
桔小实蝇Rab 家族基因鉴定和表达模式研究
2014-11-25 02:58郑薇薇吴方玉张宏宇
环境昆虫学报 2014年1期
彭 威,郑薇薇,彭 涛,吴方玉,张宏宇
(华中农业大学植物科学技术学院,昆虫资源利用与害虫可持续治理湖北省重点实验室,城市与园艺昆虫研究所,武汉 430070)
桔小实蝇Bactrocera dorsalis (Hendel)俗称果蛆,又名东方果实蝇(Oriental fruit fly),属双翅目 Diptera、实蝇科 Tephritidae、果实蝇 属Bactrocera Macquart,是一种能导致果蔬业、花卉业严重经济损失的毁灭性害虫,在我国尤以对番石榴、芒果、柑桔、沙田柚等危害最重(韩日畴等,2005),由于其食性杂、寄主范围广、繁殖力强、传播快、防治困难,被大多数国家和地区列为水果―头号杀手(季清娥等,2007)。Rab 蛋白为一类进化上保守的GTP 结合蛋白,是质膜运输的重要调控因子(Salminen et al.,1987),属GTP结合蛋白Ras 超家族的成员(Wennerberg et al.,2005)。Rab 蛋白通常作为活性和非活性状态循环的分子开关,以时空灵敏的方式整合质膜运输和细胞间的信号传导(Bucci et al.,2006)。尽管这类蛋白分子量小,只有20-25 kDa,结构分析显示它们具有多个反应表面,通过这些反应表面参与调节分子和下游效应物,从而发挥功能(Chen et al.,2003)。Rab 蛋白广泛参与了不同的细胞功能,包括质膜融合和分裂、细胞内吞作用、胞吐作用、囊泡转运、细胞骨架运输。Rab 蛋白作为囊泡运输的重要调节因子,在囊泡的出泡形成、选择、粘附、运动、锚定、融合等过程中起重要作用(Pfeffer et al.,2004)。目前发现,在裂殖酵母中含有7个Rab 蛋白,芽殖酵母编码11个Rab 蛋白,线虫、果蝇分别有29 种和33 种Rab 蛋白,拟南芥基因组编码57 种Rab 蛋白,在人类Homo sapiens 已发现超过70 种Rab 蛋白和类Rab 蛋白。大约一半已知的Rab 蛋白研究表明,Rab 蛋白对信号传递,以及对细胞增殖和分化的控制很重要(Schwartz et al.,2008)。随着研究的深入,人们发现Rab 蛋白在发育过程中调节信号传导。早期胚胎细胞的命运受分泌信号蛋白如Hh、Wnt、TGFβ/Dpp的控制,信号浓度在空间和时间上的控制对正常发育来说至关重要,而受Rab 调控的胞内运输调控着信号梯度和转导(Seto et al.,2008)。此外,Rab 基因突变会影响细胞生长、寿命和其他生物学进程(Schwartz et al.,2008)。
随着对Rab 蛋白研究的深入,关于Rab 蛋白在昆虫上的研究也有诸多报道。Uno 等对家蚕的Rab 蛋白进行研究发现,Rab 蛋白会被家蚕脑部的蛋白激酶C 磷酸化 (Uno et al.,2004)。而Hiragaki 等人(2009)发现,磷酸化的Rab8 蛋白参与了昆虫促前胸腺激素的分泌。对家蚕脑部的小分子GTP 结合蛋白Rab的研究发现,Rab7、Rab8 参与了家蚕脑部神经元中的蛋白运输,而且它们可能在控制生理节律方面发挥了作用(Uno et al.,2010)。侯丽等人在对棉铃虫幼虫到成虫的发育研究中发现,Rab32 参与了成虫肠道的重建(Hou et al.,2011)。除此之外,目前,Rab 在昆虫发育中的作用研究主要集中在果蝇。例如,在果蝇卵母细胞的发育过程中,Rab6 对微管细胞骨架的组织和定位是必须的(Coutelis et al.,2007)。而Rab30 蛋白在果蝇胚胎发育中的背部闭合、胚胎头部对合、胸部闭合等过程是必须的(Thomas et al.,2009)。张军等人对果蝇的Rab35 蛋白研究发现,Rab35 蛋白调节刚毛发育过程肌动蛋白纤丝的聚集,以及可培养细胞伪足的形成 (Zhang et al.,2010)。由此可见,Rab 家族基因参与了果蝇诸多重要的生命活动。基因表达是生物体中遗传信息传递和实现的中间环节,而基因表达模式是基因表达调控的体现和结果,因此对全基因组基因表达模式的研究有助于揭示基因表达途径及其调控规律,同时揭示生物体基因型和表型之间的特定关系(谢建明等,2003)。对桔小实蝇7个Rab 家族成员进行鉴定并研究它们的基因表达模式能为揭示桔小实蝇Rab 家族基因功能提供基础资料,同时对于筛选合适的基因功能研究靶标基因具有重要意义。
1 材料与方法
1.1 供试昆虫
本研究中桔小实蝇来自华中农业大学城市与园艺昆虫研究所建立的实验室种群。饲养条件如下:温度28℃,室内的相对湿度保持在70%-80%,光周期设置为12 h ∶12 h (L ∶D)(Li et al.,2011)。
1.2 主要试剂及试剂盒
主要试剂、试剂盒:Gibco/BRL 公司(美国)的琼脂糖、Sigma 公司(美国)的焦磷酸二乙酯(DEPC)、北京博迈德生物的2× Tag PCR Master Mix、上海申能博彩公司的Taq DNA 聚合酶、北京百泰克生物技术有限公司的高纯总RNA 快速提取试剂盒、TaKaRa 公司(日本)的RT-PCR 第一链合成试剂盒、Bio-Rad 公司(美国)的iQTM SYBR ⒸGreen Supermix 试剂盒。
1.3 生物信息学和系统进化树分析
从转录组文库中获取得到桔小实蝇7个Rab基因的核苷酸序列信息(Zheng et al.,2012)。用DNAman 完成核苷酸序列编辑及氨基酸序列推导。通过美国国家生物技术信息中心(NCBI)站点的BLASTX (http://www.nebi.nlm.nih.gov)查找各Rab 基因的高相似性序列。用ClustalX 2.0 对桔小实蝇Rab 家族基因氨基酸序列进行多重比对,分析其保守性,揭示其活性位点。用MEGA5.0 软件neighbor-joining 方法构建Rab 蛋白进化树,根据bootstrap 方法用1000个重复进行统计分析。
1.4 桔小实蝇各发育阶段总RNA的提取
取桔小实蝇二龄幼虫、三龄幼虫、预蛹、蛹、成虫各10 头,分别提取各组整虫总RNA。总RNA提取采用Bioteke 公司的高纯总RNA 快速提取试剂盒(北京),具体操作参照试剂盒说明书。
1.5 1st-Strand cDNA的合成、cDNA 质量检测
将各组样本的总RNA 反转录为cDNA,具体操作参照TaKaRa 公司(日本)的RT-PCR 第一链合成试剂盒说明书。
取各样本cDNA为模板,以桔小实蝇核糖体16S rRNA 设计引物,进行PCR 扩增,然后琼脂糖凝胶检测产物片段大小。
1.6 实时荧光定量PCR
根据转录组测序信息设计Rab 家族基因的荧光引物,桔小实蝇16SrRNA 作为内参基因。实验中用到的引物名称、碱基序列 (5' →3'):Rab1RTF (GCAGCGGAATACGCAAGTC)、Rab1RTR(TATCTCGGCAGCCATCGTC);Rab5RTF (AGCT CCTATGTACTATCGTGGTG)、Rab5RTR (TTTATG CAGTTCCTTGACCC);Rab7RTF (ACTTTCCTTTCG TGGTGCTCG)、Rab7RTR (GTACAGTTCAACCTCG GCTTC);Rab14RTF (TACTATCGTGGTGCTGCTG GTG)、Rab14RTR (GCAAACTCTTTGGCCTCTTCA);Rab18RTF (AAGTCAAGCCTCATTCGTCG)、Rab18 RTR (CTTCGGAATCTTTCAGCACC);Rab35RTF(G TAGACTTCAAGATACGCACCG)、Rab35RTR (CTA ACCATCGTCGTACATTCG);Rab40RTF (ACATGA GATATTATCCTGCCTTGA)、Rab40RTR (GCTTGA CACGTTTCCCTTCC);16s rRNARTF (CTCGTCCA ACCGTTCATACC)、16s rRNARTR (CTGACCTGCC CACTGAAGTT)。
实时荧光定量PCR 用iQTM SYBR ⒸGreen Supermix (Bio-Rad,USA)试剂盒在Bio-RadiQ5 双色实时荧光定量PCR 仪上进行。每个样品设三个重复。荧光定量PCR 结束后分析溶解曲线,以确保特异性扩增。实时荧光定量PCR 结果由Opticon Monitor 软 件2.03Version (MJ research,USA)量值2-ΔCt方法分析,以16S rRNA 表达量校正目的基因的相对表达量。
2 结果与分析
2.1 桔小实蝇Rab 基因生物信息学分析
我们从桔小实蝇转录组文库中鉴定得到了7个Rab 基因全长 cDNA 序列 (Rab1 Genbank accession number:KF 859977;Rab5 Genbank accession number:KF 859978;Rab7 Genbank accession number:KF 859979;Rab14 Genbank accession number:KF 859980;Rab18 Genbank accession number:KF 859981;Rab35 Genbank accession number:KF 859982;Rab40 Genbank accession number:KF859983)。
通过NCBI的BLASTX 查找到与桔小实蝇7个Rab 基因的氨基酸序列对应的相似序列,相似性比对结果如表2 所示。结果显示,除BdRab18 氨基酸序列与刺舌蝇Glossina morsitans Rab18的氨基酸序列相似度达到85%外,桔小实蝇其余Rab 基因的氨基酸序列均与黑腹果蝇D.melanogaster 所对应的Rab 基因的氨基酸序列相似度最高,相似度最高达到96%,最低也到达85%。
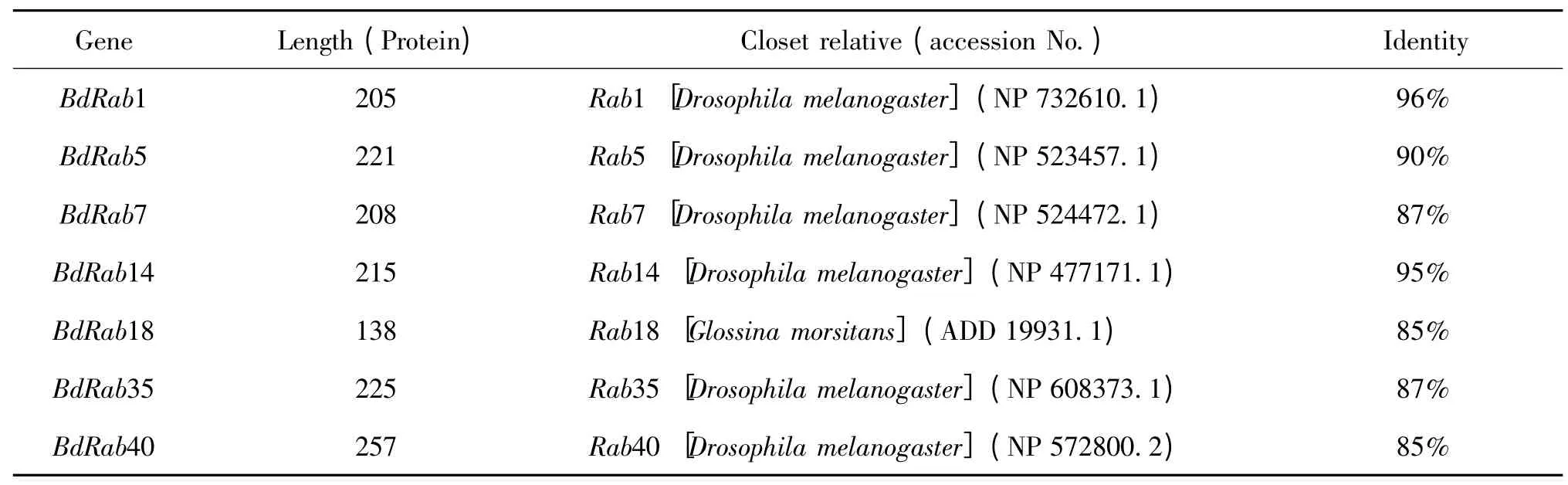
表1 桔小实蝇7个Rab 基因的氨基酸序列同源比对结果Table 1 Identity of amino acid sequence for Bactrocera dorsalis Rab genes
对桔小实蝇7个Rab 基因氨基酸序列进行比对分析,结果如图1 所示。氨基酸序列比对显示这7个Rab 氨基酸序列存在部分保守序列。而这些区域构成GTP/Mg2+结合位点和Rab 蛋白分子特征序列。其中,68 位F 和147-151 位GNKCD 构成GTP 结合位点,38-44 位GDSGVGK 构成Mg2+结合位点,64-68 位IGVDF、82-86KLQTW、94-97 位RFRSIT、100-104 位YYRGA、109-114位LVYDIT 均是Rab 蛋白特征序列。
2.2 桔小实蝇Rab 基因系统发育分析
对桔小实蝇7个Rab 基因构建系统发育树,结果如图2 所示。

图1 桔小实蝇7个Rab 基因氨基酸序列多重比对结果Fig.1 Multiple alignment of amino acid sequences of 7 Rab genes for Bactrocera dorsalis
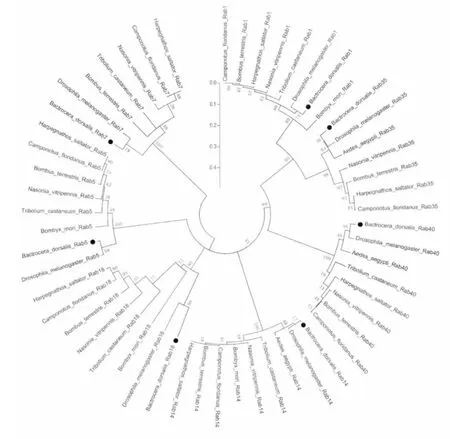
图2 桔小实蝇7个Rab 基因的系统进化树Fig.2 Phylogenetic analyses of 7 BdRab genes with other similar genes
桔小实蝇7个Rab 基因属于不同的同源蛋白。结果显示桔小实蝇7个Rab 基因与黑腹果蝇对应的7个Rab 基因进化距离最近。总共分为三大支,BdRab1、BdRab35、BdRab40 同属一支,分别与DmRab1、DmRab35、DmRab40 进化距 离最近。BdRab5、BdRab14、BdRab18 同属一支,分别与DmRab5、DmRab14、DmRab18 进化距离最近。BdRab7 对应DmRab7。
2.3 桔小实蝇Rab 基因表达模式分析
我们通过实时荧光定量PCR 检测了桔小实蝇Rab 基因在不同发育阶段的表达模式(图3)。
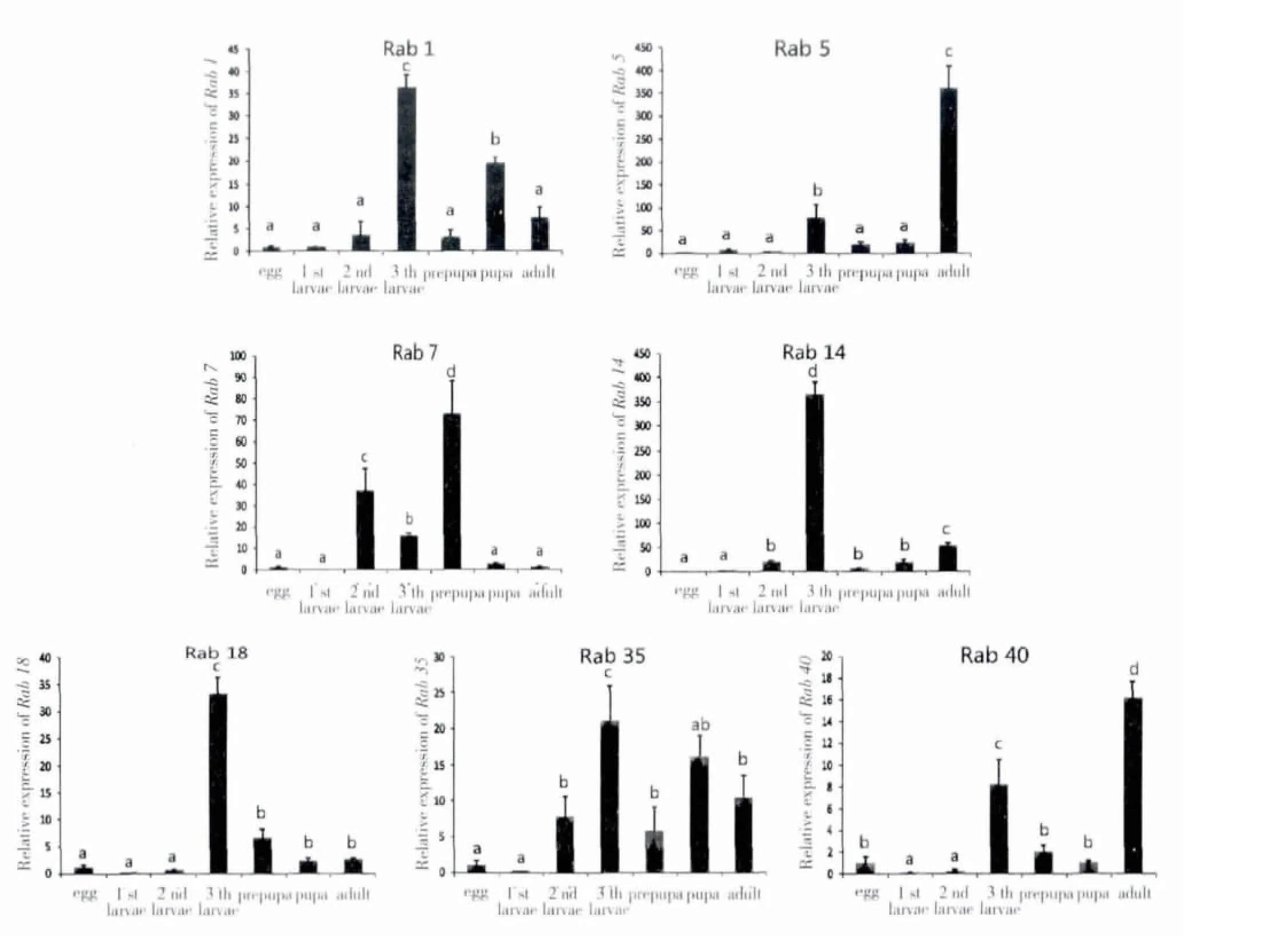
图3 BdRab 在桔小实蝇生命周期各阶段的表达模式Fig.3 The expression pattern of BdRab in the life cycle of Bactrocera dorsalis
由图3 可知,BdRab1 在三龄和蛹期表达量最高,BdRab5 在成虫期高表达,BdRab7 在预蛹期高表达,BdRab14、BdRab18 均在三龄幼虫期表达量显著上调,BdRab35 在三龄和蛹期大量表达,BdRab40 在三龄幼虫和成虫期大量表达,暗示各基因在桔小实蝇不同发育过程中发挥功能。三龄幼虫是化蛹前的准备阶段,完全变态昆虫蛹期进行着剧烈的旧组织解离和新组织的发生,需要大量营养物质和能量,化蛹前昆虫大量取食并迅速生长,形态体积增大,为化蛹做好营养积累的准备。表明BdRab14、BdRab18 可能参与化蛹的准备活动。
3 结论与讨论
我们对桔小实蝇Rab 基因的7个成员进行了生物信息学分析和表达模式研究。对桔小实蝇Rab家族成员基因进行表达模式研究,发现各基因表达模式有差异,暗示各基因在桔小实蝇生长发育过程中发挥不同的功能作用。其中BdRab5 在成虫期显著表达,而其它阶段的表达则处于极低水平,说明其可能参与了成虫期的重要生命活动过程。Seto 等研究发现Rab5 在果蝇翅的发育中通过锚定Wnt 蛋白调节了Wnt 信号通路 (Seto et al.,2006),这一报道则验证了BdRab5 可能参与成虫重要生命活动的假设。小分子GTP 结合蛋白亚家族成员Rab7 在桔小实蝇幼虫二龄蜕皮、三龄变态阶段尤其是预蛹阶段有显著表达,而这些阶段与桔小实蝇幼虫的化蛹发育阶段有时间上重合,Rab7 可能参与到该过程。Rab7 属于小分子GTP 结合蛋白亚家族Rab 家族,该家族成员较多,人类Homo sapiens 身上已发现超过70 种Rab 蛋白和Rab样蛋白,它们在生命活动中扮演多种重要角色,大约一半已知的Rab 蛋白研究表明,Rab 蛋白对信号传递,以及对细胞增殖和分化的控制很重要,Rab 蛋白的突变会影响细胞生长、寿命和其他生物学进程 (Schwartz et al.,2008)。BdRab 14、BdRab 18 均在三龄幼虫期显著表达,在其它阶段表达量都很低,而三龄幼虫是化蛹前的准备阶段,完全变态昆虫蛹期进行着剧烈的旧组织解离和新组织的发生,需要大量营养物质和能量,化蛹前昆虫大量取食并迅速生长,形态体积增大,为化蛹做好营养积累的准备。所以推测三者可能与化蛹的准备活动有关。然而,目前有关Rab 基因在桔小实蝇化蛹准备活动中的具体功能还未见报道,需要进一步的研究。Rab 蛋白是一类小分子GTP结合蛋白,对囊泡运输和信号传递十分重要,对Rab 基因在桔小实蝇变态发育中表达模式的研究,不仅有助于阐明桔小实蝇变态发育的分子机制,而且可为桔小实蝇防治提供新思路和途径。
References)
Bucci C,Chiariello M.Signal transduction gRABs attention [J].Cellular Signalling,2006,18 (1):1-8.
Chen J,Anderson JB,Weese-Scott CD,et al.MMDB:Entrez's 3Dstructure database [J].Nucleic Acids Research,2003,31:474-477.
Coutelis JB,Ephrussi A.Rab6 mediates membrane organization and determinant localization during Drosophila oogenesis [J].Development,2007,134:1419-1430.
Hiragaki S,Uno T,Takeda M.Putative regulatory mechanism of prothoracicotropic hormone (PTTH)secretion in the American cockroach,Periplaneta americana as inferred from co-localization of Rab8,PTTH,and protein kinase C in neurosecretory cells [J].Cell and Tissue Research,2009,335 (3):607-615.
Hou L,Wang JX,Zhao XF.Rab32 and the remodeling of the imaginal midgut in Helicoverpa armigera [J].Amino Acids,2011,40 (3):953-961.
Huang SQ,Han RC.Advance in the research on the quarantine pest Bactrocera dorsalis [J].Chinese Bulletin of Entomology,2005,42(5):479-484.[黄素青,韩日畴.橘小实蝇的研究进展.昆虫知识,2005,42 (5):479-484]
Ji QE,Hou WR,Chen JH.Development of a genetic sexing strain and the sterile male technique of the Oriental fruit fly,Bactrocera dorsalis (Hendel)[J].Acta Entomologica Sinica,2007,50:1002-1008.[季清娥,侯伟荣,陈家骅.橘小实蝇遗传性别品系的建立及雄性不育技术.昆虫学报,2007,50:1002-1008]
Li XX,Zhang MY,Zhang HY.RNA interference of four genes in adult Bactrocera dorsalis by feeding their dsRNAs [J].PLoS One,2011,6,e17788.
Pfeffer S,Aivazian D.Targeting Rab GTPases to distinct membrane compartments [J].Nature Reviews Molecular Cell Biology,2004,11:886-896.
Salminen A,Novick PJ.A ras-like protein is required for a post-Golgi event in yeast secretion [J].Cell,1987,49 (04):527-538.
Schwartz1 SL,Cao CC,Pylypenko O,et al.Rab GTPase at a glance[J].Journal of Cell Science,2008,120:3904-3910.
Seto ES,Bellen HJ.Internalization is required for proper Wingless signaling in Drosophila melanogaster [J].The Journal of Cell Biology,2006,173 (01):95-106.
Thomas C,Rousset R,Noselli S.JNK signalling influences intracellular trafficking during Drosophila morphogenesis through regulation of the novel target gene Rab30 [J].Developmental Biology,2009,331(2):250-260.
Uno T,Hata K,Hiragaki S,Isoyama Y,et al.Small GTPases of the Rab family in the brain of Bombyx mori [J].Histochemistry and Cell Biology,2010,134 (6):615-622.
Uno T,Nakao A,Katsurauma C.Phosphorylation of Rab proteins from the brain of Bombyx mori [J].Archives of Insect Biochemistry and Physiology,2004,57 (2):68-77.
Wennerberg K,Rossman KL,Der CJ.The Ras superfamily at a glance[J].Journal of Cell Science,2005,118:843-846.
Xie JM,Sun X,Xie XY.Gene expression profile analysis and software system [J].Journal of Southeast University (Natural Science Edition),2003,1 (2):201-204.[谢建明,孙啸,谢雪英.基因表达模式分析及软件系统[J].东南大学学报,2003,01(02):201-204]
Zhang JZ,Enbo M,Kun YZ,et al.Silencing of two alternative splicing-derived mRNA variants of chitin synthasel gene by RNAi is lethal to the oriental migratory locust,Locusta migratoria manilensis(Meyen)[J].Insect Biochemistry and Molecular Biology,2010,40 (11):824-833.
Zheng W,Peng T,He W,et al.High-throughput sequencing to reveal genes involved in reproduction and development in Bactrocera dorsalis (Diptera:Tephritidae) [J].PLoS One,2012,7:e36463.
猜你喜欢
玩具世界(2022年3期)2022-09-20
学苑创造·A版(2022年3期)2022-03-29
烟台果树(2021年2期)2021-07-21
甘肃教育(2020年2期)2020-09-11
学苑创造·A版(2019年6期)2019-07-11
测控技术(2018年7期)2018-12-09
小学生优秀作文(低年级)(2018年9期)2018-09-10
小学生必读(低年级版)(2018年11期)2018-03-13
蔬菜(2016年8期)2016-10-10
湖南农业(2016年3期)2016-06-05
