
广州稻区稻螟幼虫种的构成及其空间分布研究
2014-11-25 02:59:28廖永林张振飞肖汉祥李燕芳李怡峰
环境昆虫学报 2014年4期
廖永林,张 扬*,张振飞,肖汉祥,李燕芳,李怡峰
(1.广东省农业科学院植物保护研究所,广州 510640;2.广东省植物保护新技术重点实验室,广州 510640)
水稻螟虫俗称钻蛀性螟虫,主要包括三化螟Tryporyza incertulas、二化螟Chilo suppressalis、台湾稻螟Chilotraea auricilia 和大螟Sesamia inferens 等。它们的为害主要以幼虫钻蛀水稻茎秆,在水稻分蘖期和孕穗期最易发生为害,造成枯心苗、死孕穗、白穗、瘪谷大量增加,严重影响水稻产量(向玉勇等,2011)。长期以来,在我国华南双季稻区以三化螟为主,其它螟虫如二化螟、大螟、台湾稻螟等只是局部轻发生(周圻等,1990)。近年来,在农作物种植调整、耕作制度改变、气候变化、抗药性增强等多种因素综合作用下,广东稻区的钻蛀性螟虫种类、发生规律和为害规律出现了新变化:三化螟仍为我省水稻优势钻蛀性螟虫,二化螟为害明显上升,在粤北和粤东部分地区甚至成为了当地水稻主要钻蛀性螟虫种类;台湾稻螟、大螟在水稻、甘蔗、牧草、玉米等混栽区种群数量上升,成为近年水稻钻蛀性螟虫的新动向(刘光华等,2006;钟宝玉等,2007)。据报道2006年广州稻区螟虫有三化螟、二化螟、大螟和台湾稻螟,其中以台湾稻螟为优势种群,占总虫口数的77.18% (刘光华等,2006)。为掌握广州稻区水稻螟虫的最新为害态势,本研究拟采用聚集强度指数和地统计学方法研究目前该地区水稻螟虫幼虫种的结构及其空间分布型,以期为建立有效的防控措施提供理论依据。
1 材料与方法
1.1 调查方法
于2013年3月至7月,对广东省农科院白云试验基地(广州市白云区钟落潭镇)内未喷施化学农药稻田的钻蛀性螟虫进行调查。调查时采用平行取样法,按横坐标0、5、10、15 m,纵坐标0、5、10、15、20、25、30、35 m 上的32个点进行调查取样,每点随机选择5 丛,分蘖期调查其枯心株数,抽穗期调查其白穗数,并将上述每点的枯心苗及白穗统一用保鲜袋包装好带回室内,剥开稻穗,镜检稻螟幼虫种类并记录各螟虫数量。
1.2 分析方法
1.2.1 聚集强度指数的测定
(1)扩散系数C (丁岩钦,1980):C=S2/m,当C=1 时为随机分布;C >1 时为聚集分布:C<1 时为均匀分布。
(2)根据上述数据,应用SPSS 软件15.0 版本,采用单样本t 检验法对扩散系数C 与1 进行显著性检验。
1.2.2 半方差函数及变异曲线理论模型的拟合
根据区域化变量理论,在空间上昆虫种群数量是区域化变量,因此可用区域化变量理论和方法进行研究。应用区域变量理论,假设在空间有一系列的点(x1,x2,x3,、…)及其取值(Z1,Z2,Z3,…),那么空间所有相距为h的两点的取值两两配对形成一新的集合{[Z (x1),Z (x1+h)],[Z(x2),Z (x2+h)],…},在新集合中对变量Z(xi)和Z (xi+h)按以下公式计算其半方差值:

式中,r (h)称为半方差函数,N (h)为间隔h的样本点的对数。h为变量Z (xi)和Z (xi+h)之间的距离。
半方差函数变异曲线图通常由r (h)对h 作图,这些变异曲线有三个重要参数:块金值(nugget,C0),基台值[sill,(C0+C)]和变程(range,a)(王政权,1999;周国法和徐汝梅,1998)。C0表示随机部分的空间异质性。较大的块金值表明较小尺度上的某种过程不可忽视。与基台值相似,块金值不能用于比较不同变量间的随机性方面的差异。但是块金值与基台值之比C0/ (C0+C)反映块金方差占总空间异质性SH(Z)变化的大小却非常有意义。如果该比值较高说明随机部分引起的空间异质程度SHR较高,相反异质性则由空间自相关引起。如果比值接近1,则景观中某一变量在整个尺度上是具有恒定的变异。变程是r (h)达到基台值时的间隔距离,它表示在h ≥a 以后,区域化变量的空间相关性消失。本文中的变异曲线由Variowin 2.1 软件生成(吕昭智等,2003;李志强等,2008)。在生成曲线图过程中系统对参数进行优化处理,最终获得相应模型和模型参数。常见的模型有球状模型、指数模型、高斯模型、线性有基台值模型和线性无基台值模型等(Rossi et al.,1992;周强等,2001;陈强等,2007)。
1.2.3 地统计学的插值法
克里金法(Kriging)是利用区域化变量的原始数据和变异函数的结构特点,对未采样点的区域化变量的取值进行线性无偏最优估计的一种方法。用克里金法分析估计样区邻近位置的值,这在区域害虫的管理上,对预测和估计种群数量是很有用的方法(Lecoustre et al.,1989;Liebhold et al.,1991)。本文在Sufer 8.0 环境下利用Kriging插值技术生成种群密度等值线图。
2 结果与分析
2.1 螟幼虫种的构成
由表1 可知,广州地区早造分蘖期虫口数小于抽穗期虫口数;分蘖期和抽穗期稻螟幼虫种的构成相一致,均以台湾稻螟为主,占97.01% 以上,二化螟和大螟只是零星发生,约占2.99%,且两个生育期均未见三化螟发生为害。
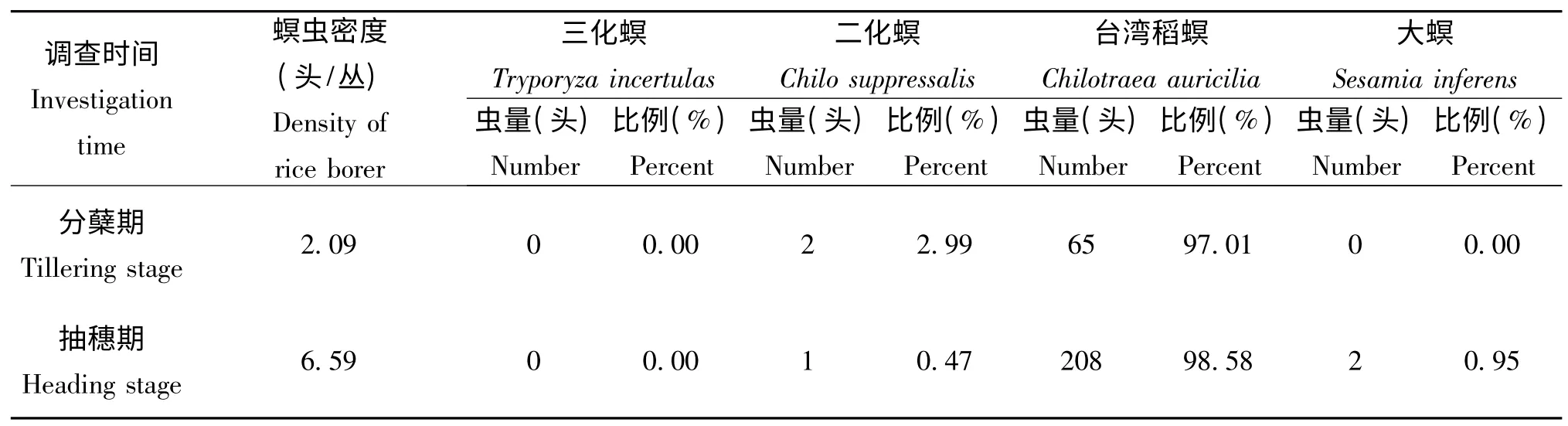
表1 广州稻区稻螟幼虫种的构成Table 1 Species of rice borer larva in paddy field in Guangzhou
2.2 稻螟幼虫聚集强度指数分析
通过对稻螟幼虫扩散系数的分析发现,分蘖期扩散系数除未见稻螟幼虫调查点外,绝大部分大于1;抽穗期扩散系数显著大于1 (表2)。说明稻螟幼虫在分蘖期和抽穗期均呈聚集分布。

表2 稻螟幼虫在田间的聚集强度指数Table 2 Measurement of the indices of aggregation intensity of rice borer larva in paddy field

(续上表)
2.3 稻螟幼虫半变异曲线的模型拟合
根据稻螟幼虫的调查结果,用Variowin 2.1 软件进行地统计学分析,获得相应模型和模型参数见图1 和表3,再对其种群空间结构进行分析。结果表明,分蘖期和抽穗期稻螟幼虫的最优拟合模型均为高斯模型r(h)=c0+c[1-exp(-h2/a2)],h>0,表明分蘖期和抽穗期稻螟幼虫在田间均呈聚集分布。由表3 中变程range (m)可知,分蘖期样点间的空间依赖性距离为18.900 m,而抽穗期样点间的空间依赖性距离为12.647 m。分蘖期稻螟幼虫C0/ (C +C0)比值小于1,说明由空间的自相关因素引起的空间异质性程度较大;而抽穗期稻螟幼虫C0/ (C +C0)比值大于1,说明由随机部分引起的空间变异性程度较大。
2.3 种群密度的等值线图
为更直观的表达稻螟幼虫在大田的空间分布,利用Sufer8.0 软件对稻螟幼虫在大田的分布进行分析,获得相应的密度等值线图。图2 显示,分蘖期和抽穗期稻螟幼虫均有聚集团块,分蘖期聚集团块主要分布在边缘位置,抽穗期则分布在全田,且分蘖期和抽穗期密度等值线均是南侧大于北侧。

表3 空间模型及特征参数Table 3 The spatial models and characteristic parameters

图1 分蘖期和孕穗期稻螟幼虫在稻田中的半变异曲线Fig.1 Semivariograms of rice borer larva in paddy field between tillering and heading stage

图2 稻螟幼虫在稻田中密度等值线图Fig.2 Contourmaps of rice borer larva in paddy field
3 结论与讨论
研究表明,目前,台湾稻螟是广州稻区钻蛀性螟虫的优势种群,此结果与刘光华等2006年对广州稻区钻蛀性螟虫种的构成调查结果相似,均认为台湾稻螟是目前广州稻区的优势种群(刘光华等,2006)。但此次调查结果发现台湾稻螟的种群优势度相对2006年有了进一步提升,虫口数比值从77.18%上升至97.01%,且我省优势钻蛀性螟虫三化螟2006年在广州地区仍有少量发生,而本次调查未见有三化螟的发生为害,这表明广州稻区钻蛀性螟虫种类、发生规律和为害规律出现了新变化。三化螟属单食性螟虫,混栽使得其寄主范围逐渐缩小,而频繁的、大面积的轮作使其食料出现断层,上述因素可能是导致广州稻区三化螟为害逐渐降低甚至消失的主要原因。
在空间分布上,由聚集强度指数和地统计学分析得出相同的结果,即广州地区以台湾稻螟为优势种群的稻螟幼虫的在分蘖期和抽穗期均呈聚集分布,这一空间分布同已报道的三化螟、二化螟和台湾稻螟3 种稻螟幼虫的空间分布相似,均呈聚集分布 (王华弟等,1994;陈观浩,1996;袁哲明等,2004)。调查田块分蘖期和抽穗期变异曲线最佳拟合模型均为高斯模型,空间异质性分蘖期由空间自相关性引起,而抽穗期由随机性因素引起,且分蘖期空间依赖距离大于抽穗期的空间依赖距离。从等值线图可见分蘖期和抽穗期稻螟幼虫均有明显的聚集团块,分蘖期主要集中在稻田边缘位置,而抽穗期则扩散至全田,说明稻螟幼虫的发生为害从稻田边缘向全田扩散。另由等值线图可知,分蘖期和抽穗期密度等值线均是南侧大于北侧,这可能是因为台湾稻螟是一种多食性螟虫,其寄主植物除水稻外,还包括玉米、甘蔗等多种作物(杨婉华和谢玲珊,2006),所以以台湾稻螟为优势种群的稻螟在靠近玉米地一侧(调查田块南侧)为害较严重与寄主植物玉米对稻螟成虫产卵的吸引作用有关,但具体作用机理是否如此有待进一步验证。对这一现象的合理阐释将有助于分析近年来我省部分水稻、玉米混栽区台湾稻螟为害逐年上升的新态势。
目前,化学防治仍是水稻螟虫防治的一项重要措施,取得良好防效的关键在于选择合适施药期。综合本研究对广州地区稻螟空间分布的研究及其世代历期(陈洪帆等,2010),早造,本地区第1 代螟虫的防治应选在分蘖期,并重点喷施稻田的边缘位置;第2 代螟虫的防治应选在孕穗期,同时进行全田均匀喷施。晚造,亦可参考早造在分蘖期和孕穗期预先采用平行取样法进行调查,对稻螟的虫口数及其空间分布进行预测,在此基础上决定是否进一步采取相应的防治措施。
References)
Chen GH.Study on the spatial patterns and sampling techniques of Chilotraea auricilia (Dudgeon) larvae [J].Hubei Plant Protection,1996,1:4-5.[陈观浩.台湾稻螟幼虫的空间分布型及抽样技术[J].湖北植保,1996,1996,1:4-5]
Chen Q,Zhang ZF,Wu WJ,et al.Geostatistic analysis of spatial pattern of Fruhstorferiola tonkinensis (Orthoptera:Catantopidae)Will nymphs [J].Chinese Journal of Applied Ecology,2007,18(2):467-470.[陈强,张振飞,吴伟坚,等.越北腹露蝗若虫空间格局的地学统计学分析[J].应用生态学报,2007,18(2):467-470]
Chen HF,Huang SS,Zhang YZ,et al.Control efficacy of Trichogramma japonicum against Chilo suppressalis and Chilaraea auricilia [J].Chinese Journal of Applied Ecology,2010,21 (3):743-748.[陈洪凡,黄寿山,张玉烛,等.稻螟赤眼蜂对二化螟和台湾稻螟的控制潜能评价[J].应用生态学报,2010,21 (3):743-748]
Ding YQ.Insects Mathematics Ecology [M].Beijing:Science Publishing Company,1994,43-53.[丁岩钦.昆虫数学生态学[M].北京:科学出版社,43-53]
Lecoustre L,Fargette D,Will PR,et al.1989.Analysis and maping of the spatial spread of African cassava mosaic virus using geostatistics and kriging technique [J].Phytopathology.79:913-920.
Liebhold,M.Zhang X,Hohn M E,et al.1991.Geostatisticalanal ysisof gypsymoth (Lepidoptera:Lymantriidae)eggmass populations[J].Environ.Entomol.,20:1407-1417.
Liu GH,Zeng L,Liang GW,et al.New trend of component occurrence and damage of rice stem borers in Guangdong Province [J].Journal of South China Agricultural University,2006,27 (1):41-43.[刘光华,曾玲,梁广文,等.广东稻区螟虫种类组成与发生为害新态势[J].华南农业大学学报,2006,27 (1):41-43]
Lu ZZ,Bao AM,Chen X,et al.Application of geostatistic software in pest management [J].Acta Ecological Sinica,2003,22 (6):132-136.[吕昭智,包安明,陈曦,等.地统计学软件在害虫管理中的应用[J].生态学报,2003,22 (6):132-136]
Li ZQ,Liang GW,Cen YJ.Geostatistic analysis of spatial pattern of the citrus red mite,Panonychus citri (McGregor) (Acarina:Tetranychidae)in citrus orchard [J].Journal of Environmental Entomology,2008,30 (3):202-206.[李志强,梁广文,岑伊静.柑橘全爪螨种群空间格局的地学统计学分析[J].环境昆虫学报,2008,30 (3):202-206]
Rossi RE,Mulla DJ,Journel AG,et al.Geostatistical tools for modeling and interpreting ecologyical spatial dependence [J].Ecological Monographs,1992,62:277-314.
Wang HD,Zhang ZS,Zhang ZC,et al.Field distribution of larva of yellow rice borer [J].Entomological Knowledge,1994,31 (3):132-135.[王华弟,张左生,张志昌,等.三化螟幼虫的田间分布[J].昆虫知识,1994,31 (3):132-135]
Wang ZQ.Geostatistics and its Application in Ecology [M].Beijing:Science Press,1999.[王政权.地统计学及其在生态学上的应用[M].北京:科学出版社,1999]
Xiang YY,Zhang F,Xiao BW,et al.Advances and control strategies of rice borers in China [J].Chian Plant Protection,2011,31(11):20-23.[向玉勇,张帆,夏必文,等.我国水稻螟虫的发生现状及防治对策[J].中国植保导刊,2011,31 (11):20-23]
Yuan ZM,Fu W,Li F.Spatial distribution pattern of Chilo suppressalis analyzed by classical method and geostatistics [J].Chinese Journal of Applied Ecology,2004,15 (4):610-614.[袁哲明,付威,李方.二化螟种群空间格局的经典分析与地统计学比较研究[J].应用生态学报,2004,15 (4):610-614]
Yang WH,Xie LS.Study on the characteristics of occurrence and control measures of Chilotraea auricilia (Dudgeon) [J].Anhui Agricultural Science Bulletin,2006,12 (2):77.[杨婉华,谢玲珊.台湾稻螟发生特点及防治对策[J].安徽农学通报,2006,12 (2):77]
Zhou Q,Lin DZ,Yao ZS,et al.Survey on the case of dominant population of rice borer in China at present [J].Chinese Bulletin of Entomology,1990,27 (6):321-324 [周圻,林德芝,姚忠生,等.我国目前稻螟优势种群区域化成因的调查研究[J].昆虫知识,1990,27 (6):321-324]
Zhou GF,Xu RM.Research on the Biogeostatistics [M].Beinjing:Science Press,1998.[周国法,徐汝梅.生物地理统计学[M],北京:科学出版社,1998]
Zhou Q,Zhang RJ,Gu DX.Spatial structure and pattern of Nilaparvata lugens population in large-scale[J].Chinese Journal of Applied Ecology,2001,12 (2):249-252.[周强,张润杰,古德祥.大尺度下褐飞虱种群空间结构初步分析[J].应用生态学报,2001,12 (2):249-252]
Zhang BY,Zhou SF,Zhang Y,et al.Research on population structure and major influencing factors of rice borer in Guangdong [J].Guangdong Agricultural Sciences,2007,6:51-53.[钟宝玉,邹寿发,张扬,等.广东省水稻螟虫种群结构及其主要影响因素[J].广东农业科学,2007,6:51-53]
猜你喜欢
河南农业·综合版(2023年8期)2023-08-12 18:25:15
现代农药(2023年1期)2023-04-15 11:35:30
中国农业科学(2022年8期)2022-05-16 05:15:10
新农村(浙江)(2021年8期)2021-09-09 16:04:32
四川蚕业(2021年3期)2021-02-12 02:38:50
四川蚕业(2021年1期)2021-02-12 02:03:18
四川农业科技(2018年12期)2018-03-17 16:17:25
吉林农业·下半月(2017年7期)2017-07-15 02:25:36
农民致富之友(2017年6期)2017-05-10 15:27:16
种业导刊(2015年3期)2015-01-22 06:19:15
