
桔小实蝇卵巢形态结构及发育特征研究
2014-11-25 02:58季清娥黎剑平
环境昆虫学报 2014年1期
陈 敏,陈 鹏,叶 辉,季清娥,黎剑平
(1.云南大学跨境生态与生态安全实验室,昆明 650091;2.云南省林业科学院,昆明 650201;3.西南林业大学生命科学学院,昆明 650224;4.福建农林大学植物保护学院,福州 350002;5.西南林业大学林学院,昆明 650224)
桔小实蝇Bactrocera dorsalis (Hendel),隶属双翅目 Diptera、实蝇科 Tephritidae、果实蝇属Bactrocera。桔小实蝇寄主广泛,可危害40 余科植物的300 余种瓜果;繁殖能力强,每雌产卵最高可达上千枚;危害严重,可取食芒果、柑桔等具有重要经济价值的热带亚热带瓜果,在防控不当情况下可导致大量减产,造成重大经济损失(Smith,1989;李红旭和叶辉,2000;Clarke et al.,2005;黄素青和韩日畴,2005)。一直以来,桔小实蝇被世界各国作为主要瓜果害虫或列为重要检疫虫害(张润杰和侯柏华,2005)
桔小实蝇繁殖生物学是开展桔小实蝇监测防控的知识基础,是阐释桔小实蝇繁殖行为的主要依据。桔小实蝇雌成虫行为及相关繁殖活动受卵巢发育进度调控,在卵巢发育成熟前主要表现为搜寻食物和补充营养,卵巢成熟后转入搜寻繁殖寄主,进行产卵繁殖(Siderhurst and Jang,2006;2010)。卵巢发育状况决定了桔小实蝇以取食为主抑或产卵为主的行为特征(Avarre et al.,2007)。
实蝇类昆虫的卵巢发育进度可以通过卵巢的形态结构特征加以反映,卵巢形态特征也由此被作为直观判别卵巢发育进度的客观依据(Kendra et al.,2006)。作为桔小实蝇繁殖生物学研究的一部分,本研究将揭示桔小实蝇卵巢形态结构及其发育特征,阐释桔小实蝇卵巢发育与其形态结构联动变化的内在关联,并从桔小实蝇预测监控出发,探索对桔小实蝇卵巢发育进程加以特征分级。本研究将为认识桔小实蝇卵巢形态结构及其发育特征提供了新的知识,为利用桔小实蝇卵巢特征开展桔小实蝇高效监测预报防控提供依据。
1 材料与方法
1.1 供试虫源
本研究在云南大学跨境生态与生态安全实验室进行。供试桔小实蝇源于福建农林大学益虫研究所,在本实验室已经两代繁殖。
1.2 试虫饲养
饲养桔小实蝇的培养箱为LRH-250-GS(广东省医疗器械厂和广东省环保仪器厂合作生产),实验条件设置为26℃温度,L ∶D=12 h∶12 h光照时间,60%相对湿度。供试用桔小实蝇雌成虫均为同日羽化,放入培养箱中养虫笼(35 cm× 30 cm× 30 cm)内,饲以糖水(1 ∶3)和酵母液(1 ∶3)。糖水和酵母液浸于棉球中,置于养虫笼中培养皿内,每隔12 小时更换一次。桔小实蝇产卵期间,在养虫笼放置带皮香蕉供桔小实蝇产卵。
1.3 卵巢观察
从雌性成虫羽化之日起,逐日取雌成虫20 头进行卵巢解剖,观察记录卵巢形态特征及发育进度。卵巢解剖在体视显微镜(PXS-2040)下进行。卵巢解剖步骤是,先在洁净载玻片凹处滴一滴Ringer's 生理盐水,用解剖剪取下桔小实蝇腹部置载玻片凹处,用解剖针去除腹部背板和腹板,用镊子取出桔小实蝇卵巢,剔除卵巢周围的脂肪等,用Ringer's 生理盐水清洗卵巢1-2 次。利用AUTO-MO1VFAGE 景深共聚焦重构分析系统获取卵巢形态特征照片,记录卵巢长宽、卵巢小管长宽和数量、卵粒长宽和数目等相关参数。
2 结果与分析
2.1 桔小实蝇雌性生殖系统结构
桔小实蝇的雌性生殖器官主要由卵巢、输卵管及附腺和受精囊等组成(图1)。卵巢有两组,每组卵巢包裹于透明卵巢膜内,整体上形似椭圆状,羽化初期为淡黄色,后期呈乳白色。每组卵巢含28个卵巢管。在发育成熟的卵巢管中,其端部可见无色的卵原细胞,及经过分裂后产生的卵母细胞和营养细胞,在卵巢管基部常见一枚成熟卵。卵巢基部与侧输卵管相接,两侧输卵管再与中输卵管汇合(图1)。受精囊呈圆球状,通过细长的受精囊管与中输卵管连结(图1)。雌性生殖器官含附腺1 对,为无色透明囊状,基部与中输卵管相连(图1)。
2.2 卵巢发育特征
逐日观察发现,卵巢在雌成虫羽化第1 d 较小,呈淡黄色球状,随时间推移逐渐增大,至第7 d增大明显,呈乳白色椭圆状,羽化第10 d 左右卵巢体积达到最大,约为1.78 mm×1.50 mm。进入产卵末期,卵巢开始逐渐变小,最小时约为0.94 mm×0.85 mm。
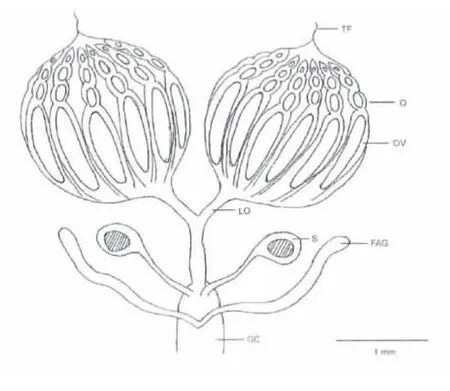
图1 桔小实蝇雌性生殖器官示意Fig.1 Ovaries of Bactrocera dorsalis female
新羽化雌成虫卵巢内似絮状物,无卵巢管,第4 d 后可见少量卵巢管,呈乳白色。卵巢管分化发育从两组卵巢相向一侧开始,随时间推移卵巢管逐渐增多,在羽化后第10 d 卵巢内各卵巢管全部发育成熟,每组卵巢内卵巢管数量为28 根。
随卵巢发育进程,各卵巢管也逐渐增长,成熟卵巢管长度约为1.50 mm,直径约为0.20 mm。进入产卵末期,卵巢管数量逐渐减少,长度逐渐缩短。在产卵末期,卵巢管渐次消解,卵巢相应萎缩。
雌成虫羽化后第4 d,先期形成的卵巢管内可见卵粒,其呈乳白色。观察发现,卵的发育与卵巢管的发育大体同步,卵母细胞和渐成熟卵粒随卵巢管发育而逐渐显现。卵巢管发育成熟后,其内均具有一枚发育近成熟的卵。观察还发现,后期形成的卵巢管较先期形成的卵巢管在发育进程上要快得多。羽化第10 d 后,各卵巢管内均可见1枚近成熟卵和2-3 枚待成熟卵,卵巢内总计可有56 枚近成熟卵。卵巢管内卵粒大小为1.05 mm×0.20 mm。桔小实蝇开始产卵时,卵巢内所有卵巢管均基本发育成熟。
2.3 卵巢分级
卵巢分级是根据卵巢发育进度基于卵巢形态特征对卵巢发育进行的一种划分方式,以便掌握和辨识卵巢发育进度,并根据卵巢发育进度开展相关科学研究和实蝇种群预测 (李汝铎等,1987)。根据桔小实蝇卵巢发育过程中卵巢的形状、大小、颜色、卵巢小管的形成、卵粒的有无等特征,本研究将卵巢发育阶段分为5 级,即发育初期(Ⅰ级)、卵黄沉积期(Ⅱ级)、成熟待产期(Ⅲ级)、产卵盛期(Ⅳ级)和产卵末期(Ⅴ级)。
Ⅰ级定为卵巢发育初期,主要特征是卵巢体积大小为0.53 mm×0.51 mm 左右,卵巢内呈乳白色絮状,卵巢表面附着大量白色端丝。卵巢管未分化。
Ⅱ级为卵黄沉积期,在该时期卵巢体积明显增大,略为1.26 mm×1.11mm 左右,呈淡黄色半透明状。卵巢管开始分化,每组卵巢内可见4-6根卵巢管,其呈淡黄色或转为乳白色,长度约1.24 mm 。卵巢内可见4-6 枚乳白色近成熟卵粒。
Ⅲ级为成熟待产期,识别特征是卵巢体积达到最大,为1.78 mm×1.50 mm 左右,呈乳白色椭圆球状;卵室内充满乳白色近成熟卵粒,排列整齐。
Ⅳ级为产卵盛期,主要表现为卵巢体积较小,大小1.46 mm×1.04 mm 左右,呈乳白色椭球状;卵巢内可见8-10 粒乳白色成熟卵粒。
Ⅴ级为产卵末期,主要特征是卵巢体积小,大小0.94 mm×0.85 mm 左右,呈淡黄色球状;卵巢表面附着大量白色端丝;卵巢管消失;卵巢内已无白色卵粒。
3 结论与讨论
卵巢是实蝇产卵繁殖的重要器官,本研究结果表明:桔小实蝇卵巢由两组卵巢管组成,每组卵巢含卵巢管28 根。新羽化桔小实蝇卵巢处于发育初期,无卵巢管。羽化后第4 d,卵巢管开始显现,卵巢管数量增加,至第10 d 左右,卵巢发育成熟。进入产卵末期,卵巢管逐渐消解,卵巢变小萎缩。桔小实蝇卵巢发育进程分为5个发育等级,即发育初期(Ⅰ级)、卵黄沉积期(Ⅱ级)、成熟待产期(Ⅲ级)、产卵盛期(Ⅳ级)和产卵末期(Ⅴ级)。
在实蝇的繁殖生物学研究中,卵巢管的结构及数量与实蝇的繁殖行为及繁殖力密切关联(Wayne et al.,1997)。桔小实蝇繁殖能力强、产卵量大,每雌平均产卵1200-1500 枚,最高可产卵1800 枚 (Ye,2001;谢琦和张润杰,2005)。本研究揭示,桔小实蝇卵巢分为两组,每组卵巢含卵巢管28个(图1)。在桔小实蝇性发育成熟期,同一卵巢管内可见有1 枚成熟及2-3 枚待成熟卵。从卵巢管结构和数量推论,桔小实蝇在正常情况下可一次性产卵56 枚以上,一生产卵至少可达224 枚。如果桔小实蝇每雌可产卵1000 余枚,则每卵巢管平均可孕育近20 枚卵。新羽化桔小实蝇需取食发育直到性成熟后才开始产卵繁殖(Siderhurst and Jang,2010)。本研究基于卵巢器官解剖观察发现,新羽化桔小实蝇卵巢仅为一雏形,需10 d 左右才发育成熟。在此期间,卵巢发育经历了卵巢管形成、卵母细胞和营养细胞发育、卵形成发育和成熟等过程。此外,随桔小实蝇产卵结束,卵巢管将随之消解,卵巢也逐渐萎缩(图2)。本研究从组织发生学上,阐释了桔小实蝇卵巢生长发育的主要特征和基本规律,揭示了新羽化桔小实蝇在产卵前期表现出积极食物搜寻行为的内在原因(图2)。
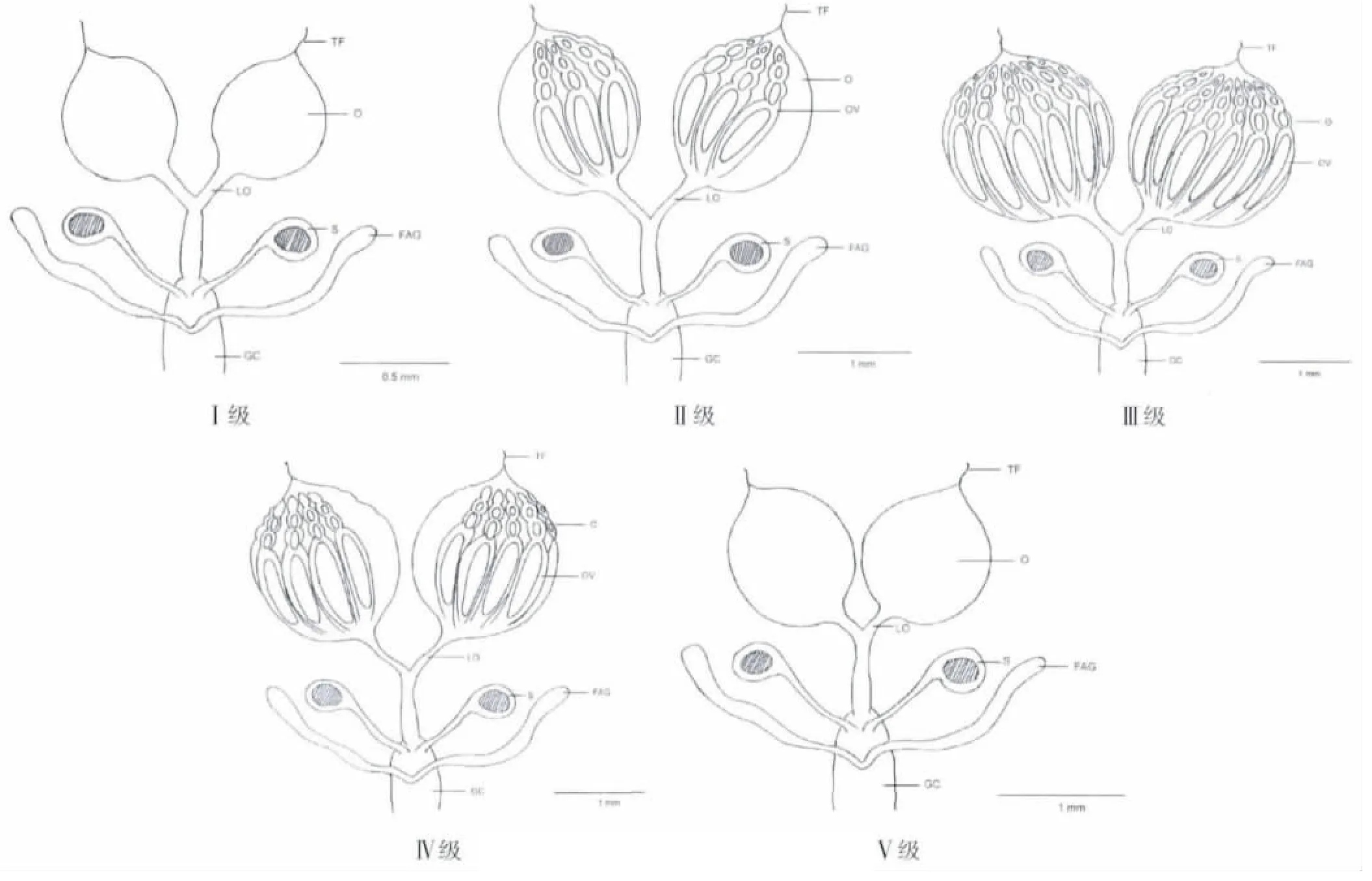
图2 桔小实蝇卵巢发育分级图Fig.2 Stages of ovarian development in Bactrocera dorsalis female
桔小实蝇卵巢管分化及发育进度因其所在卵巢中的位置而异。同时,在卵巢管的形成过程中,卵的发育也随之进行(图2)。该现象在其它昆虫中也有发现(叶辉,1996)。研究还发现,分化晚的卵巢管在发育进度上较分化早的要快,同时,尽管先期分化发育的卵巢管及所孕育的卵看似已经发育成熟,但桔小实蝇仍仅在所有卵巢管发育成熟后才开始产卵。桔小实蝇卵巢及卵发育的这一特征在过去文献报道中尚不多见,它解释了桔小实蝇集团产卵的物质基础(袁盛勇等,2003),但其适应性进化的逻辑基础及内在的调控机制仍不清楚。
实蝇性成熟进度因温度不同而表现出明显差异(Liu and Ye,2009;余丽萍和徐南昌,2012)。番石榴实蝇产卵前期在24℃温度下为38.75 d,在30℃温度下为23.25 d,在33℃温度下为20.50 d(Liu and Ye,2009)。瓜实蝇在20℃温度产卵前期下为36.43 d,在25℃温度下为23.06 d,在30℃温度下则为15.90 d (韦淑丹等,2011)。桔小实蝇在12℃温度产卵前期为48 d,在22℃温度产卵前期为18d,在28℃温度下为10 d (余丽萍和徐南昌,2012)。本研究中,桔小实蝇在26℃温度下的产卵前期为10 d。上述说明,随环境温度变化,桔小实蝇卵巢发育进度将随之发生相应改变。
实蝇性成熟期还与其所获得的营养有关。蛋白质是影响实蝇生殖系统发育的重要营养源,糖是维持生命基本活动的重要营养源 (王波等,2012)。Carey et al.(1998)认为,实蝇取食蛋白更容易释放荷尔蒙有助于提高交配成功率,当实蝇由取食糖向取食蛋白转变时,其行为则由觅食为主转到以产卵为主。王波等(2012)报道,桔小实蝇在仅饲以糖的情况下,性发育受阻,不能正常产卵。在本实验26℃温度下饲以糖水和酵母液,新羽化桔小实蝇经10 d 可达到性成熟。
本研究基于卵巢形态学特征对桔小实蝇卵巢发育进度进行了分级,旨在从卵巢形态特征上反映卵巢的发育进度,为桔小实蝇科学防控提供依据。通常类似的分级往往分为4 级(Chou et al.,2012)。本研究观察表明,桔小实蝇在产卵结束之后仍能存活一段时间,其卵巢显著萎缩,故将其列为第5 级,以便完整反映卵巢从最初形成到最终萎缩的全过程。
References)
Avarre JC,Lubzens E,Babin PJ.Apolipocrustacein,formerly vitellogenin,is the major egg yolk precursor protein in decapod crustaceans and is homologous to insect apolipophorin II/I and vertebrate apolipoprotein B.[J].BMC Evolutionary Biology,2007,7 (1):3.
Carey JR,Liedo P,Müller HG,et al.Dual modes of aging in mediterranean fruit fly females [J].Science,1998,281 (5379):996-998.
Chou MY,Mau RFL,Jang EB,et al.Morphological features of the ovaries during oogenesis of the oriental fruit fly,Bactrocera dorsalis,in relation to the physiological state [J].Journal of Insect Science,2012,12 (144):1-12.
Clarke AR,Armstrong KE,Carmichael AE,et al.Invasive phytophagous pests arising through a recent tropical evolutionary radiation:The Bactrocera dorsalis complex of fruit flies [J].Annual Review of Entomology,2005,50:293-319.
Huang SQ,Han RC.Advance in the research on the quarantine pest Bactrocera dorsalis [J].Chinese Bulletin of Entomology,2005,42(5):479-484.[黄素青,韩日畴.桔小实蝇的研究进展[J].昆虫知识,2005,42 (5):479-484]
Li HX,Ye H,Lu J.On damages and distributions of fruit fly(Bactrocera dorsalis Hendel)in Yunnan Province,Southern China[J].Journal of Yunnan University (Natural Sciences Edition),2000,22 (6):473-475.[李红旭,叶辉.桔小实蝇在云南的危害与分布[J].云南大学学报,2000,22 (6):473-475]
Liu XF,Ye H.Effect of temperature on development and survival of Bactrocera correcta (Diptera:Tephritidae)[J].Scientific Research and Essay,2009,4 (5):467-472.
Siderhurst MS,Jang EB.Cucumber volatile blend attractive to female melon fly,Bactrocera cucurbitae (Coquillett)[J].Journal of Chemical Ecology,2010,36:699-708.
Smith PH.Behavioral partitioning of the day and circadian rhythmicity.In:Robinson AS,Hooper G,eds.Fruit Flies:Their Biology,Natural Enemies and Control.World Crop Pests Series Vo1.3B.Amsterdam:Elsevier.1989.325-341.
Wang B,Ji QE,Chen JH,Han Y,et al.The fecundity and life table of experimental population of B.dorsalis under different nutrition condition [J].Chinese Journal of Tropical Crops,2012,33 (3):551-555.[王波,季清娥,陈家骅,等.不同营养条件对桔小实蝇实验种群生殖力及生命表的影响[J].热带作物学报,2012,33 (3):551-555]
Wayne ML,Hackett JB,Mackay TFC.Ouantitative genetics of ovarioIe number in Drosophila melanogaster I .Segregating variation and fitness [J].Euolution,1997,51:1156-1163.
Wei SD,Huang SS,Wang YQ,et al.Effect of temperature on the development and reproduction of Bactrocera cucurbitae (Coquillett)population [J].Guangxi Agricultural Sciences,2011,42 (7):744-747.[韦淑丹,黄树生,王玉群,等.温度对瓜实蝇实验种群生长发育及生殖的影响研究[J].南方农业学报,2011,42 (7):744-747]
Xie Q,Zhang RJ.Study advance on biology and ecology of Bactrocera dorsalis (Hendel)and its control [J].Ecological Science,2005,24 (1):52-56.[谢琦,张润杰.桔小实蝇生物学特点及其防治研究概述[J].生态科学,2005,24 (1):52-56]
Yu LP,Xu NC.Relationship with the biological characteristics of Bactrocera dorsalis and temperature [J].China Garden Digest,2012,(5):19-21.[余丽萍,徐南昌.柑桔小实蝇生物学特性与温度的关系[J].中国园艺文摘,2012,(5):19-21]
Yuan SY,Xiao C,Kong Q,et al.Oviposition prefernce of Bactrocera dorsalis Hendel [J].Acta Agriculturae Universitatis Jiangxiensis(Natural Sciences Edition),2005,27 (1):81-84.[袁盛勇,肖春,孔琼,等.桔小实蝇的产卵选择性[J].江西农业大学学报,2005,27 (1):81-84]
Yuan SY,Xiao C,Li ZQ,et al.A Study on laboratory rearing techniques for Bactrocera dorsalis (Hendal)[J].Acta Agriculturae Universitatis Jiangxiensis (Natural Sciences Edition),2003,25(4):577-580.[袁盛勇,肖春,李正跃,等.桔小实蝇实验室饲养技术研究[J].江西农业大学学报(自然科学版),2003,25 (4):577-580]
Ye H.Distribution of the oriental fruit fly (Diptera:Tephritidae)in Yunnan Province [J].Entmol.Sin.,2001,8 (2):175-182.
Ye H.Studies on the biology of Tomicus piniperda L.(Coleoptera:Scolytidae)in the shoot feeding period [J].Acta Entomologica Sinica,1996,39 (1):58-62.[叶辉.纵坑切梢小蠹蛀梢期生物学研究[J].昆虫学报,1996,39 (1):58-62]
Zhang RJ,Hou BH.Assessment on the introduction risk of Bactrocera dorsalis (Hendel)through imported fruits with fuzzy mathematics[J].Acta Entomologica Sinica,2005,48 (2):221-226.[张润杰,侯柏华.桔小实蝇传入风险的模糊综合评估[J].昆虫学报,2005,48 (2):221-226]
猜你喜欢
鸭绿江(2022年10期)2022-10-21
飞天(2021年12期)2021-12-24
ELLE世界时装之苑(2020年8期)2020-08-03
湖北植保(2017年4期)2017-08-31
小溪流(画刊)(2017年6期)2017-07-10
文艺生活·下旬刊(2017年2期)2017-03-18
蔬菜(2016年8期)2016-10-10
湖南农业(2016年3期)2016-06-05
湖南农业(2016年3期)2016-06-05
广西蚕业(2015年3期)2015-11-30
