
21个闽楠种源叶片光合色素含量及叶绿素荧光参数分析
2014-11-06 05:09陈存及林达定林思祖
江西农业大学学报 2014年1期
刘 宝,陈存及,林达定,林思祖
(福建农林大学 林学院,福建 福州 350002)
植物的重要光合色素之一叶绿素不但有传递与吸收光量子的能力,同时其含量的多少也直接影响着植物的光能利用能力,同时它可以揭示在特殊的环境下植物的适应性、生物量,以及健康程度等[1-2]。植物光合生理的状况与光合机理通常可以用叶绿素荧光参数来表达,叶绿素荧光参数的测定目前是作为研究植物外界环境与其自身光合机理之间关系的重要手段[2-3]。光是植物生长不可缺少的重要生态因子之一,对植物的生理与生长发育都起到了重要的作用,随着植物的生长发育,植物的生理也发生着相应的变化[4]。
闽楠(Phoebe bournei)是樟科(Lauraceae)的常绿大乔木,也是我国重要的濒危珍贵用材树种之一。近年来,由于生态环境的不断恶化,再加上人们对其资源的掠夺式经营以及自身特性等原因,使得闽楠资源处于极度濒危状态,资源几尽枯竭[5-6]。目前国内对闽楠研究基础相对较薄弱,目前的研究主要体现在造林丰产技术、群落生态等方面,关于闽楠叶绿素荧光特性的研究还未见报道。本文收集了全国闽楠集中分布区的21个闽楠种源为材料,采用叶绿素荧光技术测定与分析了各种源的叶绿素荧光参数的基础上,综合比较与分析了闽楠不同种源间的光合与生理状况,为今后闽楠这一珍贵树种的良种选育工作提供一定的理论依据。
1 材料和方法
1.1 材料
研究材料来自于福建省松溪国有林场的6年生闽楠试验林的21个种源,具体种源情况详见表1。

表1 闽楠种源地理位置Tab.1 Geographic positions of provenances Phoebe bournei
1.2 研究方法
1.2.1 光合色素含量测定[7]取各闽楠种源植株1/2高度的相同部位成熟叶片快速洗净擦干。剪切成1~2 mm细条,称取0.5 g鲜叶样,做3次重复,放入具塞试管中,先加入20 mL无水乙醇,密封置于黑暗处放置1 d,转入50 mL刻度的容量瓶中,再加入15 mL无水乙醇,待试管中叶肉组织完全变白后,又转入容量瓶中,最后用无水乙醇进行定容并进行摇匀,最后取浸提液7 mL放置于10 mL的比色皿中,用无水乙醇做对照,在721型分光光度计上测定其在470,649,665 nm的光密度OD值。再根据叶绿素计算公式:
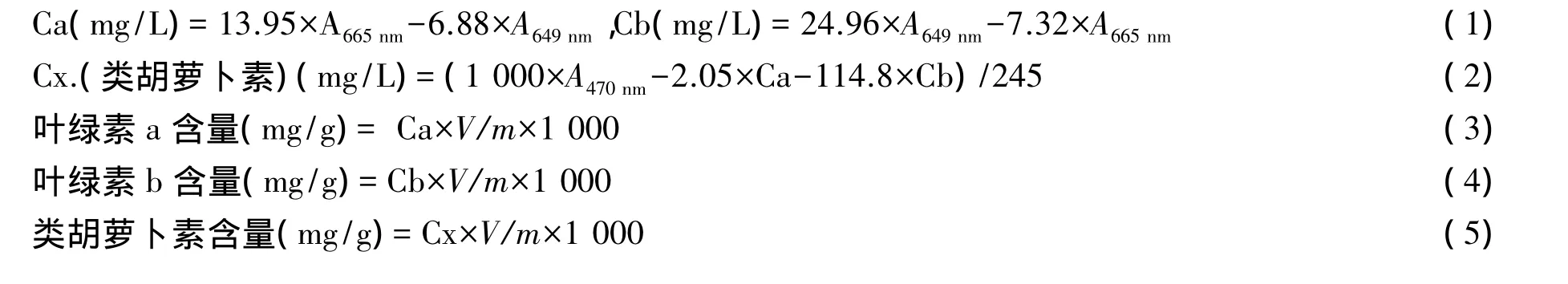
1.2.2 叶绿素荧光参数测定[8-11]在每个种源中选择并标记5片生长健壮、发育正常的叶子,然后把叶片放于暗处约20 min后,用型号为800MF/PSI便携式多谱动力学荧光成像系统对已标记的闽楠种源的叶片叶绿素荧光参数的测定。各荧光参数中:Fo代表初始荧光、Fm代表最大荧光、Fv=Fm-Fo代表可变荧光,Fv/Fm代表PSⅡ最大光化学效率或原初光能转换效率,Fv/Fo代表PSⅡ的潜在活性,ΦPSⅡ代表PSⅡ电子传递量子产率,Qp代表光化学猝灭系数,NPQ代表非光化学猝灭系数,Qy代表PSⅡ实际光化学效率。
1.3 数据分析
本文实验数据的方差分析与聚类分析均采用DPS 7.05版软件进行。
2 结果和分析
2.1 21个闽楠种源叶片光合色素含量的比较分析
在叶绿素中的少量叶绿素a具有把光能转化成化学能的功能,而其他大量的叶绿素a与叶绿素b都具备传递和吸收光能的功能[12]。从表2的数据分析表明:光合色素的含量在21个闽楠种源间存在显著差异,不同种源在吸收利用光能能力差异较大。各种源的叶绿素a含量大于叶绿素b的含量,说明在吸收利用光能的过程中,叶绿素a起着重要的作用。其中,叶绿素a含量最高的是8号种源为0.894 mg/g,是含量最低的 1 号种源为 0.271 mg/g的 3.08 倍,其次是 5 号种源(0.871 mg/g)说明 8 号种源和5号种源具有较强的吸收利用光能的能力。此外,由于类胡萝卜素具备光破坏防御与光能的捕获作用,因此类胡萝卜素能保护和协调光合作用,闽楠各种源间类胡萝卜素含量的差异也较显著,类胡萝卜素含量多介于0.1~0.2 mg/g,显示出闽楠各种源间的自我保护机制存在较大差异。
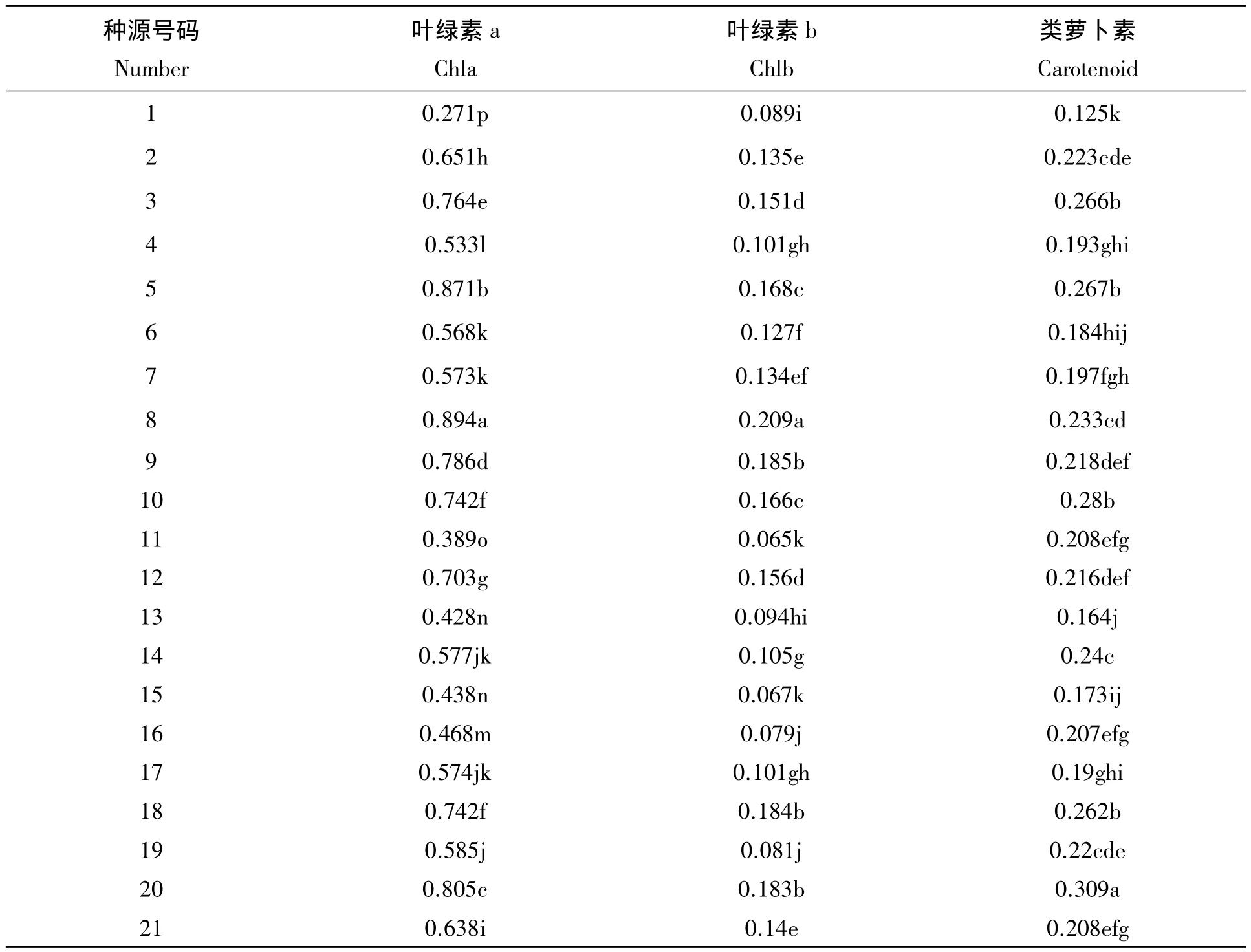
表2 闽楠不同种源叶片中光合色素含量的比较Tab.2 Comparison of photosynthetic pigment content of different provenance of Phoebe bournei
2.2 21个闽楠种源叶绿素荧光参数的差异及相关分析
2.2.1 21个闽楠种源叶绿素荧光参数分析 通过多重比较,从表3的分析结果可以得出:21个闽楠种源叶片的初始荧光(Fo)、最大荧光(Fm)、可变荧光(Fv)、PSⅡ最大光化学效率(Fv/Fm)、PSⅡ潜在活性(Fv/Fo)、非光化学淬灭系数(NPQ)、PSⅡ实际光化学效率(Qy)和PSⅡ电子传递量子产率(ΦPSⅡ)存在显著差异,光化学淬灭系数(Qp)差异不显著。

表3 21个闽楠种源叶片叶绿素荧光参数的比较Tab.3 Comparison of chlorophyll fluorescence parameter of 21 provenances of Phoebe bournei
Fv/Fm数值的波动通常可以用来评判植物受到光抑制的程度,其数值的变化表明了光系统PSⅡ光化学效率的波动[13-14];当植物的光照受到抑制时,Fv/Fm的值小于0.75,当植物在光照正常的条件下,Fv/Fm数值的变动区间在0.75~0.85,如果Fv/Fm的数值越低,表明该植物受到光照的抑制程度越大。从表3可以看出:21个闽楠种源的Fv/Fm值主要分布在0.75~0.80,表明21个闽楠种源的PSⅡ系统光能转换效率正常,其中5号、8号、20号种源的Fv/Fm数值大于其余的18个种源,表明这5号、8号、20号种源具有较高的光能转换效率。
Fv/Fo值随着具活性的 PSⅡ反应中心数量的增大而增大,其值能体现 PSⅡ潜在的光化学活性[13-14]。从表3可得到:Fv/Fo值的变化和Fv/Fm值的变化具有同步性。随着Fv/Fm值的增大,PSⅡ潜在的光化学活性也随之变大,具有活性的PSⅡ反应中心的数量相应地增多,因此植物生长需要的化学能就能从捕获的光能高效的转化而来。5号、8号、20号、14号种源的Fv/Fm值和Fv/Fo值均高于其他种源,说明这4个种源能较好地利用光能。
植物的热耗散能力常用NPQ值来表示[13-14]。从表3可得到:热耗散能力在21个闽楠种源间具有差异的显著,其中5号、8号和11号种源的NPQ 值较高(分别为0.96、0.89和0.88),表明5号、8号和11号种源的热耗散能力较强,它们的抗高温能力较强;NPQ值最低的是8号、7号种源,它们的NPQ值分别是0.53与0.59,表明7号、8号种源热耗散能力差。
原初电子受体的还原情况和PSⅡ开放的程度通常用Qp值表示[13-14]。从表3可得出,21个闽楠种源的Qp值主要的分布区间在0.70~0.75,各种源利用光能的效果较均一,种源间的Qp值差异不显著,其中8号种源和5号种源的Qp值最高,分别为0.80和0.78,表明8号、5号种源具有较强的把光能转化为化学能的能力。
Qy反映用于光合电子传递的能量占所吸收光能的比例,表示PSⅡ实际的光化学效率[13-14]。从表3可得出闽楠种源的Qy值主要分布区间在0.36~0.44,低于Fv/Fm的正常值较多,说明各种源把叶片所捕获的光能转化为植物所需的化学能的效率较低,其中8号、5号、20号种源具有较高的PSⅡ实际光化学效率。
ΦPSⅡ值的大小是反映植物光合能力的重要指标之一,它代表实际PSⅡ反应中心进行光化学反应的效率,同时也是PSⅡ反应中心部分关闭情况下的实际原初光能捕获的效率,如果植株的ΦPSⅡ值越大,其光能转化效率也就越高[13-14]。从表3可看出,种源间的ΦPSⅡ值差异显著,8号、5号、20号种源的ΦPSⅡ值较高,说明这3个种源具有较高的PSⅡ光能转化效率,更有利于碳同化的高效运转和有机物的积累。
2.2.2 21个闽楠种源叶绿素荧光参数的相关性分析 21个闽楠种源叶片的初始荧光(Fo)与最大荧光(Fm)、可变荧光(Fv)显著正相关,与非光化学淬灭系数(NPQ)呈极显著负相关,说明最大荧光产量和可变荧光产量及热耗散能力受到初始荧光产量值的影响较大;PSⅡ实际光化学效率(Qy)与可变荧光(Fv)呈极显著正相关,表明可变荧光值决定了21个闽楠种源的实际光合;PSⅡ电子传递量子产率(ΦPSⅡ)与最大荧光(Fm)、可变荧光(Fv)、PSⅡ潜在活性(Fv/Fo)、非光化学淬灭系数(NPQ)存在显著正相关,显示出闽楠不同种源叶片实际的PSⅡ反应中心进行光化学反应的效率将影响植物热耗散能力。因此,叶片实际的PSⅡ反应中心进行光化学反应的效率可以用于研究闽楠叶片实际光能利用情况。
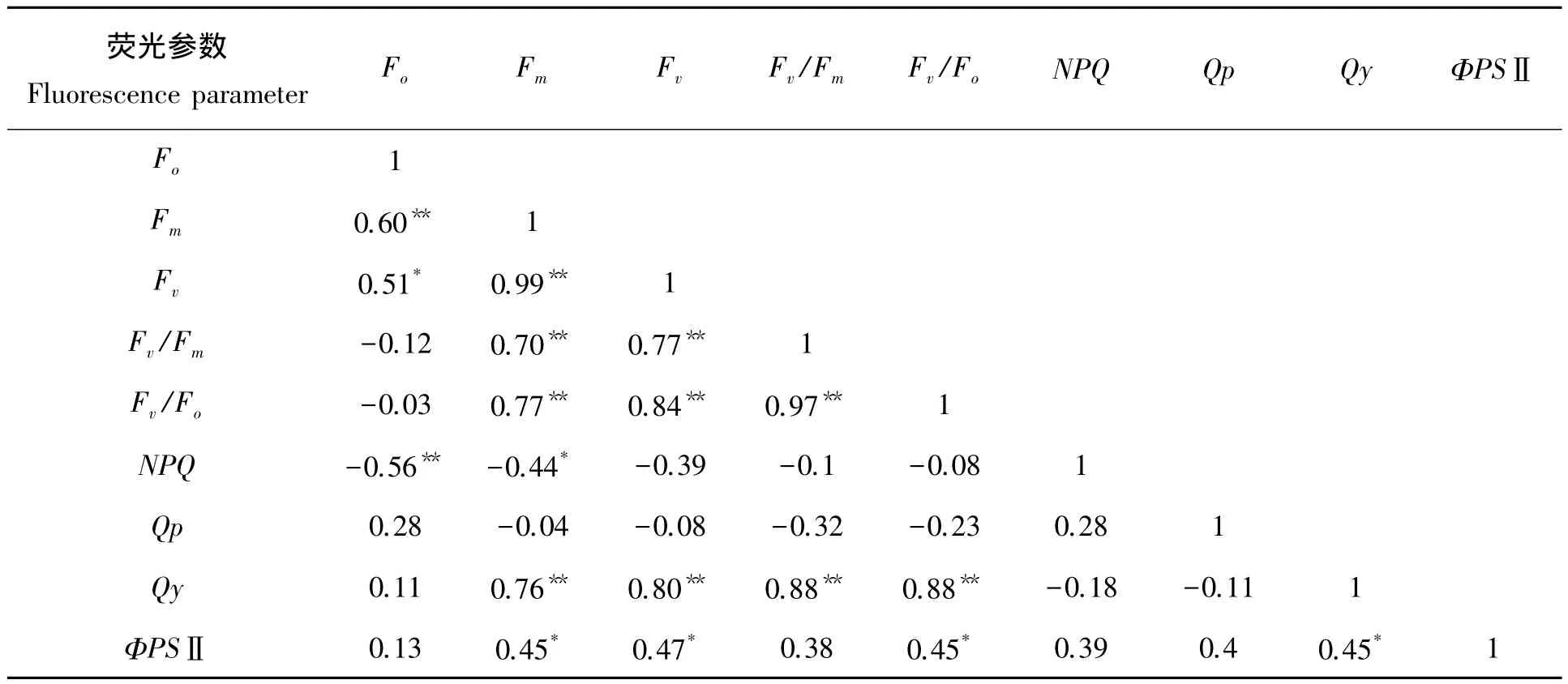
表4 21个闽楠种源叶绿素荧光参数的相关性分析Tab.4 Correlation analysis of chlorophyll fluorescence parameter of 21 provenances
此外,Fv/Fo与ΦPSⅡ的值呈显著正相关(表4),表明21个闽楠种源叶片的PSⅡ光能转换效率受到PSⅡ潜在的光化学活性的影响较大;Fv/Fo和Fm/Fo的相关系数为0.97,表明PSⅡ潜在的光化学活性受到PSⅡ的电子传递情况的显著影响。
2.3 21个闽楠种源的聚类分析
根据闽楠不同种源叶片的光合色素含量及叶绿素荧光参数测定结果,采用最短距离聚类法,获得了21个闽楠种源的聚类图(图1)。21个种源可明显划分为2类:第一类仅包含8号种源;第二类包含20个种源。第二类又可进一步划分为2个亚类:第一亚类仅包含种源5号;第二亚类共包含1号、2号、3号、4号、6号、7号、9号、10号、11号、12号、13号、14号、15号、16号、17号、18号、19号、20号和 21号等19个种源,其中10号和11号的距离最短,光合生理特征最接近;种源15号和16号、9号和21号、1号和2号、7号和17号也具有相近的光合生理特征。根据聚类分析结果结合叶绿素荧光参数的分析结果可以看出,5号、8号种源具有较好的光合生理性能。

图1 21个闽楠种源聚类分析Fig.1 Cluster analysis of 21 provenances of Phoebe bournei
3 讨论和结论
植物叶绿素吸收的光能除了一些以热量方式消耗或者用荧光重新发射出来外,其他大部分吸收的光能都用在植物的光合作用上[15-16]。21个闽楠种源叶片的叶绿素荧光参数和各光合色素的含量均不同程度地存在差异,揭示了21个闽楠种源对光能利用能力各不相同,其中叶绿素a在闽楠叶片利用光能时,起到了关键性的作用。8号、5号种源的叶绿素含量高、NPQ值较大,说明了这两个闽楠种源不仅具备良好的热耗散能力,而且还能较充分地利用光能,可为它们的生长发育提供充足的化学能,同时这两个种源还能用热量方式消耗多余的光能,因此它们的光合机构能够被更有效地保护。
21个闽楠种源叶片的Fv/Fm值的变动区间是0.75~0.80,说明各闽楠种源都具有较好的光能转换效果。从各闽楠种源的Qy值可得出,各闽楠种源之间在对实际光能利用率方面的差异不显著,同时,各种源把所得到的光能转化成化学能的效率较低。结合聚类分析的结果表明出5号、8号种源的光合生理性能较好,可作为较理想的闽楠种源选择。
ΦPSⅡ能够表征植物光能转化效率。闽楠各种源叶片的PSⅡ潜在的光化学活性与实际光合效率受到实际原初光能捕获效率的影响较大。因此,可以采用实际PSⅡ反应中心进行光化学反应的效率来研究闽楠各种源的实际光能利用率。本研究主要仅限于闽楠各种源叶绿素荧光特性方面的研究,今后我们将进一步研究闽楠种源的生长效果来进一步验证各种源荧光特性的结论,也为今后进一步探讨闽楠不同种源的生理、光合作用机制和优良种质资源的选育工作提供一定的参考价值。
[1]张友胜,张苏峻,李镇魁.植物叶绿素特征及其在森林生态学研究中的应用[J].安徽农业科学,2008,36(3):1014-1017.
[2]徐德聪,吕芳德,潘晓杰.叶绿素荧光分析技术在果树研究中的应用[J].经济林研究,2003,21(3):88-91.
[3]彭方仁,朱振贤,谭鹏鹏,等.NaCl胁迫对5个树种幼苗叶片叶绿素荧光参数的影响[J].植物资源与环境学报,2010,19(3):42-47.
[4]潘瑞炽.植物生理学[M].4 版.北京:高等教育出版社,2001:61-63.
[5]刘宝,陈存及,陈世品,等.闽楠群落优势种群结构与空间分布格局[J].福建林学院学报,2006,26(3):210-213.
[6]刘宝,陈存及,陈少杰,等.闽楠种源苗期试验初步研究[J].福建林学院学报,2007,27(3):213-216.
[7]李合生.植物生理生化实验原理和技术[M].北京:中国农业出版社,2001:45.
[8]孙猛,吕德国,刘威生.不同品种群15个杏品种荧光特性研究初探[J].中国农学通报,2011,27(6):171-176.
[9]陈四龙,孙宏勇,陈素英,等.不同冬小麦品种(系)叶绿素荧光差异分析[J].麦类作物学报,2005,25(3):57-62.
[10]吴飞燕,伊力塔,李修鹏,等.不同光照强度对石栎幼苗叶绿素含量及叶绿素荧光参数的影响[J].东北农业大学学报,2012,43(4):88-92.
[11]Maxwell I,Johnson G N.Chlorophyll fluorescence a practical guide[J].Journal of Experimental Botany,2000,51:659-668.
[12]吕福梅,沈向,王东生,等.紫叶矮樱叶片色素性质及其光合特性研究[J].中国农学通报,2005,21(2):225-228.
[13]张守仁.叶绿素荧光动力学参数的意义及讨论[J].植物学通报,1999,16(4):444-448.
[14]涂志华,卢仁锋,潘瑞,等.沿海沙地10个竹种叶绿素荧光特征比较[J].福建林学院学报,2012,32(2):131-135.
[15]何炎红,郭连生,田有亮.7种针阔叶树种不同光照强度下叶绿素荧光猝灭特征[J].林业科学,2006,42(2):27-31.
[16]张雪洁,谭晓风,袁军,等.低磷胁迫对油茶叶结汇素荧光参数的影响[J].经济林研究,2012,30(2):48-51.
猜你喜欢
特产研究(2022年6期)2023-01-17
云南化工(2021年9期)2021-12-21
辽宁林业科技(2021年1期)2021-03-16
中国新技术新产品(2019年21期)2019-01-20
电子技术与软件工程(2017年23期)2018-01-17
武夷学院学报(2017年9期)2017-11-29
科学家(2016年12期)2017-09-29
中国市场(2017年13期)2017-06-03
广西林业科学(2016年3期)2016-03-16
影像科学与光化学(2014年5期)2014-03-11
