
浑河上游河岸带土壤微生物数量及酶活性特征
2014-09-13 10:06吴祥云赵雪淞
水土保持通报 2014年1期
孔 涛, 吴祥云, 赵雪淞, 刘 民, 黄 静, 张 丹
(1.辽宁工程技术大学 理学院, 辽宁 阜新 123000; 2.中国科学院 沈阳应用生态研究所,辽宁 沈阳 110016; 3.辽宁工程技术大学 环境科学与工程学院, 辽宁 阜新 123000)
河岸带是指陆地上同河水发生作用的植被区域[1],是河水—陆地交界处的两边, 直至河水影响消失为止的地带[2],是陆地生态系统和水生生态系统的过渡区,具有明显的边缘效应[3]。河岸带具备了植物截流纳污的缓冲带功能和植物根系护土的护岸功能,并且具有强大的生态保护功能,与相邻的生态系统相比,河岸带生态系统具有很强的生物多样性[4-5]。河岸带土壤质量决定了在其上生长的植被量,从而影响了河岸带的生态功能。土壤微生物和土壤酶是土壤生态系统的重要组成成分,其中土壤微生物是土壤生化反应的推动者和参与者,是土壤有机无机复合体的重要组成部分,被认为是表征土壤质量变化最敏感最有潜力的指标[6-7],土壤酶是土壤生物化学过程的主要调节者,活性指示了土壤总生物学活性,表征了土壤的综合肥力特征及养分转化进程,可作为衡量生态系统土壤质量变化的预警和敏感指标[8]。目前,对于浑河流域的研究主要集中在水体污染及修复[9-11]、河岸带对污染物的去除等方面[12],在河岸带土壤微生物和酶活性方面,尚未见报道。本文选取了浑河上游4种典型植被河岸带土壤,探讨土壤微生物数量和土壤酶活性的差异及两者之间的关系,这对了解它们与当地生态环境关系,对研究和保护浑河上游河岸带生态资源具有重要意义。
1 研究区概况与研究方法
1.1 研究区概况
浑河为辽宁省东部河流,源于清原县滚马岭,流经抚顺、沈阳等市,在海城古城子附近纳太子河,向南流至营口市附近入辽东湾,全长415 km,河宽100~200 m,流域面积 1.14×104km2,年径流量3.05×109m3。浑河为不对称水系,东侧支流密集,坡陡谷深,水量丰富;西侧支流很少,水量不大。浑河流域属中温带大陆性季风气候,其特点是春季短回暖快,风大干旱,夏季长而炎热多雨,秋季短干燥凉爽,冬季长且较寒冷多风雪。年温差较大,年平均气温5.3 ℃,此区雨热同期,年均降雨量806.5 mm,多集中在6,7,8月,年蒸发量为1 275 mm。
本试验采样地点地处清原县最南端,以山地为主,坡度 15~35°,海拔 330~1 116 m。最高峰为龙岗山,海拔1 116 m,站区内的主要河流有大苏河、长沙屯河和平岭后河,均发源于龙岗山下,由南向北汇入浑河,是浑河的上游支流,地理坐标为41°51′10″—41°51′13″N,124°54′43″—124°54′55″E。
1.2 研究方法
1.2.1 采样点布置及样品采集 在浑河上游支流大苏河沿岸选择4个典型植被区域取样。样地基本概况见表1。每个典型区域内选择坡度、坡向基本一致的地段设置5个重复样地,每个样地土壤均为棕壤,横向宽4 m,纵向垂直溪流方向沿坡向上,长5 m,设置于河岸植被带内(距离水体4~10 m)。每个样地用土钻按S形取10个土样,深度为0—20和20—40 cm,混匀。

表1 不同植被类型河岸带基本概况
1.2.2 测试项目及方法 参照《土壤理化分析》[13]分别测定土壤的物理、化学性质。土壤密度、孔隙度采用环刀法,pH值采用电位法,有机质采用重铬酸钾法,全氮采用凯氏定氮法,全磷采用钼锑抗比色法,土壤基本理化性质见表2。土壤微生物数量测定方法参照《土壤微生物研究法》[14],细菌、真菌和放线菌数量测定采用最大或然法(MPN),相应的培养基分别采用牛肉膏蛋白胨培养基、马丁氏—孟加拉红培养基加链霉素和改良高氏1号培养基,细菌于36 ℃培养箱内培养24 h,放线菌于28 ℃培养箱内培养3~4 d;真菌于30 ℃培养5~7 d。
土壤酶活性参照关松荫[15]的方法:脲酶采用次氯酸钠比色法,蛋白酶采用茚三酮比色法,蔗糖酶采用3,5—二硝基水杨酸比色法,磷酸酶采用对硝基苯磷酸二钠法,多酚氧化酶采用没食子酸法,过氧化氢酶采用高锰酸钾滴定法。
1.2.3 数据分析 采用 Microsoft Office Excel 2007 软件及SPSS 16.0对数据进行整理及统计分析。
2 结果与分析
2.1 不同植被类型河岸带土壤微生物数量的变化
试验结果表明(表3),土壤细菌、放线菌、真菌和微生物总数量在相同土壤层次大体表现出:灌木草地>次生林>人工松林>玉米地的规律。4种植被类型河岸带土壤细菌数量在灌木草地和次生林中最高,两者之间无显著差异,但都与人工松林和玉米地差异显著,灌木草地土壤表层和深层细菌数量分别达1.057×109和6.52×108cfu/g,比最低的玉米地细菌数量分别高了330.55%和341.07%。表层土壤放线菌数量在次生林和人工松林之间差异不显著,而两者与灌木草地和玉米地之间差异皆显著。深层土壤放线菌数量在次生林、人工松林和玉米地之间差异不显著,三者却都与灌木草地之间具有显著差异,以灌木草地最多,表层和深层分别达7.01×105和5.25×105cfu/g,比最低的玉米地放线菌数量分别高了182.97%和189.46%。
表层和深层土壤真菌数量在人工松林和玉米地之间差异不显著,却都与灌木草地和次生林之间有显著差异,表现出了相同规律。土壤真菌数量以灌木草地最多,表层和深层分别达7.04×104和3.19×104cfu/g,比最低的玉米地真菌数量分别高了557.94%和275.29%。

表2 不同植被类型河岸带土壤理化性质

表3 不同植被类型河岸带土壤微生物数量
4种植被类型河岸带中,土壤微生物均以细菌占绝对优势,表层和深层细菌数量分别占微生物总数量的99.89%~99.94%和99.87%~99.94%,放线菌数量次之,表层和深层分别占0.06%~0.10%和0.05%~0.12%,真菌最少,表层和深层分别占0.003%~0.007%和0.004%~0.006%。
在土壤的不同层次中,4种植被类型河岸带土壤微生物均随着土层的加深而急剧减少,表层(0—20 cm)明显大于深层(20—40 cm),各类微生物数量在表层与深层之间都具有显著差异(p<0.05),表现出非常明显的表聚性。
2.2 不同植被类型河岸带土壤酶活性的变化
研究结果表明(表4),4种植被类型河岸带土壤酶活性大体表现为灌木草地和次生林最高,人工松林次之,玉米地最低。表层和深层土壤蔗糖酶和脲酶活性在4种植被类型河岸带中呈现相同的规律,在灌木草地和次生林中最高,之间无显著差异,但都与落叶松人工林和玉米地之间差异显著,灌木草地表层和深层土壤蔗糖酶分别比最低的玉米地高出369.58%和228.80%,脲酶比玉米地高出133.77%和262.86%。磷酸酶活性在4种植被类型河岸带表层和深层土壤之间均具有显著差异,以灌木草地中最高,比最低的玉米地高出282.63%和130.30%。多酚氧化酶在4种植被类型河岸带表层土壤之间差异均显著,而在深层土壤中人工松林和玉米地之间差异不显著,其活性在灌木草地中最高,表层和深层分别比最低的玉米地高了182.46%和108.57%。表层和深层土壤过氧化氢酶活性在灌木草地和次生林中最高,之间差异不显著,但都与玉米地之间具有显著差异,其在灌木草地的活性比最低的玉米地分别高了82.68%和67.74%。
在土壤的不同层次中,4种植被类型河岸带土壤酶活性表现出了与土壤微生物数量相同的规律,随着土层的加深而急剧减少,表层明显大于深层,各种酶活性在表层与深层之间都具有显著差异(p<0.05),同样表现出非常明显的表聚性。

表4 不同植被类型河岸带土壤酶活性
2.3 土壤微生物数量、酶活性和养分之间的相关性
相关分析表明(表5),细菌数量与蔗糖酶、脲酶、磷酸酶、多酚氧化酶、过氧化氢酶5种酶之间具有显著或极显著的相关关系。放线菌与除了过氧化氢酶之外的其他4种酶具有显著或极显著的相关性,而真菌仅和蔗糖酶、磷酸酶和多酚氧化酶具有显著的相关性。土壤微生物数量与土壤酶活性呈现了较好的正相关性,原因在于土壤酶主要来源于土壤微生物。有机质和总氮含量与细菌数量以及5种酶活性呈显著或极显著的相关性,总磷含量与5种酶活性呈显著或极显著的相关性。
土壤养分含量与土壤细菌数量和土壤酶活性之间呈现显著正相关,表明土壤养分的增加会促进微生物的生长繁殖,同时提高了土壤酶的底物供应水平,刺激了酶活性的提高。5种酶活性之间也呈现了极显著的正相关关系,表明酶在促进各自底物分解的同时,其活性既受底物浓度高低影响,也受到其他酶的影响。
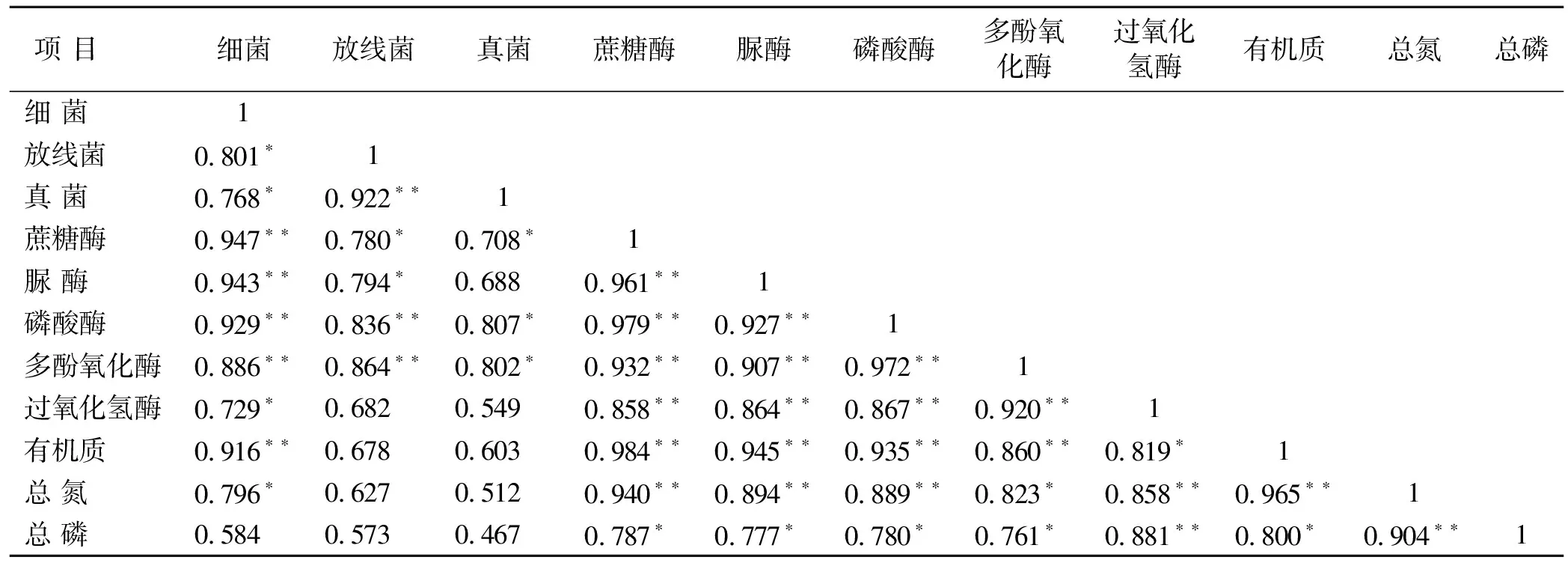
表5 土壤微生物数量、酶活性与土壤主要养分间的相关系数
3 讨 论
土壤微生物是土壤系统中极其重要和最为活跃的部分,在土壤有机物的降解、矿化释放无机元素供其他生物吸收,营养物质转化如碳氮循环、系统稳定性及抗干扰能力中居于支配地位,控制着土壤生态系统功能的关键过程[16-17]。高质量的土壤应具有稳定的微生物群落组成和结构、高微生物数量及良好的微生物活性。本文的4种植被类型河岸带土壤微生物中细菌占绝对优势,放线菌次之,真菌最少,表明在河岸带条件下,细菌的繁殖力、竞争力、土壤养分转化能力以及抗干扰能力强于其他类群,放线菌与真菌数量上不及细菌,但其绝对数量也较多,反映其对于河岸带条件下的物质循环和系统稳定也具有一定的作用,这与陆梅等[18]研究几种针叶林中的微生物数量的结果,吕可等[19]研究农田土壤微生物数量的结果是一致的。4种植被类型河岸带同层土壤微生物数量在灌木草地和次生林中最多,人工松林次之,玉米地最少。微生物的生长代谢是多维环境因子综合作用的结果,土壤微生物的营养主要来源于植物残体,在有机质丰富的土壤中,微生物数量相应偏多,而在缺乏有机质的土壤中其数量较少[20]。灌木草地由于植被类型多样且极为丰富,地上部分生物量大,凋落物非常丰富且易于分解,使其土壤表层有机质含量高,表层达到了92.69 g/kg,且地下根系发达,密集于土壤表层,根系的分泌物促进了微生物的生长繁殖,因而,微生物总数最高,这与赵聪等[21]研究森林次生演替群落的土壤微生物数量的结果是一致的。次生林内枯枝落叶丰富,且成分较复杂,使其林内表层土壤有机质丰富,表层达到了98.42 g/kg,并且土壤疏松,透气性好,为微生物生长提供了充足的养分和良好的生存环境,因而,次生林中微生物数量也非常多。人工松林中地面积累的枯枝落叶较少,且其针叶不易分解,表层土壤有机质含量较低,土壤孔隙度较低,通气性不如灌木草地和次生林,故微生物数量不如前两者。玉米地由于几乎没有积累枯枝落叶层,因而土壤表层有机质含量最低,微生物数量也最少。细菌、放线菌和真菌的数量分布都表现出了明显的表聚效应,这是由于土壤表层有机质含量较深层丰富,且通气状况良好,有利于微生物的生长,同时由于深层土壤温度较低,不利于微生物的繁殖,这与许多学者的研究结果一致[21-22]。
土壤酶是土壤组分中最活跃的有机成分之一,是土壤生物化学过程的主要调节者, 对土壤中碳、氮、磷、硫等元素的循环与迁移有着重要作用[23],主要来源于微生物[24]。土壤蔗糖酶、脲酶、磷酸酶活性能够表征土壤碳、氮、磷等养分的循环状况。蔗糖酶是表征土壤生物学活性的重要水解酶,与土壤中的腐殖质、水溶性有机质和土壤微生物的数量有关,常用于评价土壤熟化程度和肥力水平[25]。灌木草地和次生林由于枯枝落叶层厚,土壤有机质含量高,同时人为对土壤的扰动少,改善了土壤环境,土壤微生物数量最多,使得土壤蔗糖酶活性最高,而落叶松人工林和玉米地有机质含量少,人工扰动却最多,微生物数量少,尤以玉米地为甚,因此其蔗糖酶活性远小于前两者。脲酶是决定土壤氮素转化的关键,与土壤的微生物数量、有机质含量、氮含量呈正相关[26]。灌木草地和次生林由于土壤有机质、全氮含量高,土壤微生物数量也最多,从而显著提高了脲酶水平,落叶松人工林次之,玉米地最少。磷酸酶活性受土壤坚实度影响较小,但受土壤通气状况影响很大。灌木草地和次生林由于减少了土壤翻动,根系在土壤中盘根错节,以及枯枝落叶覆盖等原因,改善了土壤通气,使土壤中水分、空气和热量比例较为合理,从而使磷酸酶活性提高,其中灌木草地效果最好,落叶松人工林和玉米地由于人为对土壤的扰动,使得土壤通气状况及水热比例不合适,因而磷酸酶活性较低,以玉米地为最低。多酚氧化酶能把土壤中芳香族化合物氧化成醌,醌与土壤中其他物质反应生成有机质和色素,从而完成对芳香族物质的转化,可以表征对土壤中污染物的净化能力[27]。多酚氧化酶活性在灌木草地中最高,接着依次为次生林、人工松林,玉米地最低,表明灌木草地土壤对污染水体的修复能力是最强的,可将吸附在土壤中浑河水污染物转化,而玉米地则对浑河水体的修复能力最低。过氧化氢酶的作用在于破坏对生物体有毒的过氧化氢,可以表示土壤氧化过程的强度,与土壤有机质转化速度密切相关,其活性与土壤有机质含量和微生物数量有关[25]。灌木草地和次生林由于土壤有机质含量最高,同时适宜的营养、水热条件有利于微生物的生长,从而提高了土壤中的过氧化氢酶活性,保护了其上植物免受过氧化氢的毒害,表明这两种河岸带植被土壤对河流污水具有一定的清除能力,而玉米地的土壤过氧化氢酶活性则最低。4种植被类型河岸带土壤各种酶活性均具有明显的土体层次差异,在表层活性最高,这主要是由于表层积累了较多的枯落物和腐殖质,有机质含量高,营养状况和水热条件好,微生物的代谢能力强,使土壤表层聚积了较高的土壤酶活性。
4 结 论
由于灌木草地和次生林这两种河岸带植被类型土壤具有最高的微生物数量和土壤酶活性,有利于碳氮磷转化和浑河水质的净化,土壤质量高,是浑河生态型发展的很好的河岸带形式。而人工松林和玉米地土壤质量较低,对污染物去除能力较差,尤其玉米地的施肥措施会污染浑河水质,不适宜作为河岸带的植被类型,所以应减少对河岸带植被的人为扰动。
[参考文献]
[1]Campbell A G, Franklin J F. Riparian vegetation in Oregon’s Western Cascade Mountains: Composition, biomass, and autumn phenology[M]. Seattle: University of Washington Press, 1979:4-5.
[2]Gregory S V, Swanson F J, Mckee W A, et al. An ecosystem perspective of riparian zones[J]. Bioscience, 1991,41(8): 540-551.
[3]陈吉泉.河岸植被特征及其在生态系统和景观中的作用[J].应用生态学报,1996,7(4):439-448.
[4]Naiman R J, Decamps H, Pollock M. The role of riparian corridors in maintaining regional biodiversity[J]. Ecological Applications, 1993,3(2):209-212.
[5]夏继红,严忠民.生态河岸带研究进展与发展趋势[J].河海大学学报:自然科学版,2004,32(3):252-255.
[6]刘占锋,傅伯杰,刘国华,等.土壤质量与土壤质量指标及评价[J].生态学报,2006,26(3):901-913.
[7]薛立,陈红跃,邝立刚.湿地松混交林地土壤养分、微生物和酶活性的研究[J].应用生态学报,2003,14(1):157-159.
[8]万忠梅,宋长春.土壤酶活性对生态环境的响应研究进展[J].土壤通报,2009,40(4):951-956.
[9]梁雷,刘峰,何丽,等.浑河上游不同类型农户氮负荷特征[J].生态学杂志,2011,30(10):2315-2322.
[10]张鸿龄,孙丽娜,罗庆,等.浑河流域水体污染的季节性变化及来源[J].生态学杂志,2011,30(1):119-125.
[11]张艳强,安立会,郑丙辉,等.浑河野生鲫鱼体内重金属污染水平与金属硫蛋白基因表达[J].生态毒理学报,2012,7(1):57-64.
[12]阎丽凤,石险峰,于立忠,等.沈阳地区河岸植被缓冲带对氮、磷的削减效果研究[J].中国生态农业学报, 2011,19(2):403-408.
[13]中国科学院南京土壤研究所.土壤理化分析手册[M].上海:上海科学技术出版社,1978:62-93.
[14]中国科学院南京土壤研究所.土壤微生物研究方法[M].北京:科学出版社,1985:34-58.
[15]关松荫.土壤酶及其研究法[M].北京:中国农业出版社,1986:157-241.
[16]Stenberg B. Monitoring soil quality of arable land: Microbiological indicators[J]. Acta Agriculturae Scandinavica(Section B): Soil and Plant Science, 1999,49(1):1-24.
[17]Winding A, Hund-Rinke K, Rutgers M. The use of microorganisms in ecological soil classication and assessment concepts[J]. Ecotoxicology and Environmental Safety, 2005,62(2):230-248.
[18]陆梅,卫捷,韩智亮.滇池西岸4种针叶林的土壤微生物与酶活性[J].东北林业大学学报,2011,39(6):56-59.
[19]吕可,潘开文,王进闯,等.花椒叶浸提液对土壤微生物数量和土壤酶活性的影响[J].应用生态学报,2006,17(9):1649-1654.
[20]郑诗樟,肖青亮,吴蔚东,等.丘陵红壤不同人工林型土壤微生物类群、酶活性与土壤理化性状关系的研究[J].中国生态农业学报,2008,16(1):57-61.
[21]赵聪,李勇,杨红军. 缙云山森林次生演替群落土壤微生物、酶活性和养分的研究[J].中国农学通报,2012,28(4):46-50.
[22]漆良华,张旭东,周金星,等.湘西北小流域不同植被恢复区土壤微生物数量、生物量碳氮及其分形特征[J].林业科学,2009,45(8):14-20.
[23]Maxm C, Wood M, Jarvis S C. A microplate flurimetric assay for the study of enzyme diversity in soils[J]. Soil Biology and Biochemistry, 2001,33(12):1633-1640.
[24]Zornoza R, Guerrero C, Mataix-Solera J, et al. Assessing air-drying and rewetting pretreatment effect on some soil enzyme activities under Mediterranean conditions[J]. Soil Biology and Biochemistry, 2006,38(8):2125-2134.
[25]王芸,韩宾,史忠强,等.保护性耕作对土壤微生物特性及酶活性的影响[J].水土保持学报,2006,20(4):120-122.
[26]杨招弟,蔡立群,张仁陟,等.不同耕作方式对旱地土壤酶活性的影响[J].土壤通报,2008,39(3):514-517.
[27]丁克强,骆永明,刘世亮,等.黑麦草对菲污染土壤修复的初步研究[J].土壤,2002,4(4):233-236.
猜你喜欢
环境保护与循环经济(2021年12期)2021-03-16
青年生活(2020年31期)2020-10-14
学生天地(2020年17期)2020-08-25
乐活老年(2019年2期)2019-04-20
水利规划与设计(2018年1期)2018-01-31
现代农业研究(2017年11期)2018-01-12
水利技术监督(2017年6期)2017-12-19
辽宁林业科技(2017年4期)2017-06-22
辽宁林业科技(2017年4期)2017-06-22
现代园艺(2017年20期)2017-02-03
