
抚育间伐对油松人工林光合作用的影响1)
2014-09-18 11:11韩海荣康峰峰程小琴刘可周彬
东北林业大学学报 2014年2期
李 勇 韩海荣 康峰峰 程小琴 刘可 周彬
(北京林业大学,北京,100083)
密度是决定林分结构的重要因子之一,密度控制是提高人工林生产力的关键措施。以往人们对人工林生产力的研究,常常着重在产量角度,认为林分的高密度是提高单位面积高蓄积量和生长量的前提条件。但研究表明,在高密度条件下,产量不一定高,经济效益也不一定好[1-2]。抚育间伐是调节林分密度的重要手段,适当的密度控制,不仅能提高林分的稳定性,也能取得可观的经济收入。自20世纪70年代以后,不少学者对抚育间伐方法、间伐强度、生长效应等做了大量研究,并用来指导林业生产[3-7]。研究表明,林木生长到一定年龄需要间伐来改善林木生长条件,抚育间伐在森林经营管理中起着不可或缺的作用。抚育间伐通过对单位面积林木株数进行合理调整,以便改善林内的水热条件、林分生产力和土壤状况,进而使林分发挥最大的生态效益[8-12]。
油松(Pinus tabulaeformis)是我国特有的用材树种,适应性强,可生长在砂地、微酸性土、中性土及钙质黄土上,根系发达,也是黄土高原大面积栽植的水土保持树种[13-15]。前人对油松光合生理特性、环境因子对光合作用的影响等进行了研究[15-19],但对油松光合作用对人工抚育干扰的响应未见报道。本文通过野外活体测定不同抚育强度下油松林上层当年生针叶的气体交换、叶绿素质量分数等指标,分析人工抚育干扰对油松光合作用的影响,以及干扰后影响油松光合作用的关键因子。以期探讨怎样的抚育强度更利于油松人工林的生长,为油松人工林实现生态效益和经济效益最大化目标提供数据基础。
1 研究区概况
本研究地位于“油松之乡”山西省太岳山国有林管理局马泉林场宋家沟,其位于太岳林区中部,沁源县东部(东经 112°15'~112°33'、北纬 36°31'~36°44')。该区气候属暖温带半湿润大陆性季风气候,年均气温8.6℃,年均日照时间2600 h,年均无霜期179 d。四季分明,春季多风,雨季集中在7—9月,年均降水量662 mm,相对湿度60%~65%。地貌属大起伏喀斯特侵蚀高中山,地形西部陡峭,东部平缓。基岩以花岗岩和石灰岩为主。地带性土壤为黑炉土,山地森林有褐土和棕色森林土发育,高山顶部有草甸、草原土。油松(Pinus tabulaeformis)、辽东栎(Quercus liaotungensis)、白桦(Betula platyphylla)、山杨(Populus davidiana)是本区最有代表性树种,也是主要森林植被,其中油松是优势树种。
2 研究方法
以山西太岳山研究地20年生油松人工林为研究对象,在阳坡选择林相整齐、密度一致的油松林,随即设置4个样地(20 m×30 m)进行样地调查。将4个样地按照密度进行人工干扰,设置对照、低等强度(间伐20%)、中等强度(间伐30%)、高等强度(间伐40%)4种强度梯度。确定标准木,每个样地选择3株标准木。在标准木上层、朝阳侧枝上选择代表性的当年生枝,以活体针叶小枝为测定单位。
光合日变化测定:在油松生长季盛期选择当地典型的天气,从 7:00—17:00,用 Li-6400便携式光合仪测定光合参数,每隔2 h测定1次。每株选取3组针叶,每组记录3组数据,取平均值。测定的参数有净光合速率(Pn)、气孔导度(Cond)、胞间CO2摩尔分数(Ci)、蒸腾速率(Tr)等。
光响应曲线:在油松生长季盛期选择晴朗无云的天气,在9:00—11:00时,采用 Li-6400便携式CO2/H2O红外气体分析仪活体测定油松针叶的气体交换过程。光合有效辐射(RPA)设定为1800、1500、1200、1000、800、600、400、200、150、100、80、50、20、0 μmol·m-2·s-1共 14 个梯度值,叶室温度控制为 30℃,CO2摩尔分数控制为 400 μmol·mol-1。
叶绿素的测定:精确称取0.300 g油松针叶鲜叶片,加入V(80%丙酮)∶V(无水乙醇)=1∶1提取液10 mL,30℃黑暗浸提光合色素,直到叶片全部变白。UV-1700型分光光度计分别在440、645、663 nm处测定其吸光度值A440、A645和A663,重复3次[20-21]。
叶面积的测定:叶面积实际测量方法参考李轩然等[22]的游标卡尺测量法。
根据测量参数求出油松针叶实际叶面积,将实际叶面积输入Li-6400相应的处理软件,重新换算数据。叶片的表观量子效率利用师生波等[23]直线回归法,其他光合参数利用张国平等[24]模型拟合。数据的统计分析用SPSS18.0来完成,图形绘制用Excel软件完成。
3 结果与分析
3.1 油松人工林光合日进程对人工抚育干扰的响应
图1表明,不同强度人工干扰下的油松人工林中,大气CO2摩尔分数、大气相对湿度(HR)、光合有效辐射(RPA)、大气温度的日变化均为“单峰”曲线。大气CO2摩尔分数和大气相对湿度在中午较低,早晚较高。1 d中对照组油松人工林大气相对湿度均值大于不同强度人工干扰后油松林的大气相对湿度,而大气CO2摩尔分数1 d中变化幅度较小,最小值出现在13:00左右。大气温度和光合有效辐射早晚较低,中午较高,光合有效辐射12:00左右达到1 d最大值,大气温度在13:00左右达到最大值。随着人工干扰强度的增大,大气温度和光合有效辐射逐渐增加,而空气相对湿度逐渐减小。
另外,不同强度人工干扰后的油松人工林净光合速率日均值均大于对照组,且中等强度的人工抚育下的净光合速率日平均值最大,即净光合速率日均值中等强度人工干扰(6.42 μmol·m-2·s-1)>高强度人工干扰(5.01 μmol·m-2·s-1)>低强度人工干扰(4.84 μmol·m-2·s-1)>对照(4.33 μmol·m-2·s-1)。这说明了不同强度的人工抚育干扰对油松光合作用起到促进作用,适度的采伐强度可明显起到促进作用。
表1为不同强度人工干扰下油松净光合速率与主要环境因子之间的关系特征。不同强度干扰下油松净光合速率与光合有效辐射、大气温度、相对湿度、CO2摩尔分数呈二次多项式关系(Pn=bX2+aX+c)。从整体上看,光合有效辐射是限制油松净光合速率的关键因子。
对光合作用与主要环境因子进行双变量线性拟合Z=aX+bY+c(表2),以期进一步阐明不同强度人工干扰之后影响油松光合作用的关键因子。从整体上看,有光合有效辐射的双因子组,与净光合速率之间相关性都很显著。这表明光合有效辐射是人工抚育后影响油松光合作用的关键因子,这与单因子分析下的结果相一致。
3.2 油松人工林光响应对人工干扰的响应
不同强度人工干扰下的油松光响应曲线如图2所示,3种强度人工干扰后的油松人工林的Pn值大于对照组油松人工林的Pn值,其中,中等强度干扰和低等强度干扰下的Pn值明显大于高强度人工干扰下的Pn值。当光合有效辐射(RPA)≤800 μmol·m-2·s-1时,低强度干扰下的 Pn值接近中强度干扰下的 Pn值;当 RPA>800 μmol·m-2·s-1时中强度干扰下的Pn值大于低强度干扰下的Pn值。其中,最大净光合速率中等强度干扰下最大((12.01±0.99)μmol·m-2·s-1),这说明中等强度干扰下的油松具有更强的光合潜力。

图1 环境因子的日变化单峰曲线
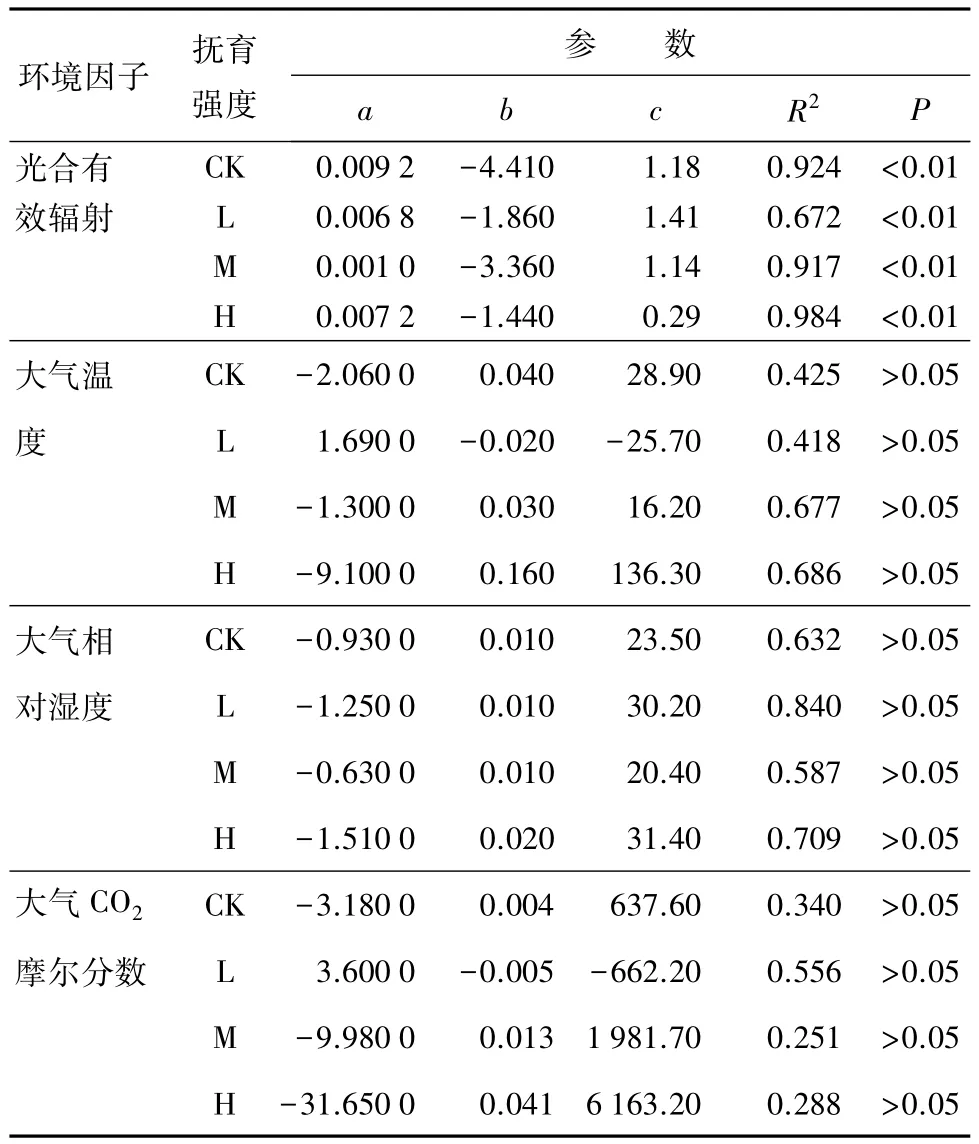
表1 不同强度人工抚育干扰下净光合速率与主要环境因子的二次多项式关系

表2 不同强度人工抚育干扰下净光合速率与环境双因子间的线性关系

图2 不同光照时油松不同强度人工干扰下针叶的净光合速率
3.3 油松人工林光合特性参数对人工抚育干扰的响应
光合特性参数是光响应特性的重要指示指标。表3反映出不同强度人工干扰后的表观量子效率均小于对照组的表观量子效率,而光补偿点均大于对照组,这说明人工干扰后的油松林利用弱光的能力有所降低。中等强度的干扰下,表观量子效率和光补偿点相对对照组变化最小,表明中等强度干扰下油松林相对于低强度干扰和高强度干扰下,油松林具有相对较强的利用弱光的能力。不同强度人工干扰后的光饱和点、暗呼吸、最大净光合速率均大于对照组,而中等强度干扰下的光饱和点和最大净光合速率相对于对照变化最大,分别为 18.5%、51.1%,这说明中等强度干扰下的油松林,具有较强的利用强光能力和较大的光合潜力。相对于对照组,中等强度干扰下的暗呼吸值变化最小(13.6%),这也说明中等强度干扰较利于植物光合产物的积累,利于油松的生长。
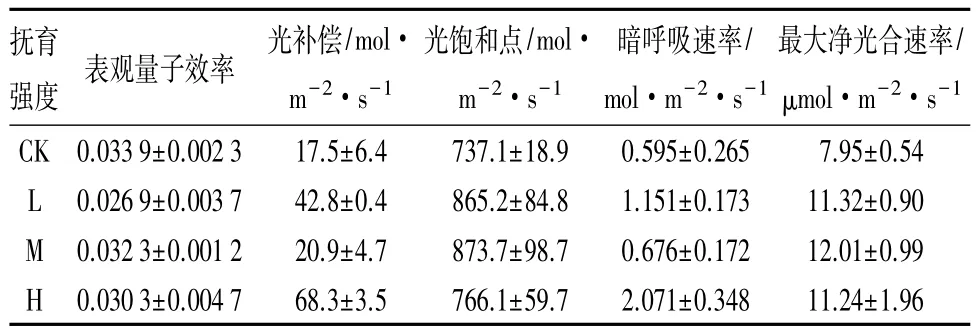
表3 油松不同强度人工干扰下的主要光合生理特征指标
3.4 油松人工林叶绿素质量分数对人工抚育干扰的响应
不同强度人工干扰下的叶绿素a、叶绿素b以及总叶绿素质量分数均小于对照组,尤其以总叶绿素质量分数变化最为明显。叶绿素a质量分数变化波动范围(10.5%~21.1%)小于叶绿素 b(15.2%~26.6%)。相对于对照组,中等强度干扰下的叶绿素a、叶绿素b及总叶绿素质量分数变化波动最小,分别为 10.5%、15.2%和 11.4%。而 m(叶绿素 a) ∶m(叶绿素b)却呈现出相反趋势,即不同强度人工干扰后的m(叶绿素a)∶m(叶绿素b)均大于对照组,其中中等强度干扰下的m(叶绿素a)∶m(叶绿素b)变化最小(4.81%)。这说明中等强度的人工干扰油松针叶相对于低强度人工干扰和高强度人工干扰,具有较强的利用弱光的能力。
3.5 油松人工林其他光合相关参数对人工抚育干扰的响应
由表4分析可知:对照组的气孔导度和蒸腾速率均小于不同强度干扰后的气孔导度和蒸腾速率。其中,中等强度的干扰气孔导度和蒸腾速率均大于低强度干扰和高强度干扰下的气孔导度和蒸腾速率。蒸腾速率与气孔导度成正相关关系,表明气孔导度直接决定蒸腾速率的大小。而胞间CO2摩尔分数值却表现出相反趋势,即干扰后的胞间CO2摩尔分数值均低于对照组。另外,净光合速率与蒸腾速率的相关性显著(P<0.001),因为蒸腾作用过程不仅为光合作用过程提供光合底物——水,还为光合作用过程积累物的运输提供动力。

表4 油松不同强度人工干扰下的其他光合相关参数
4 结论与讨论
4.1 人工抚育干扰对油松光合作用的影响
干扰是影响种群动态变化的重要因素之一[25-26],它可以通过改变生境条件、增加生境异质性等许多复杂的过程使植物群落本身发生结构、动态过程的变化。尤其是对人工林群落的人为干扰,直接影响着群落的发展与稳定。人为干扰的时间、强度等都影响着干扰的结果[27]。本研究中,人工抚育干扰对油松人工林的影响,通过光合特性参数的比较呈现,3种不同强度的人工干扰处理后的光饱和点、光补偿点、暗呼吸速率均高于对照组,羧化效率则相反。这说明人工抚育干扰后的油松林具有较强的利用强光的能力,而对弱光的利用能力有所下降。暗呼吸速率的增加(0.081 ~1.476 mol·m-2·s-1),反映了干扰后的油松林加速了对光合产物的消耗,但是最大净光合速率相对对照组增加范围3.29~4.06 μmol·m-2·s-1,相对消耗量光合产物的积累量增加,人工干扰后更有利于油松人工林的生长。相对于低强度人工干扰和高强度人工干扰,中等强度人工干扰下的油松林具有较高的光饱和点、羧化效率、最大净光合速率,而具有较低的暗呼吸和光补偿点,进一步表明了中等强度干扰下具有较高的利用强光和弱光的能力,对光的适应范围相对较宽,同时具有较低的光合产物消耗以及较高的光合潜力,更利用油松人工林的生长。
作为光合作用的光敏催化剂的叶绿素,与植物光合作用密切相关,其比例和含量是植物对环境适应性的重要标志。叶绿素a主要吸收长波光,叶绿素b以吸收短波光的漫射光和散射光为主。叶绿素a/b值降低,而叶绿素b和总叶绿素质量分数增加,有利于植物对弱光的利用,另外叶绿素质量分数的增加更利于植物对光能捕获,以保证最大限度地进行光合碳积累,这是植物对光照不足适应的重要策略之一[28-34]。本研究结果显示,不同强度人工干扰下的叶绿素a、叶绿素b以及总叶绿素T质量分数相对于对照组,均有所减少,说明随着人工干扰后随着光环境的增强油松对光能的捕获能力有所下降,这与前人研究结果相一致[35-36];而叶绿素 a/b 值随着干扰后光环境的增强有所增加,符合油松耐阴性植物[16]的特性,这与前人对其他植物研究结果相一致[35-36]。其中,中等强度人工干扰下的油松针叶叶绿素a、叶绿素b、总叶绿素质量分数大于低强度和高强度度人工干扰下的值,而叶绿素a/b值相反,这进一步说明了中等程度干扰下的油松针叶相对于低强度和高强度人工干扰下油松针叶,具有较强的利用弱光的能力。
由于蒸腾速率(Tr)=气孔导度(Cond)×水汽压亏缺(VPD),所以在环境条件不变的情况下,气孔导度决定蒸腾速率的变化,都用“蒸腾速率”表示直接反映气孔导度的大小[37]。植物通过调节气孔开闭程度来降低胞间CO2摩尔分数,气孔对Ci很敏感,Ci的增加常伴随着气孔的关闭和 Cond的降低[38]。本研究显示随着干扰强度的增加,Cond、Tr、净光合速率先增加后减小,Ci摩尔分数先减小后增加再减小。有研究表明随着Cond值降低,而Ci值增加时,利于植物对CO2的固定和光能利用效率的提高。但此时的Pn降低,表明植物对光能总量的捕获不足,这是限制植物生长的主要因素[34,39]。虽然光照可以通过增加叶面温度、促进水分蒸发来提高蒸腾速率,但本研究是在控制环境条件如叶室温度、CO2摩尔分数、光照强度等一致的情况下进行的,保证了数据相互间可比性的前提条件,因此,光照可能会通过增加Cond,促进叶片气体交换,加速蒸腾。本研究表明,相对于对照组,中等强度人工干扰下的油松人工林针叶气孔导度值最大,Ci值相对于对照组低7.5%,但光合值却增加21.8%,这表明中等程度干扰下更利于油松生长。
4.2 人工抚育后影响油松光合作用的关键因子
光照是影响植物光合作用的首要因素,光照强度的改变将对植物叶片光合速率以及光合特性参数产生重要影响。研究表明生长在强光环境下的植株,其叶片通常具有较高的光饱和点和最大光合速率,同时也具有较高的呼吸消耗[40-41];而在弱光环境中的生长植株,一般具有较高的表观量子效率,而具有较低的光补偿点和最大光合速率低[42]。本研究显示,人工抚育干扰后影响油松光合作用的关键因子是光合有效辐射,这与前人的研究结果相一致[19]。随着人工干扰强度的增加,日平均光合有效辐射逐渐增大,3种强度人工抚育干扰处理后的光饱和点、光补偿点、暗呼吸均大于对照组,且中强度人工抚育干扰下,光饱和点、最大净光合速率最大,而暗呼吸、表观量子效率相对于对照组变化最小,这说明了一定的光强范围内,光照强度增加利于油松光合作用和生长,这与前人的研究结果基本一致[36,40]。
另外,表3也从另一角度反映了人工抚育之后环境因子对油松光合作用的影响作用并不是独立的,而是相互之间协同作用。光照强度的增加影响整个林分冠层的温度,直接影响植物光合作用蒸腾速率大小,间接影响光合作用过程的酶活性,进而影响光合速率大小。此外,环境温度的升高降低了植物周围空气湿度,减少植物叶片对环境水分的摄取,造成叶片局部的水分胁迫。湿度的增强,加速植物光合作用,进而促进了植物对CO2的吸收转化速率,加速了对周边环境CO2的吸收[43]。分析显示人工干扰后其他环境因子与油松净光合速率之间的相关性不显著,这并不代表其他环境因子对油松光合作用的影响不重要。光合作用是一个错综复杂的过程的,对单因子和双因子的分析研究,只是从该角度去解释单因子或双因子对油松光合作用的影响,而对多因子共同作用下油松光合作用的响应还有待进一步探讨。
不同强度人工干扰下的油松人工林中,大气CO2摩尔分数、大气相对湿度、光合有效辐射、大气温度的日变化均为“单峰”曲线。分析结果显示,人工抚育干扰后,光合有效辐射是限制油松净光合速率的关键因子。中等强度人工干扰下的油松人工林具有相对较高的日均净光合速率。相对于对照组,不同强度人工抚育干扰后的油松针叶具有相对较高的最大净光合速率、光饱和点、光补偿点、暗呼吸,相对较低的表观量子效率。不同强度人工抚育干扰虽然增大了油松针叶对光合产物的消耗,降低了油松针叶对弱光的利用能力,但是增大了对强光的利用能力和光合潜力。中等强度人工干扰下更利于油松光合产物的积累,更利于油松的生长。不同强度人工抚育干扰下的叶绿素a、叶绿素b以及总叶绿素质量分数相对于对照组,均有所减少,而m(叶绿素a)∶m(叶绿素b)值有所增加,表明人工抚育干扰后的油松针叶对弱光的吸收能力有所减低。相对于低强度和高强度人工抚育干扰下油松针叶,中等程度干扰下的油松针叶具有较强的利用弱光的能力。
[1]康宏樟,朱教君,徐美玲.沙地樟子松人工林营林技术研究进展[J].生态学杂志,2005,24(7):799-806.
[2]盛炜彤.杉木林的密度管理与长期生产力研究[J].林业科学,2001,37(5):2-9.
[3]姜志林,叶镜中,周本琳.杉木林的抚育间伐[M].北京:中国林业出版社,1982.
[4]林业部造林经营司主编.杉木林丰产技术[M].北京:中国林业出版社,1982.
[5]吴长山,于先洲,吴延平.樟子松不同强度间伐试验初报[J].吉林林业科技,1998(2):22-23.
[6]姜凤岐.现有防护林合理经营与改造技术研究[M].北京:中国林业出版社,1996.
[7]张水松,陈长发,吴克选,等.杉木林间伐强度试验20年生长效应生物研究[J].林业科学,2005,41(5):56-65.
[8]López B C,Sabate S,Gracia C A.Thinning effects on carbon allocation to fine roots in a Quercus ilex forest[J].Tree Physiology,2003,23:1217-1224.
[9]Fukuzawa K,Shibata H,Takagi K,et al.Effects of clear cutting on nitrogen leaching and fine root dynamics in a cool-temperate forested watershed in northern Japan[J].Forest Ecology and Management,2006,225:257-261.
[10]马履一,李春义,王希群.不同强度间伐对北京山区油松生长及其林下植物多样性的影响[J].林业科学,2007,43(5):1-9.
[11]林娜,刘勇,李国雷,等.抚育间伐对人工林凋落物分解的影响[J].世界林业研究,2010,23(3):44-47.
[12]Noguchi K,Han Q M,Araki M G,et al.Fine-root dynamics in a young Hinoki cypress(Chamaecyparis obtusa)stand for 3 years following thinning[J].Journal of Forest Research,2011,16:284-291.
[13]徐化成.森林学[M].北京:中国农业出版社,1961.
[14]狄晓燕,池喜梅,陈建文,等.5种油松种源光合特性的比较研究[J].植物研究,2012,32(2):165-170.
[15]徐佳佳,张建军,王清玉,等.油松和侧柏的光合蒸腾特性及其与环境因子的关系[J].东北林业大学学报,2011,39(7):15-18.
[16]何平,高荣孚,汪振儒.光状况对油松苗生长和特性的影响[J].生态学报,1993,13(1):91-95.
[17]曹军胜,刘广全.油松光合特性的研究[J].安徽农业科学,2005,33(4):608-609.
[18]张卫强,贺康宁,王正宁,等.光辐射强度对侧柏油松幼苗光合特性与水分利用效率的影响[J].中国水土保持科学,2006,4(2):108-113.
[19]丁晓纲,何茜,李吉跃,等.毛乌素沙地樟子松和油松人工林光和生理特性[J].水土保持研究,2011,18(1):215-219.
[20]Zhang Z A,Zhang M S,Wei R H.The exprimental guide for phytophysiology[M].Beijing:China Agricultural Scientech Press,2004:43-45.
[21]韩忠明,赵淑杰,刘翠晶,等.遮荫对3年生东北铁线莲生长特性及品质的影响[J].生态学报,2011,31(20):6005-6012.
[22]李轩然,刘琪,蔡哲,等.湿地松林叶面积指数测算[J].生态学报,2006,26(12):4099-4105.
[23]师生波,李慧梅,王学英,等.青藏高原几种典型高山植物的光合特性比较[J].植物生态学报,2006,30(1):40-46.
[24]张国平,周伟军.植物生理生态学[M].杭州:浙江大学出版社,1998.
[25]Zhu J J,Liu Z G.A review on disturbance ecology of forest[J].Chinese Journal of Applied Ecology,2004,15(10):1703-1710.
[26]Aubert M,Alard D,Bureau F.Diversity of plant assemblages in managed temperate forests:a case study in Normandy(France)[J].Forest Ecology and Management,2003,175:321-337.
[27]于立忠,朱教君,孔祥文,等.人为干扰(间伐)对红松人工林林下植物多样性的影响[J].生态学报,2006,26(11):3757-3764.
[28]Zheng Zuofang.Characteristics of extreme temperature variation and their response to urbanization in Beijing[J].Scientia Geographica Sinica,2011,31(4):459-463.
[29]Givnish T J,Monigomery R A,Goldstein G.Adaptive radiation of photosynthetic physiology in the Hawaiian Lobeliads:light regimes,static light responses,and wholeplant compensation points[J].American Journal of Botany,2004,91(2):228-246.
[30]Osone Y,Tateno M.Nitrogen absorption by roots as a cause of interspecific variations in leaf nitrogen concentration and photosynthetic capacity[J].Functional Ecology,2005,19:460-470.
[31]Liu Y Q,Sun X Y,Wang Y,et al.Effects of shades on the photosynthetic characteristics and chlorophyll fluorescence parameters of Urticadioica[J].Acta Ecologica Sinica,2007,27(8):3457-3464.
[32]胡启鹏,郭志华,李春燕,等.不同光环境下亚热带常绿阔叶树种和落叶阔叶树种幼苗的也形态和光合生理特征[J].生态学报,2008,28(7):3261-3270.
[33]刁俊明,陈桂珠.盆栽桐花树对不同遮光度的生理生态响应[J].生态学杂志,2011,30(4):656-663.
[34]徐兴利,金则新,何维明,等.不同增温处理对夏蜡梅光合特性和叶绿素荧光参数的影响[J].生态学报,2012,32(20):6343-6353.
[35]Dai Y J,Shen Z G,Liu Y,et al.Effects of shade treatments on the photosynthetic capacity,chlorophyll fluorescence,and chlorophyll content of Tetra stigm a hemsleyanum Dielset Gilg[J].Environmental and Experimental Botany,2009,65:177-182.
[36]王云贺,韩忠明,韩梅,等.遮阴处理对东北铁线莲生长发育和光合特性的影响[J].生态学报,2010,30(24):6762-6770.
[37]王建林,温学发,赵凤华,等.CO2浓度倍增对8种作物叶片光合作用、蒸腾作用和水分利用效率的影响[J].植物生态学报,2012,36(5):438-446.
[38]Jiang Gaoming,Han Xingguo,Lin Guanghui.Response of plant growth to elevated CO2:a review on the chief methods and basic conclusions based on experiments in the external countries in past decade[J].Acta Phytoecologica Sinica,1997,21:489-502.
[39]Farquhar G D,Sharkey T D.Stomatal conductance and photosynthesis[J].Annual Review of Plant Physiology,1982,33:317-345.
[40]霍常富,孙海龙,王政权,等.光照和氮营养对水曲柳苗木光合特性的影响[J].生态学杂志,2008,27(8):1255-1261.
[41]Walters M B,Field C B.Photosynthesis light acclimation in two rainforest piper species with different ecological amp litudes[J].Oecologia,1987,72:449-456.
[42]Sims D A,Seemann J R,Luo Y.The significance of differences in the mechanisms of photosynthetic acclimation to light,nitrogen and CO2for return on investment in leaves[J].Functional Ecology,1998,12:185-194.
[43]许大全.光合作用效率[M].上海:上海科学技术出版社,2002.
猜你喜欢
南方农业·下旬(2022年4期)2022-05-24
山西林业(2021年2期)2021-07-21
中国水土保持科学(2019年6期)2019-04-26
现代园艺(2017年22期)2018-01-19
现代园艺(2017年19期)2018-01-19
现代园艺(2017年21期)2018-01-03
新课程·下旬(2017年7期)2017-08-14
广西林业科学(2016年3期)2016-03-16
西北林学院学报(2015年5期)2015-01-03
亚热带资源与环境学报(2014年3期)2014-01-23
