifolia population in Maixiu Forest Farm
青海云杉种群年龄结构的数量变化动态分析显示相邻各龄级间个体数量表现为:V1=-75.9%,V2=42.3%,V3=41.5%,整个种群年龄结构的动态指数Vpi=26.4%;当考虑到外部干扰对种群动态的影响时,Vpi′=4×10-4>0。因此,麦秀林区青海云杉种群在短时期内表现为稳定型种群,但随着时间的推移其动态演化趋势已经显示该种群处于衰退的早期阶段。
2.2 青海云杉种群不同发育阶段的分布格局
不同发育阶段的青海云杉种群的空间分布格局呈现出相似性(表3)。负二项式法、t检验和Morisita指数I的χ2检验均表明麦秀林区的青海云杉在不同的发育阶段,其种群分布呈集群分布。而不同发育阶段的Green指数GI和平均拥挤度m*分析反映出青海云杉种群整体上趋于集群分布,但各龄级的聚集程度不尽相同。从平均拥挤度m*来看,小树阶段极显著地高于幼苗幼树、中树以及大树阶段(p<0.01),其他各发育阶段之间没有显著性差异(p>0.05)。同时青海云杉不同发育阶段中,除幼苗、幼树外,聚块性指数均呈现小树>中树>大树的趋势,表明随着径级的增大或年龄的增加,整体上呈扩散的趋势。

表3 青海云杉不同发育阶段的分布格局Table 3 Spatial pattern of P.crassifoliaat different developmental stages

表4 青海云杉种群不同海拔高度的分布格局Table 4 Spatial pattern of P.crassifoliain different altitudes
2.3 不同海拔高度青海云杉种群的分布格局
对扩散系数DI的t检验和Morisita指数I的χ2检验结果均表明,在低、中、高海拔青海云杉种群均为聚集型分布格局。Green指数、平均拥挤度m*、聚块性指数(m*/m)分析显示不同海拔高度的青海云杉种群均趋于集群分布,但其聚集程度不尽相同,呈现中海拔>低海拔>高海拔的趋势。不同海拔高度青海云杉种群分布格局用负二项式法判别时,除了高海拔属于随机分布格局外,中、低海拔均属于聚集分布,而用方差均值比率法判别时,结果是一致的,均属于聚集分布。
3 结论与讨论
种子和幼苗期被认为是植物种群生存与发展过程中最脆弱的阶段[19-20]。麦秀林区青海云杉目前属于稳定型种群,但种群幼苗、幼树阶段所占比例低,其个体数量远低于其他3个发育阶段,从种群动态发展的角度来看,该种群已处于衰退的早期阶段。
青海云杉种群不同发育阶段均呈集群分布,但聚集程度有所不同,可能与生境空间异质性及不同程度的干扰有关。由样方资料可知青海云杉幼苗、幼树一般都出现在林窗附近,在较小规模上成聚集分布。同时麦秀林区云杉林受自然灾害(干旱、雪压、风倒风折、滑坡等),林业有害生物(云杉矮槲寄生、小蠹虫等)[21]的危害,人为干扰(放牧、采摘球果、森林砍伐)等影响,从而导致幼苗、幼树出现高的死亡率。但作者在调查期间也发现林间放牧活动已经常态化,牲畜频繁地踩踏导致林地裸露与土壤侵蚀严重,形成了植物无法定居生长的空白区,使得种群的更新极度困难。麦秀林区虽地处三江源自然保护区核心区,但又属林牧交错地区,国有林区保护措施与当地牧民放牧活动矛盾突出[22],该林区青海云杉幼苗、幼树数量严重不足与当地放牧干扰严重有关。刘建泉[23]等认为不同封育季节放牧干扰(尤其是冬季放牧)对哈溪林区青海云杉种群结构和动态的影响很大,是导致幼苗幼树的储备不足,种群结构失调的主要原因。
环境是植物生存和发展的条件,海拔梯度由于包含了温度、湿度、光照等诸多环境因子,必然会影响植物种群的分布格局[24-25]。不同海拔高度的青海云杉种群均为聚集型分布格局,但聚集程度不尽相同。对于出现此种格局的原因,一方面与青海云杉自身生物学特性有关,另一方面也与麦秀林区冬长夏短,冬季受西风带控制时间长,气候干燥、寒冷等气候条件的影响[14]有关。集群分布被认为是在各种森林生态系统中植物种群最普遍的分布类型,一定程度的群聚是树木生长、存活以及竞争等适应环境异质性的结果[26-27]。针叶林树木通常在亚高山地带多呈随机分布或聚集分布[28]。因此,麦秀林区青海云杉种群在不同海拔高度上的集群分布,是适应青藏高原各种严酷生境条件的一种生态策略。
[1] 孙儒泳,李博,诸葛阳,等.普通生态学[M].北京:高等教育出版社,1993:149-160.
[2] 彭少麟.南亚热带森林群落动态学[M].北京:科学出版社,1996:20-101.
[3] GREIG-SMITH P.Quantitative plant ecology [M].3rd ed.Oxford:Blackwell,1983:105-145.
[4] 杨心兵,覃逸明,刘胜祥.武汉市马鞍山森林公园马尾松年龄种群结构与分布格局[J].华中师范大学学报:自然科学版,2001,35(2):209-213.YANG X B,TAN Y M,LIU S X.Age structure and distribution pattern of Pinus massonianapopulation in Wuhan Ma’an Mountain Forest Park[J].Journal of Central China Normal University:Nature Science,2001,35(2):209-213.(in Chinese)
[5] 管中天.森林生态研究与应用[M].成都:四川科学技术出版社,2005:521-543.
[6] 刘鑫,包维楷.青藏高原东部近林线紫果云杉原始林的群落结构与物种组成[J].生物多样性,2011,19(1):34-40.LIU X,BAO W K.Community structure and vascular plant species composition of primary spruce forest near timberline in the eastern Tibetan Plateau[J].Biodiversity Science,2011,19(1):34-40.(in Chinese)
[7] 刘建泉,丁国民,郝虎,等.青海云杉群落特征和动态研究[J].西北林学院学报,2008,23(1):14-17.LIU J Q,DING G M,HAO H,et al.Structure and dynamic of Picea crassifoliacommunity in Qilian Mountains forest area[J].Journal of Northwest Forestry University,2008,23(1):14-17.(in Chinese)
[8] 刘建泉,屈永才,王学福,等.青海云杉群落空间结构特征研究[J].干旱区资源与环境,2009,23(7):221-227.LIU J Q,QU Y C,WANG X F,et al.Structure characteristics of Picea crassifolia communities in Qilian Mountains forest area [J].Journal of Arid Land Resources and environment,2009,23(7):121-125.(in Chinese)
[9] 刘建泉.祁连山保护区青海云杉种群分布格局的研究[J].西北林学院学报,2004,19(2):152-155.LIU J Q.Spatial pattern of Picea crassifolia population in Qilianshan Nature Conservation [J].Journal of Northwest Forestry University,2004,19(2):152-155.(in Chinese)
[10] 郝虎,刘兴明,宋采福,等.祁连山北坡苔藓—青海云杉林优势种空间分布格局研究[J].甘肃林业科技,2005,30(4):9-12.
[11] 刘建泉,李成龙,杨建红,等.青海云杉群落优势种群空间格局多样性研究[J].现代农业科技,2010,8:216-218.
[12] 刘建泉,孙建忠,杨开恩,等.东大河林区青海云杉种群结构与动态[J].西北林学院学报,2012,27(2):18-21.LIU J Q,SUN J Z,YANG K E,et al.Structure and dynamics of Picea crassifolia population in Dongdahe forest area of Qilian Mountains Nature Reserve [J].Journal of Northwest Forestry University,2012,27(2):18-21.(in Chinese)
[13] 王国仓,周华卿.麦秀林区针叶林土壤分类初探[J].青海农林科技,2007(2):64-66.
[14] 姜登伟.麦秀林区植物简志[M].西宁:青海人民出版社,1999:1-3.
[15] 周红敏,惠刚盈,赵中华,等.森林结构调查中最适样方面积和数量的研究[J].林业科学研究,2009,22(4):482-495.ZHOU H M,HUI G Y,ZHAO Z H,et al.Studies on the area and the number of the sample for forest structure[J].Forest Research,2009,22(4):482-495.(in Chinese)
[16] 陈晓德.植物种群与群落结构动态量化分析方法研究[J].生态学报,1998,18(2):214-217.CHENG X D.A study on the method of quantitative analysis for plant population and community structural dynamics[J].Acta Ecologica Sinica,1998,18(2):214-217.(in Chinese)
[17] 徐汝梅,成新跃.昆虫种群生态学—基础与前沿[M].北京:科学出版社,2005.
[18] SILVERTOWN J W.Introduction to plant population ecology[M].London:Longman Press,1982.
[19] HARPER J L.Populotion biology of plants[M].London:Academic Press,1977.
[20] ALEXANDER H M,WULFF R D.Experimental ecological genetics in Plantago[J].The Journal of Ecology,1985,73:271-282
[21] 夏博.云杉矮槲寄生对天然云杉林的影响及成灾因子[D].北京:北京林业大学,2011.
[22] 王谦,王军强.国有林区开展群众管护经营改革的探索—青海省试点调研[J].林业经济,2010(4):64-68.
[23] 刘建泉,罗永寿,吕海元.不同封育季节放牧干扰对青海云杉种群结构和动态的影响[J].草业学报,2012,29(6):983-988.LIU J Q,LUO Y T,LYV H Y.Structure and dynamics of Picea crassifolia populations with graze disturbance in different seasons of enclosed forest [J].Pratacultural Science,2012,29(6):983-988.(in Chinese)
[24] LINGUA E,CHERUBINI P,MOTTA R,et al.Spatial structure along an altitudinal gradient in the Italian central Alps suggests competition and facilitation among coniferous species[J].Journal of Vegetation Science,2008,19:425-436.
[25] 潘红丽,李迈和,蔡小虎,等.海拔梯度上的植物生长与生理生态特性[J].生态环境学报,2009,18(2):722-730.PAN H L,LI M H,CAI X H,et al.Responses of growth and ecophysiology of plants to altitude[J].Ecology and Environmental Sciences,2009,18(2):722-730.(in Chinese)
[26] MANABE T,NISHIMURA N,MIURA M,et al.Population structure and spatial patterns for trees in a temperate old growth evergreen broad leaved forest in Japan[J].Plant E-cology,2000,151:181-197.
[27] ARÉVALO J R,FERNÁNDEZ-PALACIOS J M.Spatial patterns of trees and juveniles in a laurel forest of Tenerife,Canary Islands[J].Plant Ecology,2003,165:1-10.
[28] HOLEKSA J,SANIGA M,SZWAGRZYK J,et al.Altitudinal variability of stand structure and regeneration in the subalpine spruce forests of the Pol’ana biosphere reserve,Central Slovakia[J].European Journal of Forest Research,2007,126:303-313.
猜你喜欢
作物学报(2022年7期)2022-05-11
现代农村科技(2020年7期)2020-12-19
河北果树(2020年1期)2020-02-09
花卉(2020年6期)2020-01-07
乡村科技(2019年14期)2019-02-21
启迪与智慧·教育版(2018年12期)2018-02-26
科学与财富(2017年26期)2017-09-23
现代园艺(2017年11期)2017-06-28
土壤与作物(2015年3期)2015-12-08
江苏农业科学(2014年9期)2014-11-15
