
低温胁迫对4种幼苗的叶绿素荧光特性的影响
2014-12-27 09:24卢广超许建新吴彩琼邵怡若
中南林业科技大学学报 2014年2期
卢广超 ,许建新 ,,薛 立 ,张 柔 ,吴彩琼 ,邵怡若
(1.华南农业大学 林学院,广东 广州510642;2.深圳市铁汉生态环境股份有限公司,广东 深圳518040)
低温胁迫对4种幼苗的叶绿素荧光特性的影响
卢广超1,许建新1,2,薛 立1,张 柔1,吴彩琼2,邵怡若1
(1.华南农业大学 林学院,广东 广州510642;2.深圳市铁汉生态环境股份有限公司,广东 深圳518040)
采用人工气候箱的方法,测定了低温胁迫下葛藤Pueraria lobata、假连翘Duranta repens、老鸭嘴Thunbergia erecta和台湾相思Acacia confuse幼苗的荧光参数,以便为筛选抗寒种类提供依据。结果表明,随着低温胁迫时间的延长,4种幼苗的最大荧光(Fm)、表观电子传递速率(ETR)、PSII实际光量子效率Y(II)逐渐下降,非光化学淬灭(NPQ)先上升后下降,PSII调节性能量耗散Y(NPQ) 和PSII非调节性能量耗散Y(NO)逐渐升高。经过24 h自然恢复,4种幼苗的Fm均恢复到对照水平,ETR有所回升,Y(NPQ)恢复到显著高于对照水平;葛藤、老鸭嘴和台湾相思的NPQ保持稳定,假连翘的NPQ有所下降;假连翘和台湾相思的Y(II)恢复到对照水平,葛藤、假连翘和台湾相思幼苗的Y(NO)恢复到对照水平。对荧光参数进行主成分分析表明,干旱胁迫下4种幼苗的抗寒顺序为台湾相思>假连翘>葛藤>老鸭嘴。
低温胁迫;幼苗;叶绿素荧光参数;主成分分析
低温伤害是一种常见的自然灾害,也是植物区域性分布和季节性生长的限制因素[1]。低温冷害会导致植物的光合酶活性、光合电子传递速率下降[2],从而削弱植物利用光能的能力[3],抑制叶黄素循环参与的非光化能量耗散或抑制蛋白修复循环[3],引起光抑制现象[4]。一定胁迫范围内,植物通过热耗散、PSII 反应中心可逆失活和Mehler反应等过程保护光合机构免受破坏[5]。低温胁迫严重时,会抑制PSII的修复,甚至对光合机构产生不可逆的破坏[2]。
目前,关于低温胁迫对叶绿素荧光影响的研究主要集中于蔬菜水果和禾本植物与少数南方树种。王贺等[6]发现低温使苹果最大光化学量子产量(Fv/Fm)降低,并对叶片的光合系统造成了严重破坏。马博英等[7]通过研究植物的PSII实际光量子效率(Y(II))、光合电子传递速率(ETR)和非光化学猝灭系数(NPQ)等叶绿素荧光参数,筛选出抗寒能力强的基因型草坪草。陈曦等[8]通过研究低温下油松叶绿素荧光发现,0℃以下的低温胁迫对油松造成不可恢复的光合机构损伤。梁芳等[9]发现菊花的Y(II)、ETR和Fv/Fm均随着低温胁迫程度的增加而降低。葛藤Pueraria lobata、假连翘Duranta repens、老鸭嘴Thunbergia erecta和台湾相思Acacia confuse是热带、亚热带地区的重要用材和园林绿化树种。尽管有学者报道了有低温胁迫下葛藤、假连翘和老鸭嘴的生理和光合特性[10],干旱胁迫下葛藤、假连翘和老鸭嘴的光合和荧光特性[11],干旱胁迫下葛藤的生理和荧光特性[12],中水水培下假连翘的光合特性[13],干旱胁迫和复水后台湾相思生理变化[14],尚未见到以上4种植物低温胁迫下荧光特性的报道。作者对该内容进行研究,可以从植物的荧光特性角度为筛选抗寒种类提供参考。
1 材料与方法
1.1 试验材料
试验于2011年9月在华南农业大学林学院进行。试验所用幼苗葛藤、假连翘、老鸭嘴和台湾相思均为营养袋实生苗, 营养袋直径12 cm,高10 cm。幼苗生长基本情况见表1。
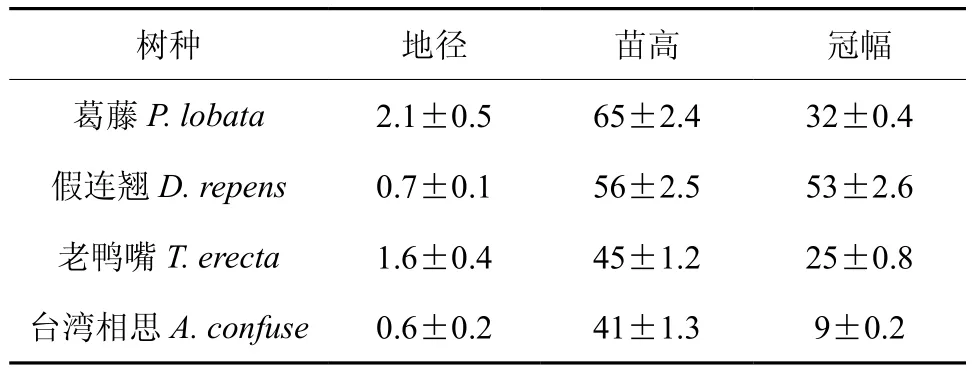
表1 幼苗生长基本情况(cm) (平均值±标准差)Table 1 General situation of experimental seedling types(cm) (mean±SD)
1.2 材料处理
每种幼苗选形态长势基本一致,生长状况良好的5株植株放入RXZ智能型人工气候箱进行6℃低温胁迫处理。光照时间8:00~17:00,光照强度为 120 mmol(photons)·m-2s-1,相对湿度为80%~85%。分别在连续低温处理后的第0 h、24 h、36 h和低温处理36 h,后在自然条件下恢复生长24 h (R24),选用幼苗顶部开始的第3~8 枚叶中的五枚成熟叶进行挂牌标记,在光合作用比较活跃的9:00~11:00测定叶绿素荧光参数。每叶片记录3个值,取平均值。
1.3 测定指标
叶片叶绿素荧光参数测定采用脉冲调制荧光仪OS-1P调制荧光仪将测定测试叶片在暗适应20 min后,在非化学淬灭(NPQ)模式下设定每次测量时间为1分钟,每次测量3次重复,测定内容有光下初始荧光(Fo)、最大荧光(Fm)、任意时间实际荧光(F′)、光适应下最大荧光(Fm′)。计算表观电子传递速率(ETR=Y(II)×PAR×0.84×0.5,PAR为光合有效幅射),非光化学淬灭(NPQ=Fm/Fm′-1),PSII实际光化学效率(Y(II)=(Fm′-F′)/Fm′)、PSII调节性能量耗散比例(Y(NPQ)=F′/Fm′-F′/Fm),PSII非调节性能量耗散比例(Y(NO) =F′/Fm)。
1.4 数据处理软件
运用Microsoft Excel 2003软件对数据进行统计分析和作图,运用SAS软件进行Duncun多重比较和主成分分析。
2 结果与分析
2.1 低温胁迫下幼苗的最大荧光变化
随着低温胁迫时间的延长,葛藤、假连翘和老鸭嘴幼苗的最大荧光(Fm)逐渐下降,在36 h时Fm降到最低值,台湾相思在低温胁迫24 h时Fm达到最低值,36 h有所回升(图1)。葛藤、假连翘、老鸭嘴和台湾相思幼苗在低温胁迫36 h后Fm分别为对照的70%、52%、82%和83%,均显著小于对照(P<0.05)。经过24 h的自然恢复,4种幼苗的Fm均恢复到对照水平。
2.2 低温胁迫下幼苗的表观电子传递速率变化
低温胁迫处理期间,4种幼苗表观电子传递速率(ETR)逐渐下降,在低温处理12、24、36 h时均显著小于对照(P<0.05),36 h分别为对照的38%、50%、49%和59%(图2)。经过24 h自然恢复,假连翘的ETR恢复到对照水平,其余3种幼苗的ETR有所回升,仍显著小于对照(P<0.05)。
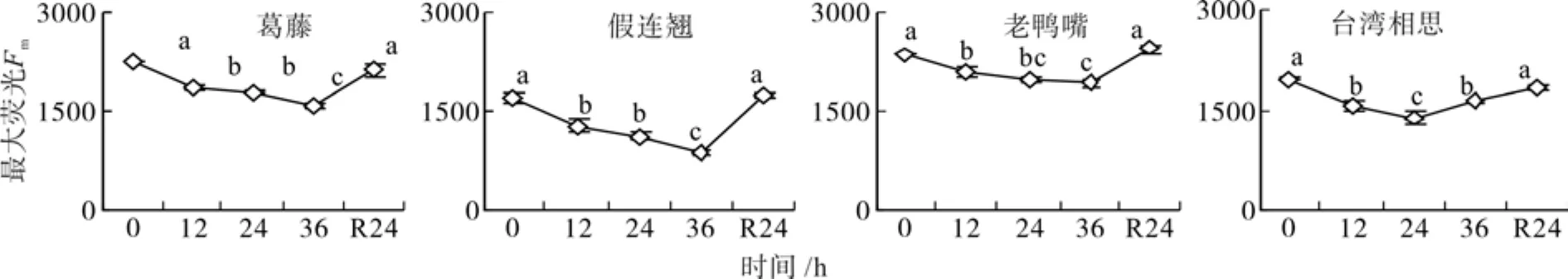
图1 低温胁迫下4种幼苗最大荧光的变化Fig. 1 Changes of maximal fl uorescence of four seedling types under low temperature stress
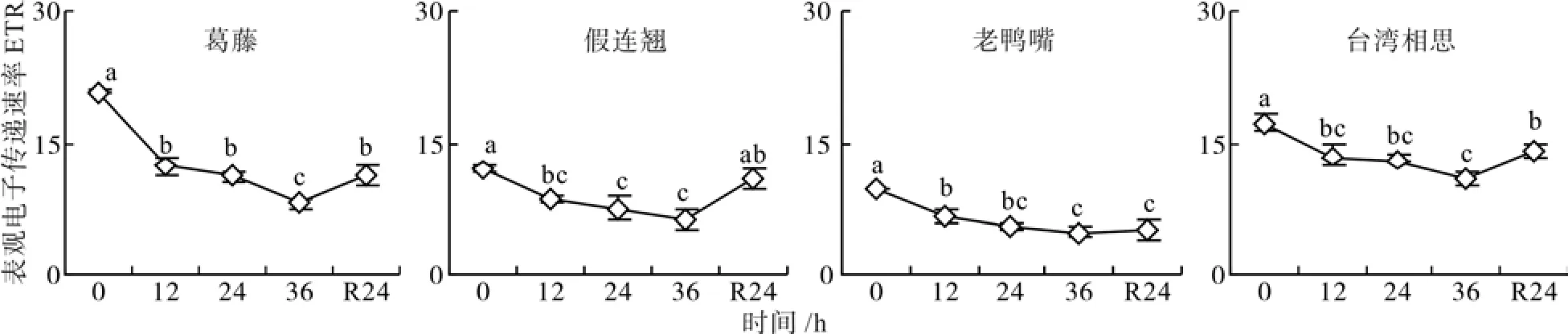
图2 低温胁迫下4种幼苗表观电子传递速率的变化Fig. 2 Changes of electron transport rate of four seedling types under low temperature stress
2.3 低温胁迫下种幼苗的非光化学淬灭变化
低温胁迫处理期间,4种幼苗的非光化学淬灭(NPQ)先上升后下降,均在24 h达到最大值(图3)。葛藤、假连翘和老鸭嘴24 h时的NPQ显著大于对照(p<0.05)。自然恢复24 h后,葛藤、老鸭嘴和台湾相思的NPQ保持稳定,假连翘的NPQ有所下降,除了台湾相思外,其余3种幼苗均显著高于对照。图3 低温胁迫下4种幼苗非光化学淬灭的变化Fig. 3 Changes of non-photochemical quenching of four seedling types under low temperature stress
2.4 低温胁迫下幼苗的PSII实际光量子效率变化
低温胁迫处理期间,4种幼苗的PSII实际光量子效率Y(II)逐渐降低,均在36 h达到最小值且显著小于对照(p<0.05),分别为对照的45%、65%、47%和48%(图4)。经过24 h自然恢复,葛藤和老鸭嘴的Y(II)有所回升,但仍显著低于对照水平(p<0.05),假连翘和台湾相思恢复到对照水平。
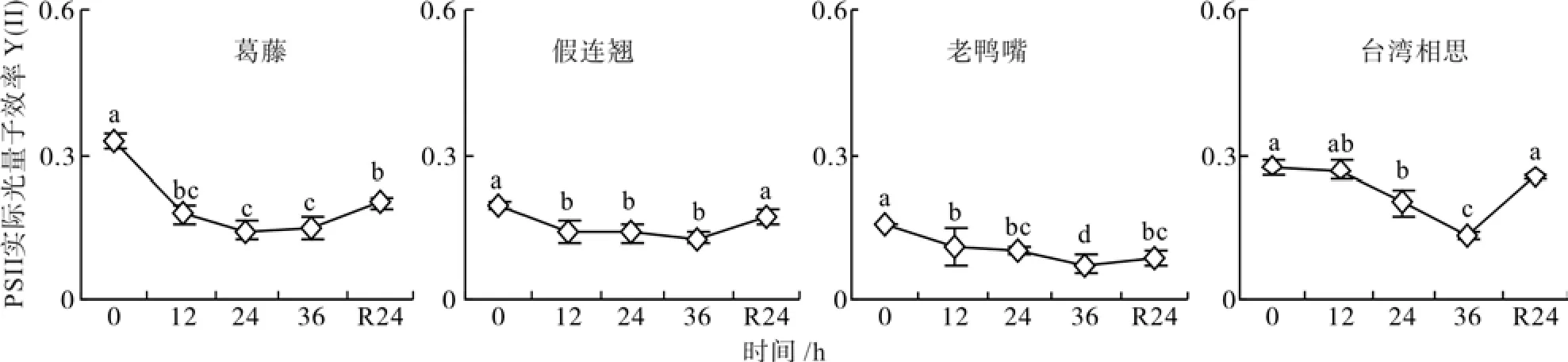
图4 低温胁迫下4种幼苗PSII实际光量子效率的变化Fig. 4 Changes of active quantum eff i ciency of PS II of four seedling types under low temperature stress
2.5 低温胁迫下幼苗的PSⅡ调节性能量耗散变化
低温胁迫处理期间,4种幼苗的PSⅡ调节性能量耗散Y(NPQ)逐渐升高,在36 h达到最大值(图5)。葛藤、假连翘和老鸭嘴的在12、24和36 h的Y(NPQ)均显著大于对照(P<0.05)。经过24 h恢复,4种幼苗的Y(NPQ)显著高于对照水平(P<0.05)。
2.6 低温胁迫下幼苗的PSII非调节性能量耗散变化
低温胁迫处理期间,4种幼苗的PSⅡ非调节性能量耗散Y(NO) 逐渐上升,在36 h达到最大值,且显著大于对照(P<0.05)(图6)。经过24 h恢复,老鸭嘴显著高于对照水平(P<0.05),其余3种幼苗恢复到对照水平。
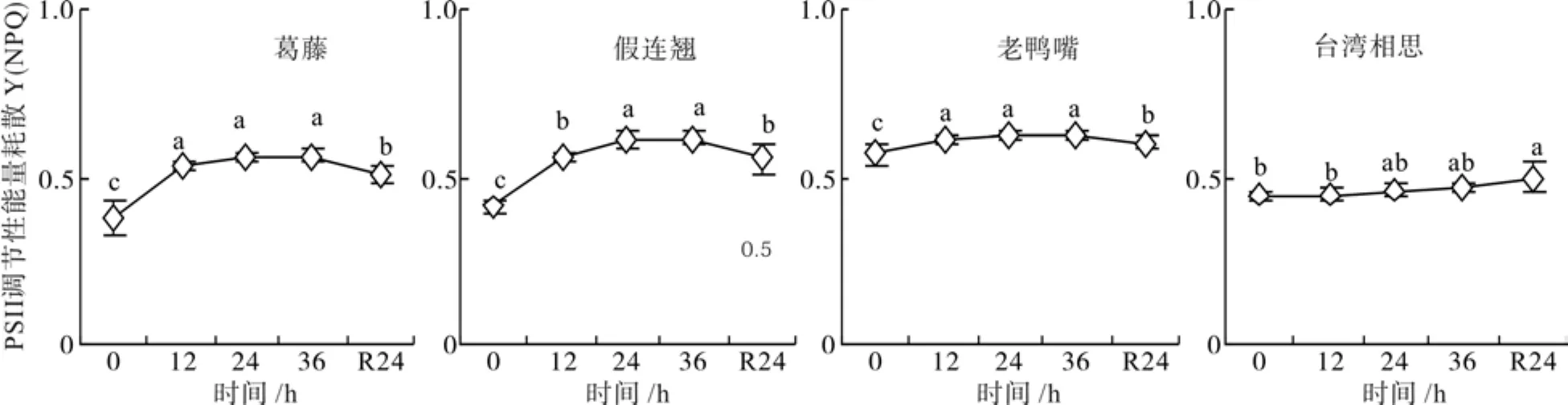
图5 低温胁迫下4种幼苗PSII调节性能量耗的变化Fig. 5 Changes of quantum yield of regulated energy of four seedling types under low temperature stress
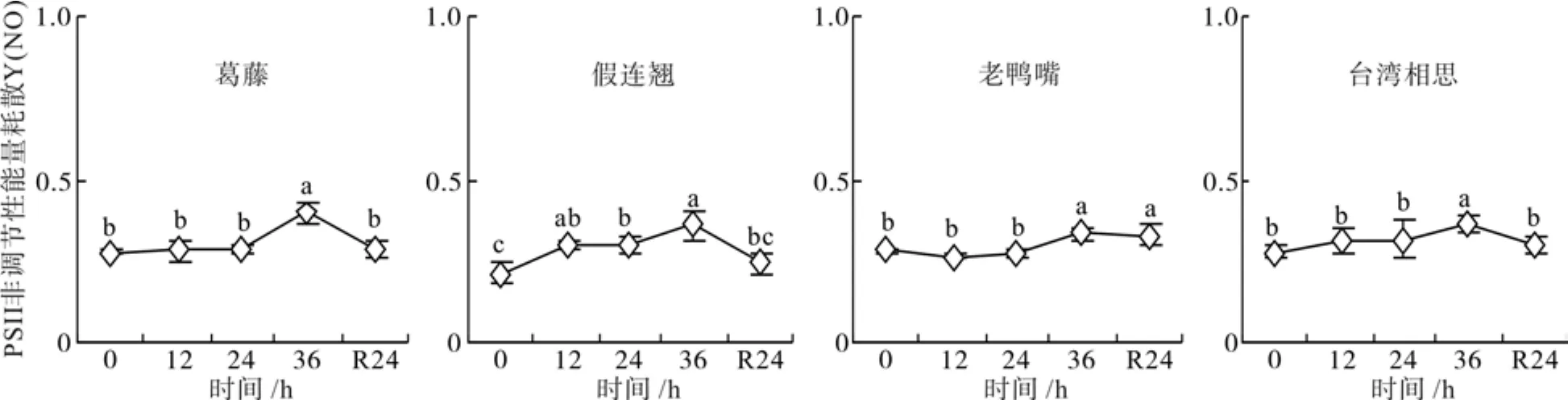
图6 低温胁迫下4种幼苗PSⅡ非调节性能量耗的变化Fig. 6 Changes of quantum yield of non-regulated energy dissipation of four seedlings types under low temperature stress
2.7 种幼苗抗寒性的综合评价
低温胁迫的程度与植物体内的Fm、Y(II)和ETR等参数的受抑制程度显著的相关,可作为植物抗逆的指标,但不同抗逆指标表现往往不一致[15]。利用主成分分析法可以全面而准确地分析植物的各种生理指标的综合抗寒性[10,16-17]。本文对所测量的低温胁迫下4种植物幼苗的6个荧光指标进行主成分分析,比较其抗寒性。利用极差法对各指标数值进行标准化,标准化公式为[18]:

式中,X为标准化后所得数据,x为某项指标的原始数据,xmin为某项指标原始数据的最小值,xmax为某项指标原始数据的最大值。
对标准化后的数据进行主成分分析。以累积贡献率达85%以上为参考值,确定主成分个数。由于主成分之间相互独立且有着不同的贡献率,因此,每个主成分得分就是其加权值,其中权重就是主成分对应的贡献率。由此分析得出4种幼苗抗寒性综合评价值大小顺序为台湾相思>假连翘>葛藤>老鸭嘴(表2)。

表2 4种幼苗抗寒性综合评价值及排序Table 2 Comprehensive assessment values and orders of cold resistance of four seedling types
3 结论与讨论
本研究中,低温胁迫导致反映PSII电子传递能力有关的荧光参数Fm和ETR均显著下降,这表明低温引起捕光天线和反应中心结构的变化,PSII的电子传递严重受阻并受到光抑制[19]。原因可能是低温导致类囊体膜的流动性减小,从而导致类囊体膜上的反应中心、捕光天线、电子传递体等蛋白的结构或构象发生变化,最终对PSII的功能造抑制[20]。低温胁迫下,4种幼苗的Y(II)下降,表明实际光化学转化效率的下降,阻止了植物同化力( NADPH、ATP) 的形成,影响了植物对碳的固定和同化[21]。4种幼苗中,假连翘的ETR和Y(II)的降幅较另外3种幼苗少,表明其在低温逆境下保持较快的光合电子传递速率,反应中心实际进行光化学反应的效率比另外3种幼苗高[22]。经过24 h的自然恢复,假连翘Y(II)和ETR和台湾相思的Y(II)均恢复到对照水平,表明其PSII反应中心失活为可逆失活,具有较好的抗寒性。与低温处理24 h相比,低温处理36 h时的葛藤的ETR和老鸭嘴的Y(II)迅速下降,表明两种幼苗在长时间的低温胁迫下,PSII的功能受损较严重。经过24 h自然恢复后,葛藤、老鸭嘴的Y(II)和ETR均未回复到对照水平,表明其PSII反应中心失活或破坏较严重,抗寒性差。
低温胁迫会引起植物叶片光能分配的变化。根据Kramer 等[23]的模型,采用叶绿素荧光参数将PSII 的量子产量分成3 部分,即PSII 实际光量子产量Y(II)、调节性能量耗散量子产量Y(NPQ)和非调节性能量耗散量子产量Y(NO)。低温胁迫下,用于光化学反应的光能部分显著减少,从而形成过剩光能。当大量过剩光能不能及时用于光化学反应时,就能形成光抑制现象,植物为了免受PSII受到破坏,往往通过叶黄素循环、荧光发射、光呼吸等热耗散途径来消耗过剩光能[24]。非光化学淬灭系数NPQ和调节性能量耗散量子产量Y(NPQ)表示PSII天然色素吸收的光能中不能用于光合电子传递而以热的形式耗散的部分,是植物的一种自我保护机制,对光合机构起一定的保护作用[7]。低温胁迫12和24 h后,4种幼苗的NPQ均有所上升,表明幼苗利用自身的保护机制使过剩的激发能通过非光化学途径耗散,阻止了由于激发能积累引起的PSII的损伤[25]。假连翘的NPQ和Y(NPQ)上升得较快,其自我保护能力较好。随着低温处理时间的延长,在36 h时4种幼苗的NPQ均有所下降,表明PSII天线热耗散的保护能力被显著削弱[19]。低温胁迫时间延长引起的NPQ下降,可能是由于低温抑制叶黄素循环的关键酶活性,影响了叶黄素循环[20]。与低温胁迫24 h相比,葛藤和假连翘在在36 h时的NPQ降幅均为13%,而老鸭嘴降幅为21%,说明前两种幼苗叶黄素循环的关键酶活性受损程度较轻,其抗寒性较好,而老鸭嘴的抗寒性差。经过24 h自然恢复,台湾相思的NPQ有所回升,表明其PSII反应中心失活为可逆的,自我恢复能力较好。低温胁迫期间,葛藤和假连翘的Y(NPQ) 较高,一方面表明植物接受的光强过剩,另一方面则说明这两种仍可以通过调节(如将过剩光能耗散为热) 来保护自身[26]。
Y(NO) 表示的是由于光钝化所引起的非调节性的量子产量,即电子传递到PSII后既不参与光化学反应、也不以叶黄素循环介导的调节性的热量耗散,而是以光呼吸、Mehler 反应等形式进行耗散或者是将电子传递给氧,参与活性氧形成[27-30],是光损伤的重要指标[21]。在本研究中,4种幼苗的Y(NO)均有所上升,而且在低温胁迫36 h时,均显著大于对照,表明光化学能量转换(用于光合电子传递)和保护性的调节机制(如热耗散) 不足以将植物吸收的光能完全消耗掉,使得叶绿素暂时积累并可能将能量传递给O2,形成破坏性极大的单线态氧(1O2),引起光合机构的失活甚至破坏[2]。4种幼苗中,低温胁迫36 h时的台湾相思幼苗的Y(NO)与24 h相比,升幅最少,表明其光损伤程度最低,抗寒性较好。经过24 h自然恢复,老鸭嘴的Y(NO)依然显著高于对照水平,且其Y(II)值亦显著低于对照,可能是其对光合机构造成了严重的破坏,光合电子传递得不到修复。其余3种幼苗恢复到对照水平,说明其光合电子传递及光保护机制运转正常。
综 上 比 较 4种 幼 苗 的 Fm、ETR、NPQ、Y(II)、Y(NPQ)和Y(NO)等一系列荧光参数,台湾相思在低温胁迫下ETR和Y(II)值较高、降幅少且自然恢复快,假连翘的NPQ和Y(NPQ)上升得较快,其自我保护能力较好,因此2种幼苗抗寒性好。低温胁迫使葛藤的ETR和老鸭嘴的Y(II)降幅大,自然恢复慢,低温解除后老鸭嘴的Y(NO)大,表明其光损伤严重,所以这2种幼苗抗寒性较差。主成分分析得出4种幼苗抗寒性为台湾相思>假连翘>葛藤>老鸭嘴,与4种幼苗荧光参数的表现一致,说明主成分分析是一种综合评价植物生理特性的有效方法。
[1] 周 建, 杨立峰, 郝峰鸽, 等. 低温胁迫对广玉兰幼苗光合及叶绿素荧光特性的影响[J]. 西北植物学报, 2009, 29(1): 0136-0142.
[2] 吴雪霞, 陈建林, 查丁石. 低温胁迫对茄子幼苗叶片叶绿素荧光特性和能量耗散的影响[J]. 植物营养与肥料学报,2009,15(1): 164-169.
[3] 周蕴薇, 刘艳萍, 戴思兰. 用叶绿素荧光分析技术鉴定植物抗寒性的剖析[J]. 植物生理学通讯, 2006, 42(5): 945- 950.
[4] 莫亿伟, 郭振飞, 谢江辉. 温度胁迫对柱花草叶绿素荧光参数和光合速率的影响[J]. 草业学报, 2011, 20(1): 96-101.
[5] 胡文海, 肖宜安, 喻景权, 等. 低夜温后不同光强对榕树叶片PSII功能和光能分配的影响[J]. 植物研究,2005,25(2):159- 162.[6] 王 贺, 刘国成, 吕德国, 等. 低温对寒富苹果及其亲本叶绿素荧光特性的影响[J]. 华北农学报, 2008, 23(2): 38-40.
[7] 马博英, 金松恒, 徐礼根, 等. 低温对三种暖季型草坪草和绿素荧光特性的影响[J]. 中国草地学报, 2006, 28(1): 58-62.
[8] 陈 曦, 陈彬丽, 周 晨, 等. 生长季油松针叶叶绿素荧光对温度变化的响应[J]. 北京林业大学学报, 2011, 33(6):70-74.
[9] 梁 芳, 郑成淑, 孙宪芝, 等. 低温弱光胁迫及恢复对切花菊光合作用和叶绿素荧光参数的影响[J]. 应用生态学报, 2010,21(1): 29-35.
[10] 邵怡若, 许建新, 薛 立, 等. 低温胁迫时间对4种幼苗生理生化及光合特性的影响[J]. 生态学报,2013,33(14):4237-4247.[11] 卢广超, 许建新, 薛 立, 等.干旱胁迫下4种常用植物幼苗的光合和荧光特性综合评价[J]. 生态学报, 2013,33(24):7872-7881.
[12] 李 玲, 徐志防, 韦 霄, 等. 金钟藤和葛藤在干旱与复水条件下的生理比较[J]. 广西植物, 2008, 28(6): 806 - 810
[13] 王 齐, 王有国, 师春娟, 等. 中水水培对4 种绿地植物生长及光合生理特性的影响[J]. 草业学报, 2010, 19(6): 106-113.
[14] 卢广超, 许建新, 薛 立, 等. 干旱胁迫和复水后台湾相思、银合欢和火棘幼苗的生理变化[J]. 林业科技开发, 2013,27(1): 55-59.
[15] 左胜鹏, 叶良涛, 马永清. 不同生态型化感冬小麦抽穗期的荧光动力学特性[J]. 中国生态农业学报, 2011, 19(2): 331- 337.
[16] 何跃君, 薛 立, 任向荣, 等. 低温胁迫对6种幼苗生理特性的影响[J]. 生态学杂志, 2008, 27(4): 524- 531.
[17] 冯慧芳, 薛 立, 任向荣, 等. 4种阔叶苗木对PEG模拟干旱的生理响应[J].生态学报, 2011, 31(2): 371- 382.
[18] 谭雪红, 高艳鹏, 郭小平, 等. 五种高速公路边坡绿化植物的生理特性及抗旱性综合评价[J]. 生态学报, 2012, 32(16):5076-5086.
[19] 许培磊, 焦竹青, 王振兴, 等. 五味子休眠前叶片的叶绿素荧光参数变化研究[J]. 华北农学报, 2012, 27 (增刊) : 209-212.
[20] 陈 梅, 唐运来. 低温胁迫对玉米幼苗叶片叶绿素荧光参数的影响[J]. 内蒙古农业大学学报, 2012, 33(3): 20-24.
[21] 刘爱荣, 陈双臣, 王淼博, 等. 高温胁迫对番茄幼苗光合作用和叶绿素荧光参数的影响[J]. 西北农业学报, 2010, 19(5):145-148.
[22] 齐红岩, 华利静, 赵 乐, 等. 夜间低温对不同基因型番茄叶绿素荧光参数的影响[J]. 华北农学报, 2011, 26(4): 222-227.
[23] Kramer D M, Johnson G, Kiirats O, et al. New fluorescence parameters for the determination of QA redox state and excitation energy fl uxes[J]. Photosynthesis Research, 2004, 79(2): 209-218.
[24] Singh S, Chaudhary H K, Sethi G S. Distribution and allelic expressivity of genes for hybrid necrosis in some elite winter and spring wheat ecotypes[J]. Euphytica, 2000, 112(1): 95-100.
[25] Costa E S, Bressan-Smith R, Oliveira J G, et al. Chlorophyll a fl uorescence analysis in response to excitation irradiance in bean plants (Phaseolus vulgaris L. and Vigna unguiculata L. Walp)submitted to high temperature stress[J]. Photosynthetica, 2003,41(1): 77-82.
[26] 官莉莉, 邓绍龙, 杨振意, 等. 五种观花树种光合与叶绿素荧光特征研究[J]. 广东林业科技, 2012, 28(4): 1-3.
[27] Wang L J, Loescher W, Duan W, et al. Heat acclimation induced acquired heat tolerance and cross adaptation in different grape cultivars: relationships to photosynthetic energy partitioning[J].Functional Plant Biology, 2009, 36(6): 516- 526.
[28] Busch F, Hunter N P A, Ensminger I. Biochemical constrains limit the potential of the photochemical reflectance index as a predictor of effective quantum efficiency of photosynthesis during the winter-spring transition in Jack pine seedlings[J].Functional Plant Biology, 2009, 36(11): 1016- 1026.
[29] 许鹏波, 薛 立, 潘 澜, 等. 稀土对低温胁迫麻楝幼苗生理生化特性的影响[J]. 中南林业科技大学学报, 2011,31(2):34-40.
[30] 姜 英, 郝海坤, 黄志玲, 等. 红锥苗期生长特性和叶绿素荧光对不同光强的响应[J]. 中南林业科技大学学报, 2013,33(10): 61-65.
Effects of low temperature stress on chlorophyll fl uorescence characteristics of four types of tree species seedlings
LU Guang-chao1, XU Jian-xin1,2, XUE Li1, ZHANG Rou1, WU Chai-qiong2, SHAO Yi-ruo1
(1. College of Forestry, South China Agricultural University, Guangzhou 510642, Guangdong, China;2. Shenzhen Tech and Ecology, Environment CO. LTD., Shenzhen 518040, Guangdong, China)
In order to provide the scientif i c basis for selecting cold resistance seedlings type, the chlorophyll fl uorescence characteristics ofPueraria lobata,Duranta repens,Thunbergia erectaandAcacia confusewere studied under low temperature stress. The results showed that with increasing time of low temperature stress, the chlorophyll fl uorescence characteristics of maximal fl uorescence(Fm),electron transport rate(ETR)and active quantum eff i ciency of PSII (Y(II)) of four seedlings types decreased gradually, whereas their non-photochemical quenching (NPQ) fi rst rose and then declined, and the quantum yield of regulated energy dissipation (Y(NPQ)) and quantum yield of non-regulated energy dissipation (YNO) increased gradually; After 24 hours’ natural recovery, theFmrecovered to the control level, ETR rose, whereas the Y(NPQ) was signif i cantly higher than control level for the four seedlings types; The value of NPQ ofP. lobata,T. erectaandA. confuseremained stable, whereas that ofD. repensdeclined; The Y(II) ofD. repensandA. confuse,and the Y(NO) ofP. lobata,D. repensandA. confuserecovered to the control level; The chlorophyll fl uorescence parameters of the four seedlings types were evaluated with principal component analysis, the order of low temperature resistance of the four seedling types wasA. confuse>D. repens>P. lobata>T. erecta.
low temperature stress; seedlings of tree; chlorophyll fl uorescence parameters; principal component analysis
S718.43
A
1673-923X(2014)02-0044-06
2013-07-06
深圳市战略新兴产业发展专项资金项目“深圳市生态修复生物工程技术研究开发中心”(GCZX20120618100801416);国家科学技术部农业科技成果转化资金项目“裸露坡面生态恢复新技术应用研究”(2009GB2E020423)
卢广超(1989-),男,硕士研究生,主要从事森林培育和树木生理学研究
薛 立(1958-),男,教授,博导,主要从事林分密度和自然稀疏及林木养分研究;E-mail:forxue@scau.edu.cn
[本文编校:吴 彬]
猜你喜欢
中国药房(2022年14期)2022-07-29
河北农机(2021年11期)2021-11-29
润·文摘(2021年9期)2021-09-22
孩子·小学版(2019年11期)2019-09-10
农业机械学报(2018年11期)2018-12-04
红岩(2018年6期)2018-11-16
安徽医科大学学报(2015年9期)2015-12-16
中华胰腺病杂志(2015年5期)2015-12-08
中国医疗美容(2015年1期)2015-07-12
中国医疗美容(2015年1期)2015-07-12
