
不同放牧梯度上草原群落植物功能群的结构组成及其变化
2014-09-19 06:42:26道日娜乌云娜霍光伟张庆昕贾子金
大连民族大学学报 2014年3期
道日娜,乌云娜,林 璐,3,霍光伟,张庆昕,贾子金
(1.内蒙古大学生命科学学院,内蒙古呼和浩特010021;2.大连民族学院
环境与资源学院,辽宁大连116605;3.辽宁师范大学生命科学学院,辽宁大连116029)
草原生态系统是中国陆地上最重要的生态系统类型之一,具有巨大的生态功能价值[1]。近年来,功能群概念的引入和功能群的划分,为研究生物多样性的生态系统功能和维持机制注入了新的活力[2-3]。许多研究者从不同的角度、尺度来对植物功能群进行研究,这些研究结果有不同的针对方向和目的,使人们可以从不同的角度更全面的理解复杂的陆地生态系统,这也大大拓宽了植物功能群的应用范围。
目前,功能群多样性、功能群丰富度及其类型已成为生态学和保护生物学新的热点问题[4]。对不同载畜率水平下内蒙古荒漠草原功能群物种组成及功能多样性变化特征的研究表明:灌木类和多年生禾草特性比较稳定,多年生杂类草随着载畜率的增加,物种组成下降幅度较大,使得群落结构和组成随载畜率的增加趋于简单化。丰富度指数和多样性指数在重牧区最低[5]。一年生、二年生和旱生植物功能群在不同退化阶段始终保持着较高的优势地位,对群落生态功能的发挥和维持起着重要作用。草地退化过程中不同群落的组成变化是物种适应性和群落环境变化相互作用的结果,而物种、功能群组成的变化基本反映了草地生态系统退化、群落结构简单化和环境基质稳定性减弱等一系列植被与生态环境变化的趋势[6]。李里认为,物种丰富度和物种多样性指数在中度退化草地最高,轻度退化草地次之,重度退化草地最低;群落生产力则在轻度退化草地最高,中度退化草地次之,重度退化草地最低,且与物种总数和多样性指数无显著相关性。相关分析结果表明,不同植物功能群的变化不尽相同,但功能群生物量与功能群内植物种数不存在明显的相关关系[7]。
功能群自提出以来,已经在功能群-干扰、功能群-全球变化及功能群-多样性研究中发挥了重要作用[8]。国内外相关研究更多的主要体现在自然或人为干扰背景下生态系统中单一功能群组成及动态变化,其研究结果尚难以说明植物功能群及其组成对生态系统过程的相对重要性,而且对功能群间的相互作用及其群落功能和稳定性的维持机制尚不太清楚,特别是对于自然生态系统运作机制的解释还一直缺乏令人信服的证据[9]。干扰与植被的关系主要表现在干扰对物种的进化意义和干扰对植被过程的调整作用,植物种对干扰的适应表现在繁殖、物候、形态、生理等方面。
本研究通过探讨放牧梯度上生活型和水分生态型植物功能群的变化,阐释草原退化的生态演替过程及其机制,为退化生态系统的恢复与重建提供科学依据。
1 研究区域概况
以蒙古高原东部呼伦贝尔草原的克鲁伦河流域草原为研究对象。研究区位于东经115°31'~117°43',北纬 47°36'~49°50',属中温带大陆性季风半干旱气候,年均降水量200~300 mm,年平均气温为-5~2℃,≥10℃的年积温2 320℃,持续日数125 d左右。地带性土壤类型主要为栗钙土。群落优势种为克氏针茅(Stipa krylovii)、羊草(Leymus chinensis)、糙隐子草(Cleistogenes squarrosa)等。
2 研究方法
2.1 样地设置
在研究区采用空间演替系列代替时间演替系列的方法,利用GPS定位,以具代表性的克氏针茅草原群落的建群种变化为依据选取轻度放牧、中度放牧、重度放牧三个实验样地(见表1)。参考王明玖和马长升[10]、郑阳[11]等对内蒙古典型草原载畜量的研究方法,结合实地调查结果,估算出三个样地的载畜量分别为0.62,1.55,2.79羊·hm-2。轻度放牧样地为围栏区,封育时间为2001年,季节性放牧,在生长季不受人为干扰;中度放牧样地为轮牧活动区,植物得以恢复生长;重度放牧样地为自由放牧区,常年受到放牧压力影响。

表1 研究样地基本特征
2012年7月,在各个样地选择典型地段设置一条50 m样线,间隔5 m做1 m×1 m的样方,记录10个样方内的植物种类组成、株丛数、种群的盖度(Penfound法)、平均高度和最高高度,采用刈割法获取种群的地上部现存量,80℃下烘干24 h,获得干物质量。
2.2 植物功能群的类型和划分
依据物种的生态生物学特性,将群落物种划分为生活型和水分生态型两种不同类别的功能群。生活型功能群包括:(1)一、二年生草本(Annual or biennial herb);(2)多年生杂类草(Perennial forbs);(3)多年生禾草(Perennial grasses);(4)灌木、半灌木类(Shrub,subshrub)。水分生态型功能群包括:(1)旱生植物(Xerophyte);(2)中旱生植物(Meso-xerophyte);(3)旱中生植物(Xero-mesophyte);(4)中生植物(Mesophyte)。
2.3 数据分析
(1)综合优势比
采用(SDR4)计算样方中各物种的综合数量指标:

式中,C'为相对盖度,H'为相对高度,D'为相对密度,W'为相对生物量(干重)。
(2)物种多样性
物种多样性分析采用Patrick丰富度指数(Pa)和香农-韦纳(Shannon-Weaver)指数(H'):
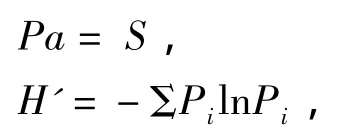
式中,S为物种数,Pi为各种植物的相对生物量,i代表第i个物种(其中i物种在整个样带内的平均优势度为i物种在该样带所有样方内的优势度之算术平均值)。
(3)物种均匀度指数
均匀度采用Pielou均匀度指数(E):

式中,H'为Shannon-Wiener指数,S为物种数。
3 结果与讨论
3.1 不同放牧梯度上群落物种组成的变化
不同放牧梯度上的种类组成与出现频率见表2。
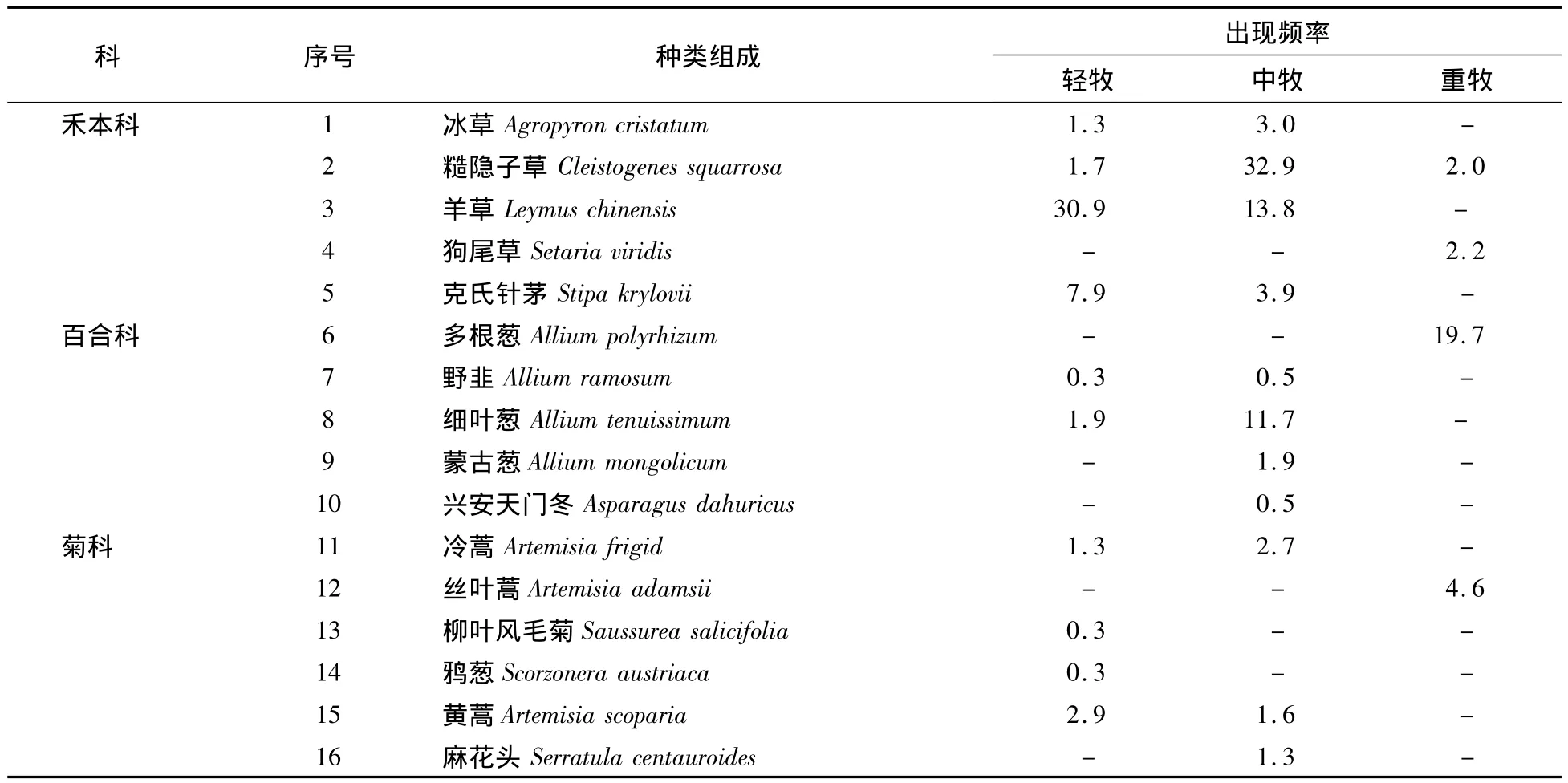
表2 不同放牧梯度上的种类组成与出现频率

续表
表2表明,在克鲁伦河流域物种组成有32种,且不同放牧压力对物种组成结构影响较大,随着放牧强度的增加,物种组成呈递减趋势,轻牧、中牧、重牧下物种数分别为23,18,10种。轻牧状态下出现频度较高的植物有羊草和阿氏旋花,其次为克氏针茅、星状刺果黎、尖头叶藜,出现频度较少的是乳白花黄芪、披针叶黄华、猪毛菜、燥原荠;中牧出现频度较高的植物有糙隐子草、尖头叶藜,其次是蒙古葱、黄囊苔草,出现频度较少的是狭叶锦鸡儿;重牧出现频度较高的植物有黄囊苔草、多根葱。不同物种在不同放牧梯度上的变化规律亦不同,呈递减趋势的物种有克氏针茅、羊草、黄蒿、星状刺果藜,呈增加后降低的物种有冰草、糙隐子草、细叶葱、蒙古葱、冷蒿、猪毛菜、尖头叶藜等,呈增加特征的物种则有狗尾草、多根葱、丝叶蒿、黄囊苔草。
同一科物种在不同放牧梯度下出现的频度差异较大,禾本科在轻度、中度、重度放牧下呈现频度最高,依次为克氏针茅、羊草,其次为糙隐子草、冰草,频度最低的是狗尾草;藜科植物的变化特征为星状刺果藜频度最高,其次是猪毛菜、星状刺果藜,频度最低的是猪毛菜;百合科植物均在中度放牧下出现频度较高,其中频度最高的为细叶葱;菊科物种则以丝叶蒿在重度放牧下出现频度最高;随着放牧强度的增加,莎草科的黄囊苔草递增明显,旋花科的阿氏旋花递减明显;苋科的尖头叶藜则表现明显的先增加后降低的变化趋势。
3.2 不同放牧梯度上植物功能群的结构组成
不同放牧梯度上各个功能群的物种数及其百分比见表3。
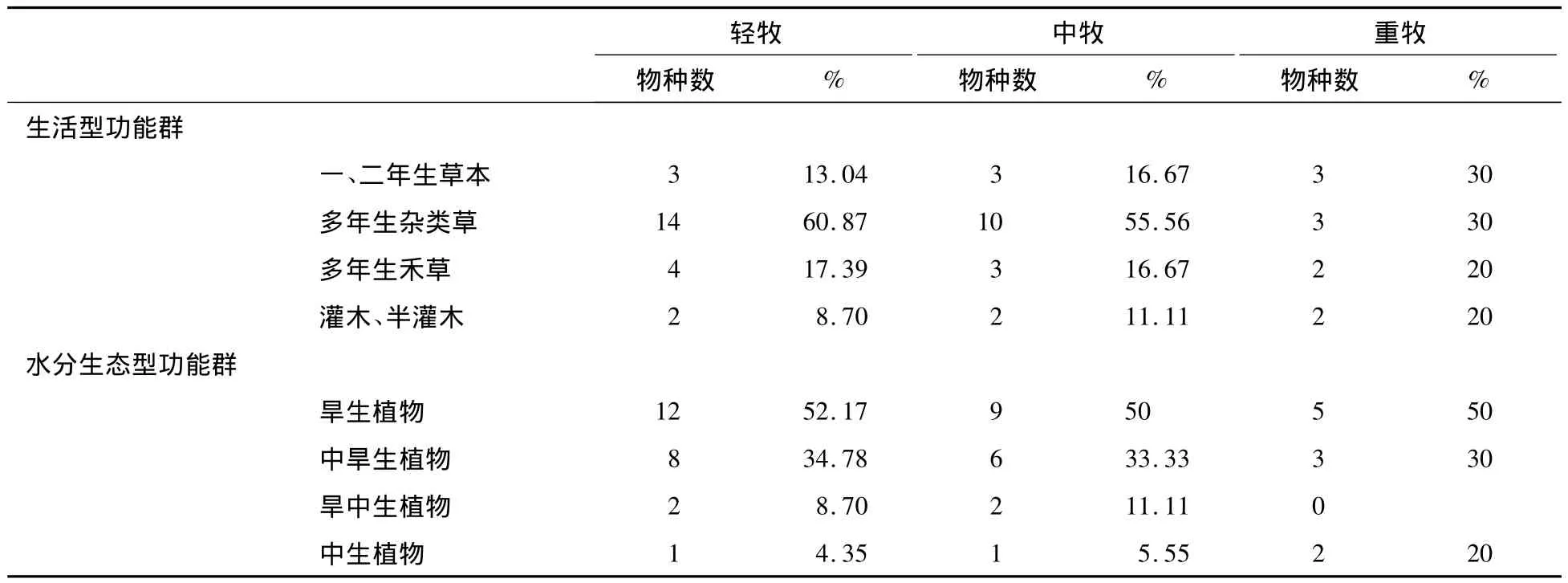
表3 不同放牧梯度上各个功能群的物种数及其百分比
从各生活型功能群的种类组成看,多年生禾草在轻牧、中牧和重牧中物种数分别为4,3,2,其中糙隐子草为共有种;多年生杂类草共20种,轻牧阶段有14种,中牧下10种,重牧中3种,其中黄囊苔草为共有种;一、二年生草本共4种,其中共有种为尖头叶藜和猪毛菜。种数最多的是多年生杂类草,在各放牧梯度上均表现出最大值,其次是多年生禾草,且多年生杂类草和多年生禾草在重牧区迅速降低;一、二年生草本植物在各放牧梯度上分布均匀;而灌木、半灌木功能群种数最少且基本保持不变。
从水分生态功能群的物种结构看,中生植物共2种,中旱生植物共11种,旱中生植物共2种,旱生植物共17种。物种数最多的是旱生植物,其次是中旱生植物,第三是旱中生植物,最小的是中生植物。旱生、中旱生、旱中生植物的种数由轻牧到重牧均逐渐降低,而中生植物随放牧梯度的增加呈上升趋势。
3.3 不同放牧梯度上植物功能群综合优势比的变化
3.3.1 不同放牧梯度上生活型功能群综合优势比的变化
在不同放牧梯度上,一、二年生草本、多年生杂类草、多年生禾草及灌木、半灌木各功能群的综合优势比存在较大差异(如图1)。
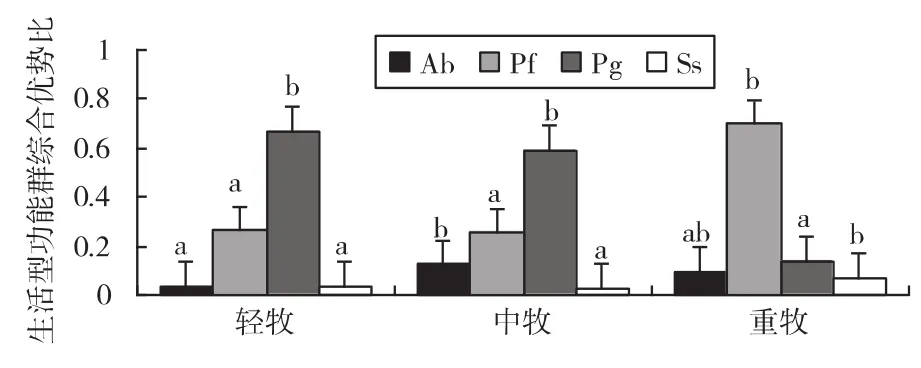
图1 生活型功能群综合优势比的变化
由图1可知,从轻牧到重牧,灌木、半灌木的综合优势比逐渐升高,依次为0.033 3,0.027 6及0.067 5,灌木、半灌木在轻牧和中牧间差异不显著(p>0.05),但两者与重牧差异显著(p<0.05);多年生禾草的综合优势比呈相反趋势,其优势比依次为:轻牧0.6675、中牧0.593 5、重牧0.139 8,重牧与轻牧、中牧差异性显著;多年生杂类草的优势比在重牧下达到最高,数值为0.697 8,与中牧(0.2531)、轻牧(0.2632)差异显著;一、二年生草本的优势比在中牧阶段最高,为0.125 8,与轻牧区(0.0383)差异显著,与重牧区(0.0948)差异不显著。
3.3.2 不同放牧梯度上水分生态型功能群综合优势比的变化(如图2)
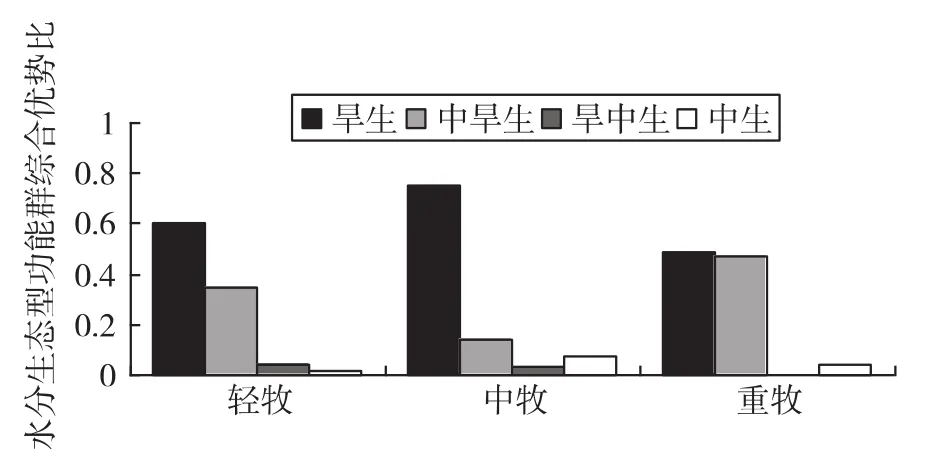
图2 水分生态型功能群综合优势比的变化
由图2可知,旱生植物在中牧阶段的优势比最高,为0.754 4,轻牧为0.600 4,重牧为0.485 7;而中旱生植物在中牧阶段的优势比最低,为0.143 4,在重度最高,为0.468 9;旱中生植物随着放牧梯度的增强,优势比呈现逐渐降低的趋势,在轻牧阶段最高,为0.037 8;中生植物在不同放牧梯度上优势比的变化与旱生植物相同,在中牧阶段最高,为0.072 8。
3.4 不同放牧梯度上植物功能群组成的变化
3.4.1 不同放牧梯度上生活型功能群组成的变化
生活型的差别反映了群落的生境,特别是小生境、小气候的变化[12]。不同放牧梯度上组成群落的各生活型种数百分比的变化如图3。

图3 群落生活型功能群组成的变化
由图3可知,不同放牧强度下,植物生活型功能群组成基本一致,均含有四个功能群。灌木、半灌木所占百分比在重牧区中最高,为17.8%,在中牧区为9.1%,轻牧下为7.3%。中牧与轻牧、中牧与重牧差异性不显著(p>0.05),轻牧、重牧差异性显著(p<0.05);多年生禾草所占百分比在轻牧、中牧、重牧区分别为22.4%,20.2%,21.5%,三者差异性均不显著。多年生杂类草在轻牧区所占比例最高,为59.1%,其次是中牧区,为52.9%,最次是重牧区,为32.2%。轻牧、中牧差异性不显著,重牧与中牧、重牧与轻牧差异性显著。一、二年生草本所占百分比呈递增趋势,在轻牧、中牧区分别为11.2%,17.8%,轻牧、中牧差异性不显著,在重牧区占28.5%,中牧、重牧差异性显著。
3.4.2 不同放牧梯度上水分生态型功能群组成的变化
植物的水分生态类型是依据植物对生态环境中水分状况的适应性而划分的。不同放牧梯度直接影响着草原群落的盖度,进而影响水分生态环境。群落水分生态型功能群组成的变化如图4。
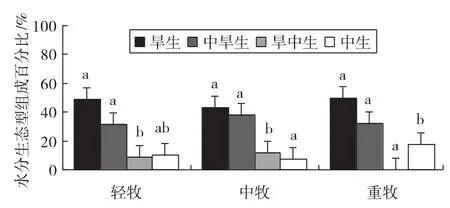
图4 群落水分生态型功能群组成的变化
由图4可知,随着放牧梯度的逐渐增强,旱生植物功能群种数的百分比基本保持不变,中旱生植物在中度放牧中所占比例最高,而旱中生植物种数百分比随放牧强度的增加呈现先增强后消失的特征,中生植物种数的百分比随放牧强度的增强呈现先增加后减少的趋势。旱生植物在轻牧、中牧及重牧区所占比例分别为49.10%,43.33%及50.00%,各放牧梯度间差异不显著(p>0.05)。中旱生植物在轻牧、中牧及重牧区分别为31.60%,37.78%及32.22%,并且各放牧梯度间差异不显著。旱中生植物在中牧区占11.66%,与重度区(占0.00%)差异显著(p<0.05),而与轻牧区(占8.85%)差异不显著。中生植物在轻牧下占10.44%,它与中牧、重牧所占百分比均差异不显著,中牧(占7.22%)与重牧(占17.78%)差异显著。
3.5 不同放牧梯度上植物功能群生物量的变化
3.5.1 不同放牧梯度上生活型功能群生物量的变化(如图5)

图5 生活型功能群生物量的变化
放牧梯度对于草地生产力具有决定性作用。由图5可知,在生活型功能群组成中,多年生禾草占有绝对优势,在轻牧区的生物量最大,为53.45%,并随放牧梯度的加大而减少;多年生杂草在轻牧区所占比例最大,为40.78%;灌木、半灌木在各放牧区的生物量相近;一、二年生草本植物在轻牧区较少,中牧区和重牧区占明显优势。
3.5.2 不同放牧梯度上水分生态型功能群生物量的变化(如图6)
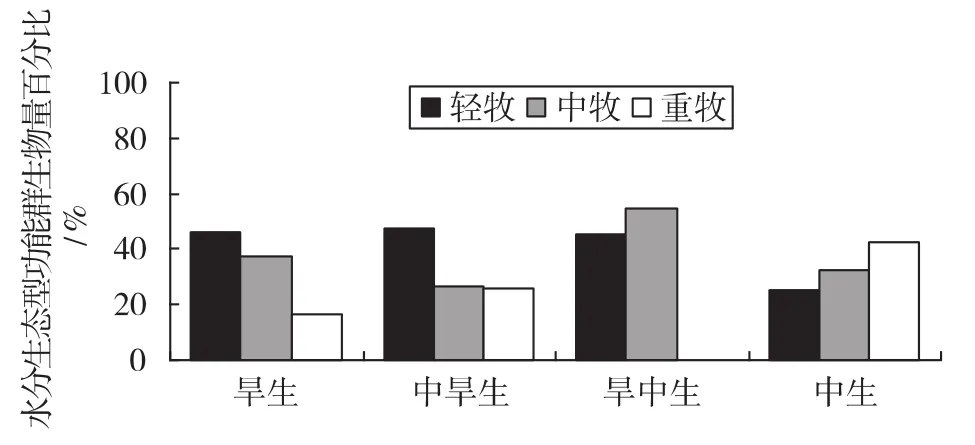
图6 水分生态型功能群生物量的变化
由图6可知,旱生植物在轻牧区和中牧区占绝对优势,在轻牧区生物量最大,为46.35%;随放牧强度的加大,相对生物量呈现逐渐减少的趋势,在中牧区和重牧区生物量依次为37.20%和16.45%;中旱生植物相对生物量在轻牧区最大,为47.46%,在中牧区和重牧区所占比例接近,分别为26.49%和26.03%;旱中生植物在重牧区未被统计,可能与选取样地的局限性有关;旱中生植物和中生植物在各个放牧区的相对生物量均较少,所占比例分别为3.35%和3.80%。
3.6 不同放牧梯度上功能群和群落多样性的变化
3.6.1 不同放牧梯度上功能群多样性的变化
从轻牧到重牧,不同生活型功能群和水分生态型功能群的多样性变化见表4。可以看出,生活型功能群中,多年生杂类草的丰富度指数随放牧梯度加强而明显减小,在轻牧区最高,为14;多年生禾草的均匀度指数在各放牧梯度上均较高,在中牧区最高,为0.77;多年生杂类草的多样性指数随放牧梯度的增加呈现减小趋势,重牧区多样性指数与轻牧区和中牧区有显著性差异;多年生禾草的多样性指数在重牧区最低,Shannon-Weaver指数为0.417 6,中牧区多样性指数与轻牧区和中牧区有显著性差异;而一、二年生草本与灌木、半灌木相同,随放牧强度的加大而呈现递增规律,并且轻牧区多样性指数与重牧区有显著差异。
水分生态型功能群中,旱生植物和中旱生植物的丰富度指数随放牧梯度的增加呈现递减趋势,且在重牧区均匀度指数较高;中旱生植物多样性指数在轻牧区最高,重牧区多样性指数与轻牧区和中牧区有显著性差异;旱生植物的多样性指数在中牧区最高,轻牧区多样性指数与重牧区有显著性差异;中生植物多样性指数随放牧梯度的增强而增加。

表4 不同放牧梯度上植物功能群多样性的变化
3.6.2 不同放牧梯度上群落多样性的变化
群落的物种丰富度及多样性是群落的重要特征。不同放牧梯度上群落多样性及其相关关系见表5。可以看出,中牧下多样性指数最高,为2.25,其次为轻牧区,为2.20,但二者不存在差异(p>0.05),重牧下多样性指数最低,为1.95。重牧区多样性与中牧、轻牧区有显著性差异(p<0.05)。多样性指数在中牧下最大,随着放牧强度的加大,呈单峰式曲线变化特征,符合“中度干扰”假说,这与不同植物功能群在放牧退化演替过程中的消长变化密切相关。由于牧草在营养价值和采食难易程度等方面的差异,家畜具有择食性,可显著改变植物种间竞争格局、物种侵入或迁出以及群落组成。

表5 不同放牧梯度上群落多样性及其相关关系
从轻牧到重牧,丰富度指数呈递减趋势,轻牧区丰富度指数最高,为18.00,三个放牧阶段间均呈现显著差异;均匀度指数呈递增趋势,在重牧区最高,为0.87,重牧与中牧条件下二者不存在显著性差异,而轻度区为0.76,与中牧、重牧有显著性差异。
4 结论
(1)不同功能群对草原退化的响应程度不同。植物生活型功能群中,多年生杂类草在轻牧、中牧区都占有明显优势,多年生杂类草的生物量随着放牧强度的增加显著减少,说明在群落植物功能群组成中,多年生杂类草的作用逐渐减弱,一、二年生草本在重牧区占有绝对优势,使群落结构趋于简单,生态系统稳定性变低;植物水分生态型功能群中,旱生植物在轻牧区、中牧区和重牧区均占有绝对的优势。
(2)从植物功能群的地上生物量来看,生活型功能群组成中,多年生禾草占有绝对优势,在轻牧区的分布量最大;多年生杂类草在轻牧区占有优势,一、二年生草本植物在中牧区和重牧区较有优势。水分生态型功能群中,旱生植物在轻牧区和中牧区占绝对优势,随放牧强度的加大,相对生物量呈现逐渐减少的趋势。
(3)从物种多样性的变化来看,多年生杂类草和多年生草本功能群多样性指数随放牧梯度的增加呈现减小趋势,一、二年生草本与灌木、半灌木以及中生植物功能群多样性,随放牧强度的加大而呈现递增规律。由轻牧到重牧,物种丰富度指数逐渐降低,而均匀度指数逐渐增加,Shannon-Weaver指数在中牧区最大,符合“中度干扰”假说。适度放牧是保护草原生物多样性、维护放牧生态系统功能与健康的有效途径。
[1]尹剑慧,卢欣石.中国草原生态功能评价指标体系[J].生态学报,2009,29(5):2622-2630.
[2]KOBRNER C H.Scaling from species to vegetation:the usefulness of functional groups[M]∥SCHULZE E D,MOONEY H A.eds.Ecosystem function of biodiversity.Berlin:Springer,1994,99:117-140.
[3] WILSON J B.Guilds,functional types and ecological groups[J].Oikos,1999,86:507-522.
[4]林丽.石羊河上游退化草地植物功能群特征研究[D].兰州:甘肃农业大学,2007.
[5]焦树英,韩国栋,赵萌莉,等.荒漠草原地区不同载畜率对功能群特征及其多样性的影响[J].干旱区资源与环境,2006,20,161-165.
[6]马建军,姚红.内蒙古典型草原区3种不同草地利用模式下植物功能群及其多样性的变化[J].植物生态学报,2012,36(1):1-9.
[7]李里,刘伟.退化草地植物功能群和物种丰富度与群落生产力关系的研究[J].草地学报,2011,19(6):917-921.
[8]李荣平,刘志民,蒋德明,等.植物功能型及其研究方法[J].生态学杂志,2004,23(1):102-106.
[9]MC CANN K S.The diversity stability debate[J].Nature,2000,405:228-233.
[10]王明玖,马长升.两种方法估算草地载畜量的研究[J].中国草地,1994(5):19-22.
[11]郑阳.内蒙古克氏针茅典型草原生态系统放牧管理优化模式研究[D].兰州:甘肃农业大学,2010.
[12]王正文,邢福,祝延成,等.松嫩平原羊草草地植物功能群组成及多样性特征对水淹干扰的响应[J].植物生态学报,2002,26(6):708-716.
猜你喜欢
数学物理学报(2021年6期)2021-12-21 06:24:38
西藏艺术研究(2021年3期)2021-06-02 09:37:16
应用数学(2020年2期)2020-06-24 06:02:50
西藏农业科技(2019年1期)2019-07-25 00:37:08
数学年刊A辑(中文版)(2018年2期)2019-01-08 01:59:52
成才之路(2017年21期)2017-07-27 20:52:40
安徽农学通报(2017年9期)2017-05-19 23:03:05
现代农业科技(2016年22期)2017-03-24 12:21:57
兽医导刊(2016年6期)2016-05-17 03:50:31
学周刊·下旬刊(2015年10期)2015-07-22 12:36:17
