
不同精粗比全混合颗粒饲粮对母羊营养物质表观消化率、氮代谢和能量代谢的影响
2014-12-20 12:25王文奇侯广田罗永明刘艳丰卡那提沙力克
动物营养学报 2014年11期
王文奇 侯广田 罗永明 刘艳丰 卡那提·沙力克
(1.新疆畜牧科学院饲料研究所,乌鲁木齐 830000;2.新疆动物生物技术实验室,乌鲁木齐 830000;3.新疆畜牧科学院畜牧研究所,乌鲁木齐 830000)
饲粮营养平衡是反刍动物生产的基础,饲粮中精饲料比例不适宜或粗饲料使用不适当均会导致反刍动物瘤胃健康受损、营养物质消化率降低及生产性能的下降,甚至会影响反刍动物繁殖性能的发挥。因此,适宜的精粗比是科学饲养反刍动物的基础,粗饲料的合理使用及其与精饲料的合理配比对优化反刍动物瘤胃环境和提高动物生产性能具有非常重要的意义。对于反刍动物而言,粗饲料是反刍动物重要的营养源,对刺激反刍和咀嚼、维持瘤胃液正常pH、维持瘤胃微生物正常发酵、促进营养物质消化代谢均具有重要作用。中性洗涤纤维(NDF)是粗饲料代表性营养指标,其在反刍动物营养中的应用受到了广泛重视,许多研究表明,饲粮NDF水平对反刍动物的采食量[1-2]、唾液分泌和瘤胃内环境[3]、营养物质消化率等均会造成影响,适宜的饲粮NDF水平有助于维持机体正常生理功能和保障动物健康。目前,国内外对于反刍动物适宜精粗比的研究多集中在奶牛及肉牛方面,在肉羊及母羊营养方面的研究报道相对较少。鉴于此,本试验通过研究不同精粗比全混合颗粒饲粮对母羊营养物质表观消化率、氮代谢和能量代谢的影响,旨在探讨母羊饲粮适宜的精粗比和NDF水平,为实际生产中母羊饲粮的配制提供基础理论指导及数据支持。
1 材料与方法
1.1 试验时间与地点
本试验于2012年7月在新疆畜牧科学院科创畜牧繁育中心进行,共历时30 d。
1.2 试验设计与动物
选择30只体况良好、8月龄、平均体重为(49.63±2.38)kg 的杂交母羊(萨福克羊×阿勒泰羊),随机分为6个处理(每个处理5只羊),分别饲喂 6 种不同精粗比(85∶15、70∶30、55∶45、40∶60、25∶75、10∶90)的全混合颗粒饲粮。试验饲粮组成及营养水平见表1。预试期10 d,正试期20 d。在正试期第 1、3、5、7、9、11、13、15、17、19 天分 10 批(每批3只羊),将羊分别移入3个带头箱的代谢笼内进行为期2 d的呼吸测热试验,适应24 h,随后的24 h采集气体。除了试验羊进入代谢笼内的2 d,其余18 d采用全收粪尿法进行消化代谢试验。

表1 试验饲粮组成及营养水平(干物质基础)Table 1 Composition and nutrient levels of experimental diets(DM basis) %
1.3 饲养管理
试验开始前,对所有试验羊进行驱虫,然后进行单笼饲喂,自由饮水。由于饲粮营养水平的差异性会造成各处理试验羊采食量的不同,所以在预试期预先测定各处理试验羊的采食量,以最低的一个处理的采食量(1.4 kg)作为正试期的饲喂量。试验羊每天饲喂2次(09:00和21:00),每次饲喂0.7 kg。每天准确记录每只羊采食量及剩料量。
1.4 样品采集与处理
1.4.1 消化代谢试验
分别在正试期第1、10和20天采集各处理饲粮样本,混匀后,作为整个试验期的饲粮样本,备测;将每天收集得到的剩料混匀后作为剩料样本,备测;采用全收粪尿法,收集每只羊每天排粪,混匀后按质量的5%取样,将每只羊18 d的粪样混匀,-20℃保存,备测;用盛有20 mL 6 mol/L的硫酸(H2SO4)的塑料桶收集尿液,每天记录每只羊的尿液体积后混匀,按体积的5%取样,将每只羊18 d尿样混匀,-20℃保存,备测。
1.4.2 呼吸测热试验
试验羊在代谢笼内适应24 h后,测定随后24 h的甲烷产量、氧气(O2)消耗量及二氧化碳(CO2)生成量。试验羊进入和离开代谢笼时均称重,平均值作为该羊的测热体重。
1.5 测定指标和方法
1.5.1 营养物质表观消化率
饲粮、剩料和粪样中干物质(DM)、粗蛋白质(CP)、有机物(OM)、粗脂肪(EE)、NDF、粗灰分(ash)含量的测定参照《饲料分析及饲料质量检测技术》[4]中的方法。总能(GE)的测定用氧弹测热法进行。饲粮非纤维性碳水化合物(NFC)含量按照如下公式计算:

饲粮中各营养物质的表观消化率按如下公式计算:

1.5.2 氮代谢
测定摄入氮、粪氮、尿氮,计算消化氮、氮消化率、沉积氮、沉积氮/摄入氮、沉积氮/消化氮。
1.5.3 能量代谢
粪能采用氧弹测热法测定。取5块定量滤纸分别测定能值,计算出滤纸的平均能值。将10 mL尿液滴在滤纸上,65℃烘干后于氧弹测热法仪中测定,得到滤纸和尿液的总能值,减去滤纸能即得尿能。用开放式呼吸测热系统(美国Sable公司)测定甲烷产量。计算公式[6]如下:


1.6 数据统计分析
试验数据用Excel 2007初步整理后,利用SPSS 18.0统计软件进行单因素方差分析和显著性检验,多重比较采用one-way ANOVA中的LSD法,试验结果以平均值±标准差表示。P<0.05和P<0.01分别为差异显著和极显著水平。
2 结果与分析
2.1 营养物质表观消化率
由表2可知,全混合颗粒饲粮精粗比对肉用绵羊DM、OM、NDF表观消化率均产生了极显著影响(P<0.01),DM、OM 的表观消化率均随着饲粮精粗比的升高而出现极显著升高(P<0.01)。1号、2号和3号饲粮DM表观消化率均极显著高于4号、5号和 6号饲粮(P<0.01),5号和 6号饲粮DM表观消化率差异不显著(P>0.05);1号饲粮OM 表观消化率极显著高于其他饲粮(P<0.01),2号饲粮极显著高于4号、5号和6号饲粮(P<0.01),5号和6号饲粮OM 表观消化率差异不显著(P>0.05);3号饲粮NDF表观消化率最高,与1号和2号饲粮相比均差异不显著(P>0.05),显著高于4号饲粮(P<0.05),极显著高于5号和6号饲粮(P<0.01)。
2.2 氮代谢
由表3可知,随着摄入氮的极显著降低(P<0.01),尿氮的排出量略有下降,粪氮和尿氮的排出量各组间差异不显著(P>0.05);1号饲粮消化氮最高,显著或极显著高于其他饲粮(P<0.05或P<0.01),2 号和 3 号饲粮差异不显著(P>0.05),但均极显著高于4号、5号和6号饲粮(P>0.01);1号饲粮氮消化率与2号和3号饲粮差异不显著(P>0.05),显著或极显著高于 4号、5号和 6号饲粮(P<0.05 或 P<0.01);沉积氮在 3 号饲粮最高,显著高于6号饲粮(P<0.05),但与其他饲粮差异不显著(P>0.05);3号饲粮沉积氮/摄入氮和沉积氮/消化氮均最高,但与其他饲粮相比均差异不显著(P>0.05)。

表2 不同精粗比全混合颗粒饲粮对母羊营养物质表观消化率的影响Table 2 Effects of different concentrate to roughage ratios of total mixed pellet diet on nutrient apparent digestibility of ewes
2.3 能量代谢
如表4可知,在限饲条件下,全混合颗粒饲粮精粗比显著或极显著影响了各能量代谢指标(P<0.05或 P<0.01)。5号、6 号饲粮总能摄入量极显著高于1号、2号、3号饲粮(P<0.01),而 1号、2号和3号饲粮间差异不显著(P>0.05);粪能随精粗比的下降,出现极显著的上升(P<0.01),1号、2号、3号饲粮差异不显著(P>0.05),均显著或极显著低于其他饲粮(P<0.05 或 P<0.01);1 号饲粮尿能最低,显著低于3号饲粮(P<0.05),其他饲粮间差异不显著(P>0.05);甲烷能 5号饲粮最高,3号、4号、5 号、6 号饲粮差异不显著(P>0.05),但均极显著高于1号和2号饲粮(P<0.01);消化能和代谢能均随精粗比的下降出现极显著的下降(P<0.01),1号、2 号、3 号饲粮消化能均极显著高于6号饲粮(P<0.01),3号、4号、5号和 6号饲粮代谢能差异不显著(P>0.05),均极显著低于1号饲粮(P<0.01);甲烷能占总能比例5号饲粮最高,与3号、4号和6号饲粮差异不显著(P>0.05),极显著高于1号和2号饲粮(P<0.01);总能消化率随精粗比的下降出现极显著的下降(P<0.01),2号和3号饲粮差异不显著(P>0.05),1号、2号和3号饲粮极显著高于5号和6号饲粮(P<0.01);总能代谢率也随精粗比的下降出现显著的下降(P<0.01),1号和2号饲粮极显著高于6号饲粮(P<0.01);各饲粮消化能代谢率差异显著(P<0.05),1 号饲粮分别显著(P<0.05)和极显著(P<0.01)高于3号和6号饲粮,与2号、4号、5号饲粮差异不显著(P>0.05)。

表3 不同精粗比全混合颗粒饲粮对母羊氮代谢的影响Table 3 Effects of different concentrate to roughage ratios of total mixed pellet diet on nitrogen metabolism of ewes
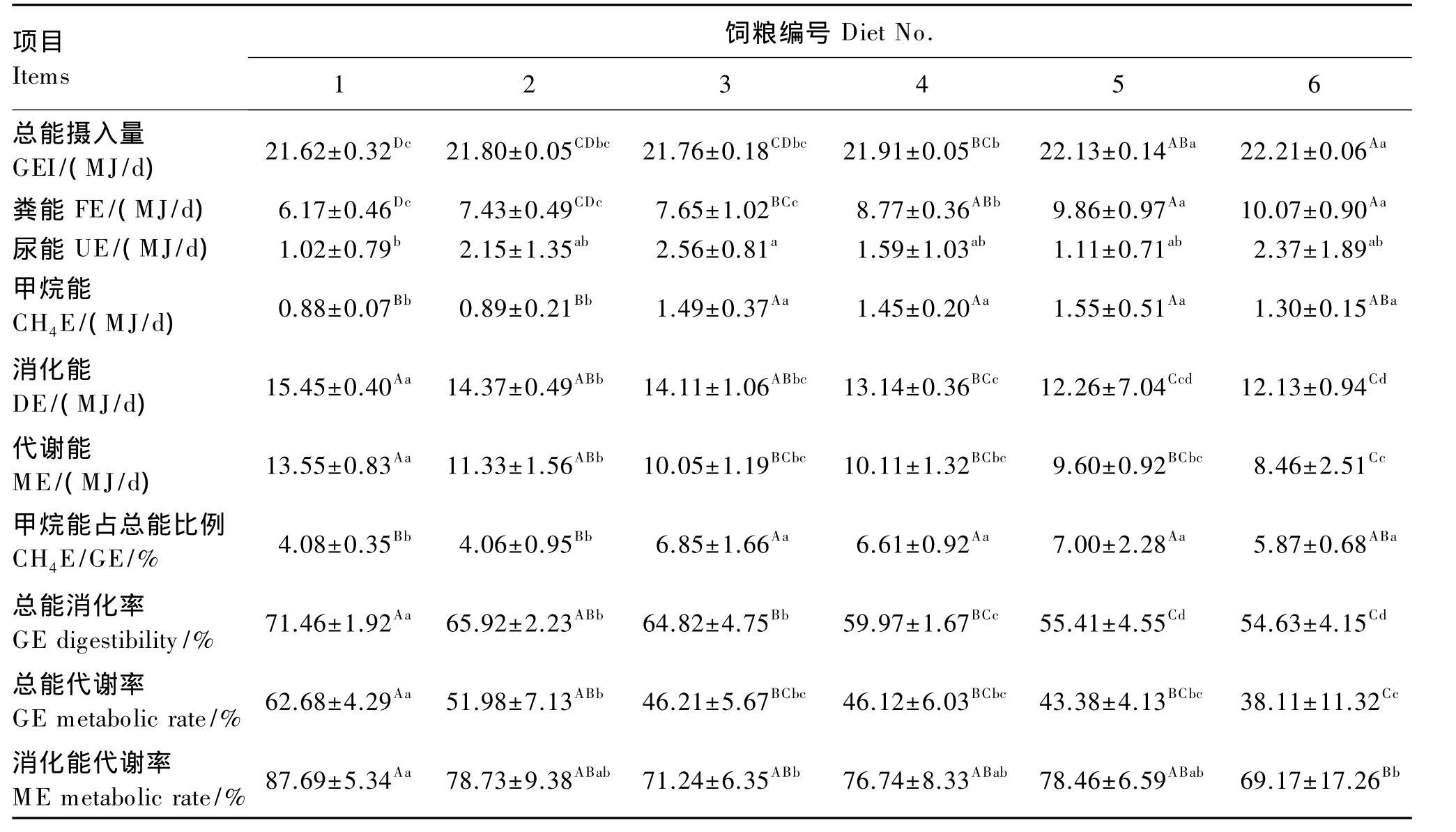
表4 不同精粗比全混合颗粒饲粮对母羊能量代谢的影响Table 4 Effects of different concentrate to roughage ratios of total mixed pellet diet on energy metabolism of ewes
3 讨论
3.1 不同精粗比全混合颗粒饲粮对DM和OM表观消化率的影响
饲粮中NDF和NFC水平因饲粮精粗比的不同而存在差异。随着饲粮精粗比的降低,NDF水平提高,NFC水平降低,饲粮纤维含量增加时,饲粮非结构性碳水化合物(NSC)的比例减少,结构性碳水化合物(SC)的比例增加,降低饲粮在瘤胃的滞留时间,加快饲粮在动物胃肠道中的流通速率,降低DM和OM的降解率,进而降低营养物质肠道消化率和全消化道消化率[7]。Valdés等[8]研究了精粗比分别为80∶20、60∶40、40∶60 和 20∶80 的饲粮对绵羊营养物质的消化代谢的影响,结果表明随饲粮精粗比的下降,DM、OM表观消化率降低。门小明等[9]、Yang 等[10]、刘洁等[11]、张立涛等[12]也获得类似的结果。本试验设计的6种饲粮,随着精粗比的降低,饲粮NDF水平由33.96%提高 到 53.29%,NFC 水 平 由 38.97% 降 低 到18.81%,饲粮DM和OM的表观消化率呈现显著降低的趋势,这与上述研究报道一致。
有研究表明,当饲粮中精饲料比例超过70%时,动物DM、OM的消化率都有不同程度地降低[13-14]。张立涛等[12]研究发现,肉羊饲粮 NDF水平为26.51%时的DM和OM的表观消化率显著低于为33.35%时,进一步验证了饲粮过低的NDF水平或过高的NFC水平会降低饲粮DM和OM的表观消化率。在本试验中,在精粗比为85∶15的高精饲料条件下,饲粮中 NDF水平为33.96%,稍高于张立涛等[12]所报道的 33.35%,母羊DM、OM表观消化率仍较高,这与上述研究报道一致。
3.2 不同精粗比全混合颗粒饲粮对NDF表观消化率的影响
反刍动物饲粮纤维的消化受饲粮组成成分、瘤胃发酵环境、瘤胃微生物组成以及饲粮在瘤胃内滞留时间等多方面的影响。有研究发现,反刍家畜在采食较高精饲料比例饲粮时,大量可发酵碳水化合物在瘤胃内发酵,会导致瘤胃液pH降低[15],抑制纤维素分解菌的活性,致使NDF表观消化率下降。然而,适宜的精饲料比例,会增加过瘤胃蛋白质、脂肪等营养物质的数量,为后肠道尤其是盲肠提供更适宜的发酵环境,促进盲肠微生物的生长[16],增强对NDF的消化,使NDF在后肠消化道得到补偿,从而对全消化道NDF表观消化率产生较大影响。在动物DM采食量相同的情况下,NDF的表观消化率会随着饲粮中精饲料比例的增加而下降,但如果在自由采食情况下,NDF的表观消化率一般不受饲粮精粗比的影响[17],这在Valdés等[8]、门小明等[9]、Yang 等[10]的研究报道中得到了验证。本试验研究采用6种饲粮对母羊进行限饲,随着精粗比的下降,NDF摄入量和排出量均增加,NDF表观消化率表现先下降再上升后下降的趋势,在NDF水平为45.80%时NDF表观消化率最高。霍鲜鲜等[18]、谭支良[19]认为反刍动物饲粮中有一个理想的精粗比,更确切地说是有理想的SC/NSC,使得纤维物质的表观消化率最高。在本试验中,在精粗比为55∶45时NDF表观消化率最高,与以上论述相符合。
3.3 不同精粗比全混合颗粒饲粮对氮代谢的影响
反刍动物对蛋白质的消化代谢受瘤胃微生物对其降解作用的影响。在瘤胃内,瘤胃可降解蛋白质(RDP)被蛋白质分解菌分解成氨、氨基酸和小肽,然后利用碳水化合物降解产生的能量与之合成微生物蛋白(MCP)[20],MCP与饲粮中的瘤胃非降解蛋白质(UDP)进入后消化道继续消化,到达小肠可消化蛋白质中MCP的比例对蛋白质消化和利用率影响较大。因此,反刍动物能否充分消化吸收饲粮蛋白质,主要取决于RDP是否最大效率的用于合成MCP。RDP的利用率受饲粮NDF水平或者是饲粮中SC/NSC影响较大,这是因为瘤胃微生物对饲粮瘤胃可降解氮(RDN)和可发酵有机物(FOM)的需要量有一定的比例关系,而饲粮 NDF水平则会影响RDN/FOM,从而对RDP的利用率产生间接影响。当RDN与FOM的释放速度同步时,RDP的利用率最好,饲粮蛋白质的消化率和利用率最高。本试验中,随精粗比的增加,摄入氮量极显著增加,这是由于高精饲料饲粮氮含量高,且各饲粮风干物质采食量相同造成的;饲粮蛋白质摄入量的增加,对肉羊粪氮和尿氮的排出量影响不显著,且尿氮的排出量明显高于粪氮,全消化道消化氮呈极显著增加趋势,这与Reynolds等[21]、Devant等[22]、黄洁等[23]的报道相符合。张立涛等[12]报道,分别饲喂肉羊不同梯度的NDF水平的饲粮,随着NDF水平的升高,饲粮中NFC的比例逐渐降低,CP表观消化率呈现显著的下降趋势,这与本试验结果一致。本试验中沉积氮、沉积氮/摄入氮和沉积氮/消化氮均随着饲粮摄入氮的增加,呈先增加后降低的趋势,这与杨膺白等[24]和 Jetana等[25]报道并不一致。这一方面可能是由于饲粮SC/NSC的差异性造成瘤胃内微生物合成自身MCP量不同,饲粮RDP的利用率也存在差异;另一方面可能是进入后肠道(盲肠、结肠和直肠)含氮物质成分、含量以及氨气(NH3)在后肠道再吸收量存在差异,后肠道利用再吸收的氮合成非必需氨基酸或再循环到瘤胃合成MCP量造成不同,影响了饲粮氮的利用率[26]。
3.4 不同精粗比全混合颗粒饲粮对能量代谢的影响
反刍动物在采食饲粮后,饲粮内的蛋白质、碳水化合物和脂肪在动物机体内经一系列的消化和代谢作用产生的能量,最终以ATP的形式满足机体的需要。其中可发酵碳水化合物产生的乙酸、丙酸、丁酸等挥发性脂肪酸(VFA)是反刍动物主要的能量来源,冯仰廉[27]认为VFA能量约占动物摄入消化能的70%~80%。大量研究发现,饲粮的精粗比对VFA的产量没有太大的影响,但对乙酸、丙酸和丁酸等VFA组成比例影响很大。当反刍动物饲粮中NDF水平较高时,乙酸/丙酸较高,其能量利用率较低,反之利用率则较高。因此,饲粮类型对动物能量代谢的影响较大。
粪能约占饲粮摄入总能的1/3[28],是饲粮能量中损失最大的部分,粪能排出量的高低与饲粮原料组成及特性有很大关系。反刍动物采食精饲料时粪能为20%~30%,采食粗饲料时粪能占总能的40% ~50%,采食低质粗饲料时为 60%[29]。Tyrrell等[30]、许贵善等[31]报道,总能摄入量的增加会造成粪能排出量的相应增加。本试验结果显示,随着饲粮精粗比的降低,总能摄入量极显著增加,粪能的排出量极显著增加,粪能占总能的比例为 28.53% ~45.37%,且随着精粗比的降低而上升。
尿能主要为尿中含氮有机物的能量,其排出量受饲粮结构,尤其是饲粮蛋白质水平、氨基酸平衡状况、能量摄入状况等的影响。本试验中,尿能排出量在精粗比为85∶15时最高,显著高于精粗比为55∶45时,而与其他处理差异不显著,尿能占摄入总能的比例为 4.73% ~11.78%,高于 3% ~5%[30]的报道。
甲烷是饲粮在反刍动物瘤胃中发酵的主要能量损失,其产量受饲粮精粗比的影响,以甲烷形式损失的能量约占饲粮总能的2%~15%[32],特别是饲粮糖和淀粉的含量[33],改变饲粮精粗比,会使SC/NSC发生变化,从而显著影响瘤胃发酵。饲粮中精饲料含量的增加会改变瘤胃内VFA组成,造成乙酸比例下降,丙酸比例上升,而丙酸是氢的受体,导致甲烷合成的底物氢减少,从而减少甲烷产量;而饲粮中粗饲料比例增加,则瘤胃VFA变化却恰恰相反,造成乙酸比例的增加,产生大量氢,瘤胃氢分压升高,刺激甲烷菌大量增殖,甲烷产量增加。饲粮精饲料比例与瘤胃甲烷产量呈负相关[32,34]。本试验中,随着精粗比的提高,甲烷能整体表现下降的趋势,甲烷能占总能比例为4.06%~7.00%。
王吉峰[35]研究了精粗比分别为 30∶70、50∶50、65∶35的饲粮对泌乳奶牛能量代谢的影响,发现消化能和总能消化率均随着精饲料比例的增加而显著提高。刘洁[36]研究了不同精粗比饲粮对肉用绵羊能量消化代谢的影响,发现随着精饲料比例的提高,饲粮消化能、代谢能和总能消化率整体表现上升的趋势。本试验结果显示,高精粗比饲粮的总能消化率、总能代谢率、消化能和代谢能均显著高于低精饲料比饲粮,精粗比的改变对能量消化和代谢会产生显著影响。
4 结论
①全混合颗粒饲粮精粗比对母羊DM、OM和NDF表观消化率均产生了极显著影响,DM和OM表观消化率随精粗比的上升而升高,精粗比为55∶45的饲粮NDF表观消化率最高。
②消化氮随着精粗比的上升而升高,过高或过低精粗比均不利于氮的沉积,精粗比为55∶45时,母羊对氮的利用效率最高。
③消化能、代谢能、总能消化率和总能代谢率均随着精粗比的上升而出现极显著的升高。
④在本试验条件下,母羊对精粗比为70∶30和 55∶45(NDF 水平分别为 37.78%和 45.80%)的全混合颗粒饲粮消化吸收较好。
[1] TJARDES K E,BUSKIRK D D,ALLEN M S,et al.Neutral detergent fiber concentration of corn silage and rumen inert bulk influences dry matter intake and ruminal digesta kinetics of growing steers[J].Journal of Animal Science,2002,80(3):833-840.
[2] AITCHISON E M ,GILL M ,DHANOA M S ,et al.The effect of digestibility and forage species on the removal of digesta from the rumen and the voluntary intake of hay by sheep[J].British Journal of Nutrition,1986,56(2):463-476.
[3] BEAUCHEMIN K A.Effects of dietary neutral detergent fiber concentration and alfalfa hay quality on chewing,rumen function,and milk production of dairy cows[J].Journal of Dairy Science,1991,74(9):3140-3151.
[4] 张丽英.饲料分析及饲料质量检测技术[M].3版.北京:中国农业大学出版社,2007.
[5] NRC.Nutrient requirements of dairy cattle[S].Washington,D.C.:National Academy Press,2001.
[6] 杨嘉实,冯仰廉.畜禽能量代谢[M].北京:中国农业出版社,2004.
[7] 祁茹,林英庭.日粮物理有效中性洗涤纤维对奶牛营养调控的研究进展[J].粮食与饲料工业,2010(5):52-55.
[8] VALDÉS C,CARRO M D,RANILLA M J,et al.Effect of forage to concentrate ratio in complete diets offered to sheep on voluntary food intake and some digestive parameters[J].Journal of Animal Science,2000,70(1):119-126.
[9] 门小明,雒秋江,唐志高,等.3种不同精粗比日粮条件下空怀小尾寒羊母羊的消化与代谢[J].中国畜牧兽医,2006,33(10):13-17.
[10] YANG W Z,BEAUCHEMIN K A,RODE L M.Effects of grain processing,forage to concentrate ratio,and forage particle size on rumen pH and digestion by dairy cows[J].Journal of Dairy Science,2001,84(10):2203-2216.
[11] 刘洁,刁其玉,赵一广,等.肉用绵羊饲料养分消化率和有效能预测模型的研究[J].畜牧兽医学报,2012,43(8):1230-1238.
[12] 张立涛,李艳玲,王金文,等.不同中性洗涤纤维水平饲粮对肉羊生长性能和营养成分表观消化率的影响[J].动物营养学报,2013,25(2):433-440.
[13] 王加启,冯仰廉.不同粗饲料日粮发酵规律及合成瘤胃微生物蛋白质效率研究[J].黄牛杂志,1994,20(增刊):82-87.
[14] 周汉林,莫放,李琼,等.日粮中性洗涤纤维水平对中国荷斯坦公牛营养物质消化率的影响[J].海南大学学报:自然科学版,2006,24(3):276-281.
[15] GRANT R J,MERTENS D R.Influence of buffer pH and raw com starch addition on in vitro fiber digestion kinetics[J].Journal of Dairy Science,1992,75(10):2762-2768.
[16] 禹爱兵,范忠军,周永康,等.不同碳水化合物结构组成日粮在徐淮白山羊消化道内降解利用的研究[J].安徽农业科学,2012,40(12):7157-7160.
[17] BECKMAN J L,WEISS W P.Nutrient digestibility of diets with different fiber to starch ratios when fed to lactating dairy cows[J].Journal of Dairy Science,2005,88(3):1015-1023.
[18] 霍鲜鲜,侯先志,赵志恭.日粮不同碳水化合物比例对绵羊瘤胃内纤维物质降解率的影响[J].甘肃畜牧兽医,2004(1):6-8.
[19] 谭支良.绵羊日粮中不同碳水化合物和氮源比例对纤维物质消化动力学的影响及其组合效应评估模型研究[D].博士学位论文.呼和浩特:内蒙古农业大学,1998:42-46.
[20] 冯仰廉.反刍动物营养学[M].北京:科学出版社,2004:235-238.
[21] REYNOLDSC K,TYRRELL H F,REYNOLDS P J.Effects of diet forage-to-concentrate ratio and intake on energy metabolism in growing beef heifers:whole body energy and nitrogen balance and visceral heat production[J].The Journal of Nutrition,1991,121(7):994-1003.
[22] DEVANT M,FERRET A,GASA J,et al.Effects of protein concentration and degradability on performance,ruminal fermentation,and nitrogen metabolism in rapidly growing heifers fed high concentrate diets from 100 to 230 kg body weight[J].Journal of Animal Science,2000,78(6):1667-1676.
[23] 黄洁,申跃宇,姜军,等.日粮蛋白质进食量对杂交育成母牛氮代谢的影响[J].中国畜牧杂志,2012,48(11):52-55.
[24] 杨膺白,梁贤威,郭辉,等.山羊尿中嘌呤衍生物排出规律的研究[J].黑龙江畜牧兽医,2011(1):64-66.
[25] JETANA T,ABDULLAH N,HALIM R A,et al.Effects of energy and protein supplementation on microbial-N synthesis and Allantoin excretion in sheep fed guinea grass[J].Animal Feed Science and Technology,2000,84(3/4):167-181.
[26] 孔祥浩,贾志海,等.后肠道对反刍动物氮代谢的作用[J].饲料工业,2004,25(5):4-6.
[27] 冯仰廉.反刍动物营养学[M].北京:科学出版社,2004.
[28] 王新谋.家畜粪便学[M].上海:上海交通大学出版,1997.
[29] 杨凤.动物营养学[M].北京:中国农业出版社,2006.
[30] TYRRELL H,MOE R P W.Effect of intake on digestive efficiency[J].Journal of Dairy Science,1975,58(8):1151-1163.
[31] 许贵善,刁其玉,纪守坤,等.不同饲喂水平对肉用绵羊能量与蛋白质消化代谢的影响[J].中国畜牧杂志,2012,48(17):40-44.
[32] HOLTER J B,YOUNG A J.Nutrition,feeding and calves:methane prediction in dry and lactating Holstein cows[J].Journal of Dairy Science,1992,75:2165-2175.
[33] HOOVER W H,STOKES S R.Balancing carbohydrates and proteins for optimum rumen microbial yield[J].Journal of Dairy Science,1991,74(10):3630-3644.
[34] BEAUCHEMIN K A,MCGINN S M.Methane emis-sions from feedlot cattle fed barley or corn diets[J].Journal of Animal Science,2005,83(3):653-661.
[35] 王吉峰.日粮精粗比对奶牛消化代谢及乳脂肪酸成分影响的研究[D].硕士学位论文.北京:中国农业科学院,2004.
[36] 刘洁.肉用绵羊饲料代谢能与代谢蛋白质预测模型的研究[D].博士学位论文.北京:中国农业科学院,2012.
猜你喜欢
吉林畜牧兽医(2022年6期)2022-11-16
情感读本·道德篇(2022年2期)2022-04-12
三农资讯半月报(2020年16期)2020-09-14
科学(2020年2期)2020-08-24
英语文摘(2019年9期)2019-11-26
湖南饲料(2019年5期)2019-10-15
湖南畜牧兽医(2016年3期)2016-06-05
特产研究(2016年3期)2016-04-12
海峡姐妹(2016年6期)2016-02-27
诗选刊(2015年6期)2015-10-26
