
湘村黑猪繁殖性能的遗传参数估计
2014-10-18 07:39:06孙建帮杨仕柳李静如谢菊兰谭建坤彭英林
湖南畜牧兽医 2014年4期
朱 吉 ,刘 建 ,孙建帮 ,杨仕柳 ,李静如 ,谢菊兰 ,谭建坤 ,彭英林 ,※
(1.湖南省畜牧兽医研究所,湖南长沙 410131;2.湘村高科农业股份有限公司,湖南娄底417000;3.湖南省畜牧水产局,湖南长沙 410006)
随着育种技术的进步和发展,猪的育种目标不再单纯注重降低背膘厚、提高生长率,逐步发展为提高母猪的窝产仔数、产活仔数等繁殖性状,因此探索猪繁殖性状中各种效应的作用和规律,并将其运用到养猪育种实践中显得越来越重要。
育种方案的制定必须以准确、可靠的遗传参数估计为前提条件。国内外的大量研究证明,MTDFRE mL方法是目前动物育种中遗传参数估计较理想的方法,且得到了广泛应用。本研究旨在用MTDFRmL方法估计湘村黑猪某些繁殖性状的遗传力、母体效应和遗传相关,为选择提供必要的遗传参数[1~5]。
1 材料与方法
1.1 材料
本文所用资料取自2005~2010年湘村高科农业发展有限公司黑猪原种场进行的湘村黑猪选育群育种资料。本文分析的性状主要有:产仔数(TNB)、产活仔数(NBA)、初生窝重(LWB)和 21日龄窝重(LW21)等繁殖性状,总记录数为823胎次。
1.2 数据整理
用EXCEL对系谱资料和数据资料进行整理,然后利用SAS统计分析比较固定效应不同水平间的差异显著性,然后将差异不显著的水平合并从而减少水平数,根据结果重新合并固定效应、确定水平;随机效应中,母猪个体号计算个体的动物效应,系谱中的母猪号用于计算母体遗传效应、永久环境效应,母猪出生所在窝号计算窝效应。
1.3 混和模型:
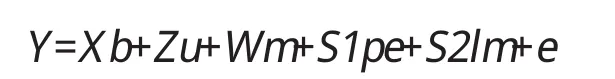
Y—观察值向量;b—固定效应向量(世代、季节、胎次);u—随机动物效应向量;
m—随机母体遗传效应向量;pe—影响同一母猪永久环境效应向量;t—母猪出生所在窝的窝效应向量;e—随机残差效应向量;X、Z、W、S1、S2-分别对应固定效应、随机效应、母体效应、永久环境效应、窝效应的结构矩阵。
2 结果与分析
2.1 表型参数
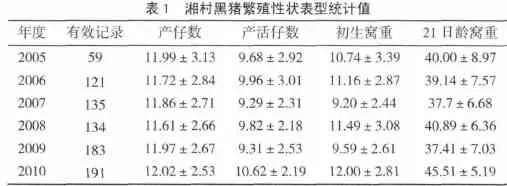
所分析的性状有效样本数、平均数和标准差见表1。由表1可见,经过五个世代的选育,湘村黑猪的各项繁殖性状基本上处于稳步上升的趋势,表明选育方法是适当的。2010年与2005年比较,产仔数、产活仔数、初生窝重、21日龄窝重分别提高了 0.03头、0.94头、1.26 kg、5.51 kg。
?
?
2.2 繁殖性状的遗传力
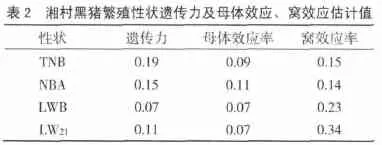
表2列出了湘村黑猪各繁殖性状的遗传力、母体效应率、窝效应率估计值。繁殖性状估计遗传力的变化范围为:0.07~0.19,TNB(头)、NBA(头)、LWB(kg)、LW21(kg)的遗传力估计值分别是 0.19、0.15、0.07、0.11。母体效应率、窝效应率的变化范围分别是 0.07~0.11、0.14~0.34。
2.3 繁殖性状间的相关分析
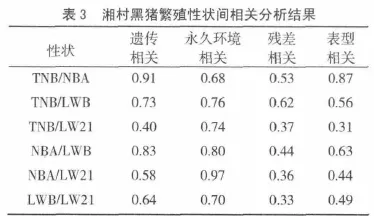
表3可见各繁殖性状间的相关分析结果。两两性状间遗传相关、永久环境相关、残差相关、表型相关范围分别为 0.40~0.91、0.68~0.97、0.33~0.62、0.31~0.87,遗传相关和表型相关均表现为TN B/NBA的最高(0.91,0.87),而 TN B/LW21最低(0.40,0.31),总的来说,性状间相关达到了中等偏上水平。
?
3 讨论
3.1 繁殖性状遗传力
繁殖性状的遗传力一般都很低。因为窝产仔数、窝重等窝性状由公猪、母猪及胚胎的基因型等因素互作决定。本试验结果,TNB、NBA、LWB、LW21的遗传力估值分别为0.19、0.15、0.07、0.11,略高于国外部分学者的报道结果[6-10]。分析差异产生的原因可能如下:①国内一般样本含量小,取样误差大,而国外样本含量大,取样误差小;②采用方法的不同;③估计遗传参数时,模型的选择和设计不同。
3.2 母体效应率和窝效应率
许多性状受母体环境影响,特别是对那些哺育早期度量的性状。当母性效应被忽视时,其它效应的方差组分就有可能被过高地估计。但目前对母性效应影响的估计结果相差较大,Preze[11]等结果发现母性效应不显著或没有,而Ferraz[12]等发现母性方差占表型方差的比例为0.01~0.13。本研究四个性状的母体效应率为0.08~0.10、窝效应率的变化范围分别为0.10~0.35,表明母体遗传效应、窝效应均占有一定比例,对繁殖性状产生一定的影响。
3.3 性状相关
两两性状间遗传相关、永久环境相关、残差相关、表型相关范围分别为0.40~0.91、0.68~0.97、0.33~0.62、0.31~0.87,基本处于中等偏上的正相关,符合一般规律。R.Roehe和B.W.Kennedy[13]报道了Yorkshire TNB与NBA的遗传相关为0.993,张伟峰[14]等报道北京花猪TNB和NBA的遗传相关为0.91,LBW与TNB、NBA的遗传相关为0.71、0.75,与本试验的结果基本相近。 □
[1] 张 沅,等.畜禽育种中的线型模型[M],北京:北京农业大学出版社,1993.
[2] 张 勤.家畜育种值和遗传参数估计方法的发展及现状[J],草食家畜,1990,(6):1-4,46.
[3] Monchai等,动物育种中解决混合模型复杂计算问题的软件应用[J],华南农业大学学报,2005,26(S1):83-78.
[4] 张胜利.遗传参数不同估计方法的比较[J],遗传,1995,17(6):21-23.
[5] 郑友民.用MTDFREML方法估计大白猪遗传参数[J],养猪,1999,(1):32-33.
[6] Cho.Y C,et al,Estimation of genetic parameters for major economic traits in swine[J],Korean journal of animal sicence,1998,40(5):447-454.
[7] Bidanel J P,et al,Use of a BLUP animal model for gnetic evaluation of French Large white and French Landrace Pig for Prolificacy[J],Pig news and information,1998,18(l):4.
[8] Estany J,et al.Estimation of genetic Parameters for liter size in Danish Landrace and yorkshire Pigs[J],Anim.sci,1995,60(2):315-324.
[9] Irgang R.,et al.,Genetic parameters for liter size of different paritied in Durco,Landrace and Large white sows[J].J of Anim.sci,1994,72(9):2237-2246.
[10] Rosa V C.Revista-Brasileria de genetica[J].1995,18(2):199-205.
[11] Misztel I,Perez-Enciso M.Sparse matrix inversion for REML estimation of variance components by exepctation maximization[J].J.diary Sci.1993,76(5):1479-1483.
[12] FerrnaJ B S,et al,Animal model estimation of genetic parameters and response to selcetion for litter size and weight,gorwth and backfatin c losed seedstock porductions of Large white and Landrace swine[J].J of Anim sci.1993,71(4):850-858.
[13] Roehe R,et al,estimation of genetic paramaters for little size in canadian Yorkshire and Landrace swine with each parity of forrowing treated different trait[J].J Anim Sci,1995,73:2959-2970.
[14] 张伟峰,等.应用动物模型REML估计猪的遗传参数的研究[J].养猪,1999,(3):31-32.
猜你喜欢
国外畜牧学·猪与禽(2024年4期)2024-01-01 00:00:00
养猪(2022年4期)2022-08-17 07:07:02
辽河(2021年10期)2021-11-12 04:53:58
读友·少年文学(清雅版)(2019年1期)2019-05-09 02:35:26
湖北畜牧兽医(2015年11期)2016-01-11 10:08:24
中国当代医药(2015年20期)2015-03-01 02:04:31
娃娃画报(2014年9期)2014-10-15 16:31:59
火炸药学报(2014年1期)2014-03-20 13:17:23
小朋友·快乐手工(2014年1期)2014-02-25 12:19:55
当代修辞学(2014年3期)2014-01-21 02:30:44
