
转Cry1Ac+Cry2Ab棉花生长势及其对棉田节肢动物物种丰富度的影响
2014-08-30 06:05:30雒珺瑜吕丽敏王春义朱香镇李春花崔金杰
生物安全学报 2014年4期
雒珺瑜, 张 帅, 吕丽敏, 王春义, 朱香镇, 李春花, 崔金杰
中国农业科学院棉花研究所,棉花生物学国家重点实验室,河南 安阳 455000
转基因在给人类带来巨大经济利益的同时,食品与环境安全的问题也备受社会关注。其中转基因棉花因大面积种植而显得尤为重要,其环境安全性问题是国内外学者关注的焦点和热点,也是需要长期研究的内容(左娇等,2013)。
转Bt基因抗虫棉对棉铃虫HelicoverpaarmigeraHübner具有良好的控制作用,它的种植减少了化学杀虫剂的使用,降低了环境污染和人畜中毒事件,大大提高了农民的经济效益(马慧等,2009; 苏宏华等,2010)。但任何一种作物都遭受多种害虫危害,一个抗虫基因不能抵御所有害虫,而且不同类型的基因其抗虫机制、杀虫作用和杀虫谱也不尽相同。单个转Bt基因棉花随着种植时间的推移和种植面积的扩大,棉铃虫对其抗性上升的风险也在逐年加大(梁革酶等,2000; 徐广等,2002; Bashir,2014),而将2个或2个以上抗虫基因转入同一植物,可使受体植物获得多机制的抗虫性,不仅可进一步加强转基因植物的抗虫能力,拓宽其抗虫谱,并有助于延缓害虫产生耐受性,从而延长转基因棉花的使用寿命(McGaugheyetaletal.,1992; Tabashniketal.,1997)。
本文以中国农科院棉花研究所生物技术研究室新转化成功的转Cry1Ac+Cry2Ab棉花为试验材料,以转单个Cry1Ac基因棉花和非转基因棉花为对照,从棉花的生长势及其对棉田节肢动物物种丰富度的影响系统进行了研究,为新型转基因棉花的生产应用价值进一步进行评估,同时为转基因棉花环境安全评价提供科学数据。
1 材料与方法
1.1 试验材料
639020为转Cry1Ac+Cry2Ab棉花,由中国农业科学院棉花研究所生物技术研究室提供;中棉所41为转Cry1Ac棉花,中棉所49为非转基因棉花,均由中国农业科学院棉花研究所遗传育种研究室提供。
1.2 试验方法
1.2.1 试验设计 试验在中国农业科学院棉花研究所试验农场进行,棉花播种时间为4月28日。试验共设3个处理:转Cry1Ac+Cry2Ab棉田、Cry1Ac棉田和非转基因棉田;每个品种3个小区,每个小区面积122 m2,小区间随机排列,共9个小区;棉花种植株距0.27 m,行距0.80 m;棉花全生育期不进行化学防治,其他农事操作按常规棉田操作和管理。
1.2.2 棉花生长势和产量构成参数调查 叶绿素:在棉花生长的苗期(05-27)、蕾期(06-20)和花铃期(07-24)采集棉花顶部第1片完全展开叶,每处理每小区随机采集5片叶片,室内通过SPAD测定仪进行检测。
比叶面积:在棉花生长的苗期(05-27)、蕾期(06-20)和花铃期(07-24)采集棉花顶部第1片完全展开叶,每处理每小区随机采集5片叶片,室内测定叶面积,然后将叶片置于108 ℃杀青0.5 h,再于80 ℃烘12 h后,叶片干重称重;计算比叶面积。
棉花生长势和产量构成参数:在棉花生长的初蕾期(06-20)、盛蕾期(07-24)和花铃期(08-18),分别测定棉株株高,调查棉花果枝数、蕾数、幼铃数、成铃数和脱落蕾铃数,每小区选取3个点,每点顺行连续测定和调查10株棉花。
1.2.3 昆虫群落调查方法 试验采用对角线五点取样方法(中华人民共和国农业部,2007),分别在二代、三代和四代棉铃虫发生高峰期,调查不同棉田主要害虫及其天敌的种群数量,每个棉铃虫发生世代每隔1 d调查1次,共计调查3次,每次每小区调查5个样点,每个样点连续调查2行20株棉花,共100株。
1.3 统计分析
对每个棉铃虫发生世代调查的每种节肢动物的3次数值进行平均,以非转基因棉田和转Cry1Ac基因棉田每种主要节肢动物每世代出现的个体平均值占该世代调查到的节肢动物个体总数的比率代表该节肢动物的丰度。运用SPSS 17.0软件对所有数据进行统计分析,采用单因素方差分析(One-way analysis,ANO-VA)和Duncan′s差异显著性分析,检验同一时期不同小区内棉花生长势、产量构成参数和节肢动物物种丰富度的差异。
2 结果与分析
2.1 对棉花生长势和产量构成参数的影响
2.1.1 叶绿素含量 3种棉田不同时期棉花叶片叶绿素含量如表1。苗期(05-27)、蕾期(06-20)和花铃期(07-24)3种棉花顶部叶片叶绿素含量在同一时期变化无显著差异;随着棉花生长,顶部叶片的叶绿素含量呈逐渐下降的趋势。

表1 3种棉花叶片叶绿素含量Table 1 Content of chlorophyll of 3 studied types of cotton
数据为平均数±SD。大写字母表示差异极显著(P<0.01),小写字母表示差异显著(P<0.05)。
Values are means±SD. Captical letters show highly extremely significant differences(P<0.01), and small letters show significant differences (P<0.05) .
2.1.2 比叶面积 由表2可知,与非转基因棉花和转Cry1Ac基因棉花相比,苗期(05-27)转Cry1Ac+Cry2Ab棉花比叶面积分别降低5.6%和22.5%,前者差异不显著,后者差异显著;蕾期(06-20)转Cry1Ac+Cry2Ab棉花的比叶面积分别增加9.6%和5.2%,差异均不显著;花铃期(07-24)转Cry1Ac+Cry2Ab棉花的比叶面积分别增加5.7%和62.9%,前者差异不显著,后者差异达极显著水平。

表2 3种棉花叶片比叶面积 Table 2 Specific leaf areas of 3 studied types of cotton
数据为平均数±SD。大写字母表示差异极显著(P<0.01),小写字母表示差异显著(P<0.05)。
Values are means±SD. Captical letters show highly extremely significant differences(P<0.01), and small letters show significant differences (P<0.05) .
2.1.3 生长势及产量构成参数 3种棉田不同时期棉花生长势和产量构成参数见表3。蕾期(06-20)和花铃期(07-24)转Cry1Ac+Cry2Ab棉花株高、主茎叶片数、果枝数和蕾数均与非转基因棉花和转Cry1Ac基因棉花相当,差异不显著。花铃期(07-24)成铃数、幼铃数和脱落率均与转Cry1Ac棉相当,但成铃数和幼铃数分别比非转基因棉花增加395.6%和211.3%,差异达极显著水平;蕾铃脱落数较非转基因棉花降低83.0%,差异显著。盛铃期(08-18)转Cry1Ac+Cry2Ab棉花主茎叶片数、果枝数和成铃数均与非转基因棉花和转Cry1Ac基因棉花相当,差异不显著;蕾数和幼铃数与转Cry1Ac基因棉花相当,差异不显著,与非转基因棉花相比,蕾数增加63.0%,幼铃数减少69.3%;蕾铃脱落数均较非转基因棉花和转Cry1Ac基因棉花降低51.0%和35.9%,差异均达到显著水平。

表3 3种棉花田间生长势及产量构成参数Table 3 Growth potential and yield component parameters of 3 studied types of cotton
数据为平均数±SD。大写字母表示差异极显著(P<0.01),小写字母表示差异显著(P<0.05)。
Values are means±SD. Captical letters show highly extremely significant differences(P<0.01), and small letters show significant differences (P<0.05) .
2.2 对棉田节肢动物物种丰富度的影响
2.2.1 靶标害虫 棉铃虫卵:二代、三代和四代棉铃虫发生期,3种棉田棉铃虫落卵量丰度如图1所示。3种棉田落卵量在二代棉铃虫发生期种群丰度较高,其次是三代,四代棉铃虫发生期落卵丰度最低;与非转基因棉田相比,二代棉铃虫发生期转Cry1Ac+Cry2Ab基因棉田棉铃虫落卵量丰度提高了76.4%,差异达显著水平;与转Cry1Ac基因棉田相比,转Cry1Ac+Cry2Ab基因棉田棉铃虫落卵量丰度降低了8.9%,差异不显著;三代棉铃虫和四代棉铃虫发生期,3种棉田落卵量丰度均较低,均未达到显著差异。
棉铃虫幼虫:二代、三代和四代棉铃虫发生期3种棉田棉铃虫幼虫丰度如图2所示。3种棉田棉铃虫幼虫在二代棉铃虫发生期种群丰度较高;与非转基因棉田和转Cry1Ac基因棉田相比,二代棉铃虫发生期转Cry1Ac+Cry2Ab基因棉田棉铃虫幼虫丰度分别降低了49.2%和26.19%,前者差异显著,后者差异不显著;三代棉铃虫和四代棉铃虫发生期,3种棉田棉铃虫幼虫丰度均较低,均未达到显著差异。
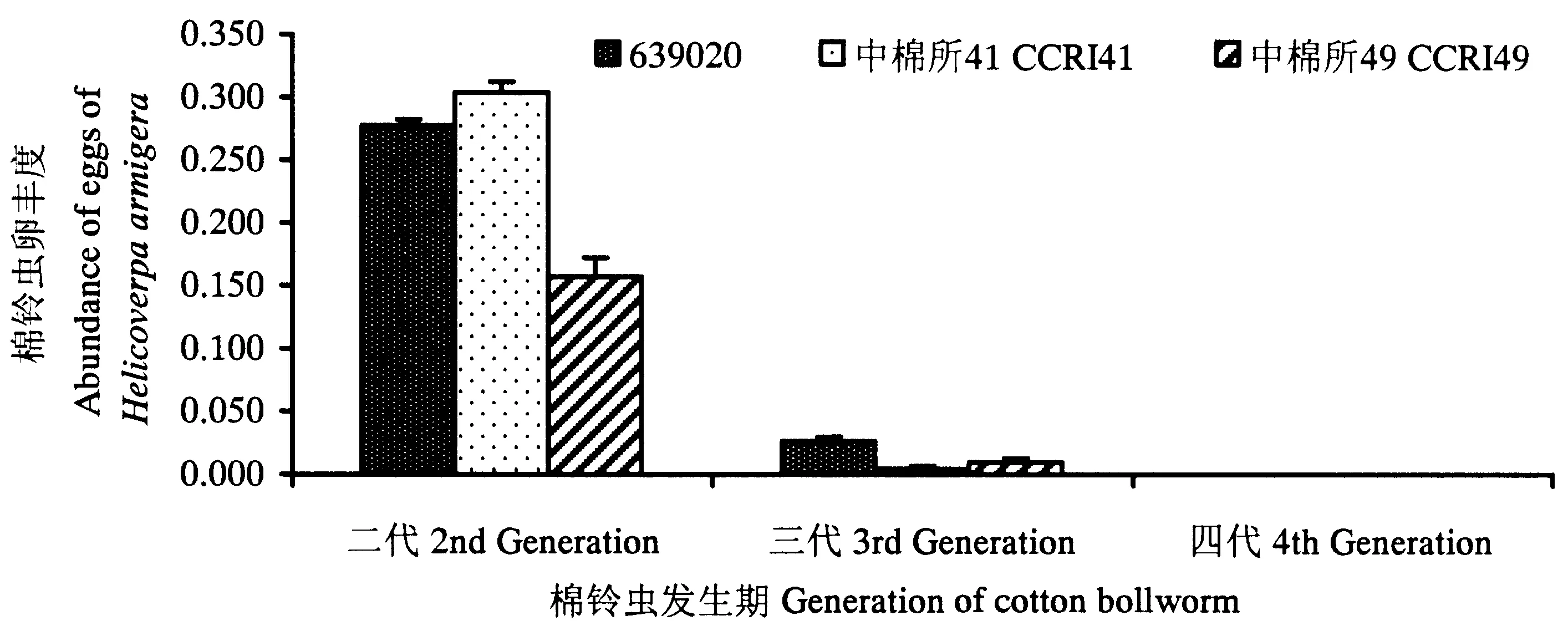
图1 二代、三代和四代棉铃虫发生期三种棉田棉铃虫落卵量丰度Fig.1 The species abundance of eggs of cotton bollworm during different generation in the fields of the 3 studied types of cotton

图2 二代、三代和四代棉铃虫发生期3种棉田棉铃虫幼虫数量丰度Fig.2 The species abundance of larvae of cotton bollworm during different generation in the fields of the 3 studied types of cotton
2.2.2 刺吸性害虫 棉蚜:二代、三代和四代棉铃虫发生期3种棉田棉蚜丰度如图3所示。3种棉田棉蚜在四代棉铃虫发生期种群丰度最高,其次是三代和二代;与非转基因棉田和转Cry1Ac基因棉田相比,二代棉铃虫发生期转Cry1Ac+Cry2Ab基因棉田棉蚜丰度分别提高了10.3%和32.3%,差异均不显著;三代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田棉蚜丰度分别降低了48.6%和-0.9%,前者差异显著,后者差异不显著;四代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田棉蚜丰度分别降低了3.0%和-30.4%,差异均不显著。
棉蓟马:二代、三代和四代棉铃虫发生期3种棉田棉蓟马丰度如图4所示。二代和三代棉铃虫发生期,3种棉田棉蓟马种群丰度较高,四代棉铃虫发生期,种群丰度较低;与非转基因棉田和转Cry1Ac基因棉田相比,二代棉铃虫发生期转Cry1Ac+Cry2Ab基因棉田棉蓟马丰度分别升高了1.4%和11.9%,差异均不显著;三代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田棉蓟马丰度比常规棉田增加113.5%,比转Cry1Ac基因棉田降低5.7%,前者差异极显著,后者差异不显著;四代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田棉蓟马丰度均降低了79.6%,差异均达显著水平。
烟粉虱Bemisiatabaci(Gennadius):二代、三代和四代棉铃虫发生期三种棉田烟粉虱丰度如图5所示。四代棉铃虫发生期,3种棉田烟粉虱种群丰度最高,二代和三代棉铃虫发生期种群丰度较低;与非转基因棉田和转Cry1Ac基因棉田相比,二代棉铃虫发生期转Cry1Ac+Cry2Ab基因棉田烟粉虱丰度均降低了350.0%,差异极显著;三代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田烟粉虱丰度分别增加17.3%和50%,前者差异不显著,后者差异显著;四代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田烟粉虱丰度分别降低了-4.3%和3.1%,差异均未达显著水平。

图3 二代、三代和四代棉铃虫发生期3种棉田蚜虫数量丰度Fig.3 The species abundance of cotton aphids during different generation in the fields of the 3 studied types of cotton

图4 二代、三代和四代棉铃虫发生期3种棉田蓟马丰度Fig.4 The species abundance of thrips during different generation in the fields of the 3 studied types of cotton

图5 二代、三代和四代棉铃虫发生期3种棉田烟粉虱丰度Fig.5 The species abundance of B.tabaci during different generation in the fields of the 3 studied types of cotton
绿盲蝽Llyguslucorμm(Meyer-Dür.):二代、三代和四代棉铃虫发生期3种棉田绿盲蝽丰度如图6所示。三代棉铃虫发生期,3种棉田绿盲蝽种群丰度较高,二代和四代棉铃虫发生期种群丰度相对较低;与非转基因棉田和转Cry1Ac基因棉田相比,二代棉铃虫发生期转Cry1Ac+Cry2Ab基因棉田绿盲蝽丰度均降低了50.0%和0.00%,差异均不显著;三代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田绿盲蝽丰度分别降低57.1%和40.0%,差异不显著;四代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田绿盲蝽丰度分别降低了71.4%和50.0%,前者差异显著,后者差异不显著。
2.2.3 天敌 龟纹瓢虫Propylaeajaponica(Thunberg):二代、三代和四代棉铃虫发生期3种棉田龟纹瓢虫丰度如图7所示。3种棉田龟纹瓢虫在二代棉铃虫发生期时发生数量最多,四代棉铃虫发生期次之,三代棉铃虫发生期种群丰度最低;与非转基因棉田和转Cry1Ac基因棉田相比,二代棉铃虫发生期转Cry1Ac+Cry2Ab基因棉田龟纹瓢虫丰度均增加了131.3%和94.7%,差异均达显著水平;三代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田龟纹瓢虫丰度分别升高-33.3%和50.0%,差异不显著;四代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田烟粉虱丰度分别增加75.0%和55.6%,前者差异显著,后者差异不显著。

图6 二代、三代和四代棉铃虫发生期3种棉田绿盲蝽丰度Fig.6 The species abundance of L.lucorum during different generation in the fields of the 3 studied types of cotton
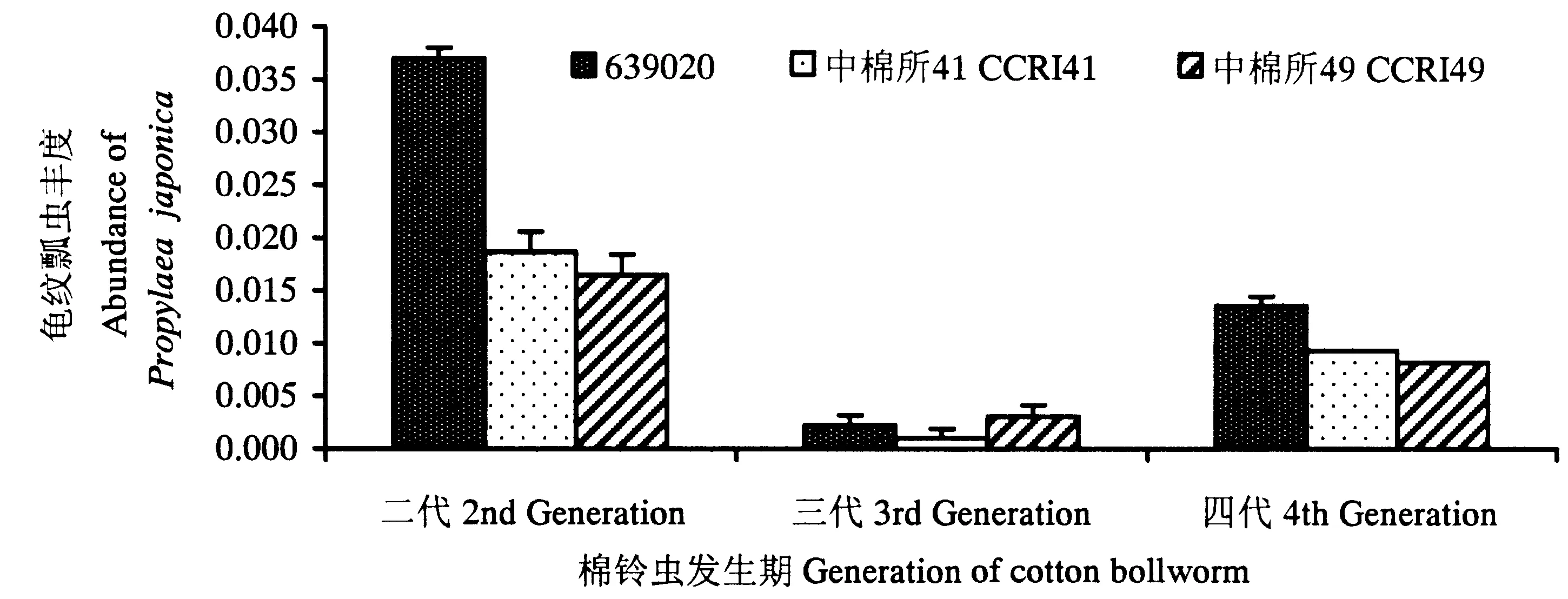
图7 二代、三代和四代棉铃虫发生期棉田龟纹瓢虫丰度Fig.7 The species abundance of P.japonica during different generation in the fields of the 3 studied types of cotton
草间小黑蛛Erigonidiumgraminicolum(Sundevall):二代、三代和四代棉铃虫发生期3种棉田草间小黑蛛丰度如图8所示。3种棉田草间小黑蛛在二代和四代棉铃虫发生期发生数量较多,三代棉铃虫发生期种群丰度相对较低;与非转基因棉田和转Cry1Ac基因棉田相比,二代棉铃虫发生期转Cry1Ac+Cry2Ab基因棉田草间小黑蛛丰度在3种棉田种群丰度相当;三代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田草间小黑蛛丰度分别降低10.0%和25.0%,差异不显著;四代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田草间小黑蛛丰度分别降低12.5%和41.7%,差异均未达显著水平。
草蛉:二代、三代和四代棉铃虫发生期3种棉田草蛉丰度如图9所示。3种棉田草蛉在三代棉铃虫发生期发生数量较多,二代和四代棉铃虫发生期种群丰度相对较低;与转Cry1Ac基因棉田相比,二代棉铃虫发生期转Cry1Ac+Cry2Ab基因棉田草蛉丰度降低了62.5%,差异显著,比非转基因棉田增加了36.4%,差异不显著;三代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田草蛉丰度较非转基因棉田和转Cry1Ac基因棉田分别降低38.2%和15.0%,差异均不显著;四代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田草蛉丰度较非转基因棉田增加62.5%,较转Cry1Ac基因棉田降低55.2%,差异均达显著水平。
小花蝽:二代、三代和四代棉铃虫发生期3种棉田小花蝽丰度如图10所示。3种棉田小花蝽在二代棉铃虫发生期发生数量较多,三代和四代棉铃虫发生期种群丰度相对较低;与非转基因棉田和转Cry1Ac基因棉田相比,二代棉铃虫发生期转Cry1Ac+Cry2Ab基因棉田小花蝽丰度分别降低了16.7%和14.6%,差异均不显著;三代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田小花蝽丰度较非转基因棉田和转Cry1Ac基因棉田分别增加了16.7%和23.5%,差异均不显著;四代棉铃虫发生期,转Cry1Ac+Cry2Ab基因棉田小花蝽丰度较非转基因棉田增加8.3%,较转Cry1Ac基因棉田降低43.5%,差异均未达显著水平。

图8 二代、三代和四代棉铃虫发生期棉田草间小黑蛛丰度Fig.8 The species abundance of E.graminicola during different generation in the fields of the 3 studied types of cotton

图9 二代、三代和四代棉铃虫发生期棉田草蛉丰度Fig.9 The species abundance of lacewing during different generation in the fields of the 3 studied types of cotton

图10 二代、三代和四代棉铃虫发生期棉田小花蝽丰度Fig.10 The species abundance of Orius during different generation in the fields of the 3 studied types of cotton
3 讨论
转基因棉花的环境安全性研究包括对靶标害虫的影响、对棉田生物多样性的影响、生存竞争能力和基因漂移4个方面,每个方面具体的研究内容涉及多方位、多角度。基因漂移的研究结果对一类植物均可通用,如转Bt基因棉花的基因漂移主要是由虫媒引起,而风媒对其产生的作用比较弱(贺娟等,2013);朱家林等(2013)通过网室内试验,发现定向风基因漂移频率在6.4 m处漂移频率为33.33%,最远漂移距离为25.6 m;非定向风基因漂移频率在12.8 m为36.67%,最远漂移距离为36 m。
棉花生长势对棉田节肢动物群落尤其是棉田植食性昆虫的影响较大,可为其提供食物资源,生长势弱,其食物资源少且质量低,植食性昆虫会随之改变,从而改变食物链上的其他昆虫的丰富度或多样性。
本文从棉花叶片叶绿素含量、比叶面积、株高、主茎叶片数和蕾数、成铃数、幼铃数和蕾铃脱落数等方面系统研究了不同时期转Cry1Ac+Cry2Ab基因棉花的生长势和产量构成参数,表明转新型Cry1Ac+Cry2Ab基因棉花的生长势从生存竞争能力方面考虑其没有较强的竞争优势,从生物安全角度考虑该试验材料在本文的研究中没有变为超级杂草的风险,这与张兴华(2012a)的研究结果一致;马小艳等(2013a、2013b)在抗病、抗除草剂棉花试验研究中也得到相同结论。转Cry1Ac+Cry2Ab基因棉花在目标性状方面发挥出更优的产量性状,可达到转外源基因的目的,为进一步在生产上推广应用奠定了基础,与张兴华等(2012a、2012b)、李捷等(2013)、乔艳艳等(2013)、雒珺瑜等(2014a、2014b)研究结果基本一致。
转Cry1Ac+Cry2Ab基因棉田节肢动物的物种丰富度表现出,该新型基因棉花对靶标害虫棉铃虫具有良好的控制作用,这与雒珺瑜等(2014a)在对鳞翅目害虫小地老虎的研究不完全一致,其原因在于本文研究的是鳞翅目害虫中的靶标害虫棉铃虫,而雒珺瑜等(2014a)是对鳞翅目非靶标害虫的研究,虽然都是鳞翅目,但Cry1Ac+Cry2Ab基因是有特异性和专一性的,对非靶标鳞翅目虽然有一定的影响,但具体情况不完全和棉铃虫一致,这和李海强(2012)研究抗虫抗除草剂对棉铃虫和小弟老虎的研究结论基本一致;对非靶标害虫和主要天敌的物种丰富度总体上没有明显的影响,表明该新型转基因棉花材料在本文的研究中对棉田节肢动物物种没有负面的影响,这与雒珺瑜等(2011a、2011b、2012)的研究结果一致。
植物的比叶面积是反映植物生长势和氮代谢的重要指标,比叶面积越大,生长势和氮代谢越强。王香河(2003)在研究中发现,棉花在不同生长时期,由于生长环境和自身生长发育的变化,各有不同的补偿效应。本文研究结果中,转Cry1Ac+Cry2Ab基因棉花在生长前期生长弱,氮代谢差,比叶面积低;而在生长后期,由于抗虫性较好、生长旺盛,表明其补偿能力强,生长势和氮代谢均有所提高。
棉花生长势的强弱,直接影响棉田植食性害虫的生长和繁衍。本文对节肢动物物种丰富度的研究中,个别时期有些害虫或天敌物种丰富度的变化可能与该新型转基因棉花的生长势有关。生长势弱,植食性害虫丰富度降低,引起食物链上的天敌丰富度降低;当棉花生长势增强,叶片、蕾铃增多时,可以为棉田植食性害虫提供更多的食物资源,从而其食物链上的天敌昆虫也跟随增多。因此其对环境安全方面的研究需长期跟踪,并可以考虑从建立模型等方面进一步深入研究。
贺娟, 朱威龙, 朱家林, 潘李隆, 张青文, 刘小侠. 2013. 风、蜜蜂因素对转Cry1Ac基因棉花花粉介导的基因漂移的影响. 棉花学报, 25(5): 453-458.
李海强, 李号宾, 王冬梅, 丁瑞丰, 汪飞, 阿克旦·吾外士, 徐遥, 刘建. 2012. 转基因抗棉铃虫抗草甘膦棉花对棉铃虫的抗虫性及对二种地老虎幼虫生长发育的影响. 应用昆虫学报, 49(4): 882-888.
梁革梅, 谭维嘉, 郭予元. 2000. 棉铃虫对转Bt基因棉的抗性筛选及遗传方式的研究. 昆虫学报, 43(S1): 57-62.
李捷, 张兴华, 马艳, 杨兆光, 乔艳艳. 2013. 转双价抗虫基因(Cry1Ac+Cry2Ab)棉靶标害虫不防治环境下检测技术和生存竞争能力研究. 安徽农业科学, 41(6): 2523-2526.
雒珺瑜, 崔金杰, 辛惠江. 2011a. 转Cry1Ac+Cry2Ab基因棉花对天敌生长发育的影响. 中国棉花, 38(12): 11-12.
雒珺瑜, 崔金杰, 张帅, 陆雪君. 2011b. 转Cry1Ac+Cry2Ab基因棉对棉田节肢动物群落的影响. 植物保护, 37(6): 90-92, 111.
雒珺瑜, 崔金杰, 张帅, 王春义, 辛惠江. 2012. 转Cry1Ac+Cry2Ab基因棉对棉蚜生命表参数及种群动态的影响. 应用昆虫学报, 49(4): 906-910.
雒珺瑜, 崔金杰, 王春义, 辛惠江. 2014a. 转Cry1Ac+Cry2Ab基因棉花对小地老虎生长发育的影响. 棉花学报, 26(5): 179-183.
雒珺瑜, 刘传亮, 张帅, 王春义, 吕丽敏, 李春花, 李付广, 崔金杰. 2014b. 转RRM2基因棉生长势和产量及对棉田节肢动物群落的影响. 植物生态学报, 38(7): 785-794.
马惠,夏晓明,周玉,赵鸣,王红艳. 2009. 转Bt基因抗虫棉对非靶标昆虫的影响. 中国农学通报, 25(7): 214-218.
马小艳, 马艳, 彭军, 姜伟丽, 马亚杰. 2013a. 转基因抗草甘膦抗虫棉的荒地生存竞争能力. 生物安全学报, 22(2): 103-108.
马小艳, 彭军, 姜伟丽, 马亚杰, 马艳. 2013b. 转基因(Bn-csRRM2)高产棉花的荒地生存竞争能力. 生物安全学报, 22(4): 248-252.
乔艳艳, 张兴华, 马艳, 李捷, 杨兆光. 2013. 转Cry1Ac+Cry2Ab基因棉与转Cry1Ac+EPSPS基因棉在黄萎病菌压力环境下检测技术和生存竞争能力研究. 中国农学通报, 29(36): 281-286.
戎俊, 宋志平, 苏军, 夏辉, 王锋, 卢宝荣. 2006.Bt/CpTI转基因稻及其非转基因亲本对照在间隔种植条件下的转基因漂移. 生物多样性, 14(4): 309-314.
苏宏华,范娜,李达,杨益众. 2010. 转Bt基因棉对甜菜夜蛾的影响. 植物保护, 36(5): 1-5.
王香河. 2003. 棉花留叶枝对产量的超补偿效应研究. 北京:中国农业科学院.
徐广, 王桂荣, 吴孔明, 郭予元. 2002. 棉铃虫不同地理种群间基因流动的RFLP分析. 棉花学报, 14(6): 352-355.
张宝红, 郭腾龙. 2000. 转基因棉花基因花粉散布频率及距离的研究. 应用与环境生物学报, 6(1): 39-42.
张兴华, 田绍仁, 张天玉, 李捷, 乔艳艳. 2012a. 转cry1Ac+cry2Ab基因棉与转cry1Ac+EPSPS基因棉荒地的生存竞争能力. 生物安全学报, 21(2): 119-124.
张兴华, 田绍仁, 张天玉, 李捷, 杨兆光, 乔艳艳. 2012b. 转双价双Bt抗虫基因(Cry1Ac+Cry2Ab)棉与非转基因棉生物学特性差异分析. 中国农学通报, 28(24): 35-41.
中华人民共和国农业部. 2007. 农业部 953 号公告-12.2-2007 转基因植物及其产品环境安全检测抗虫棉花第2部分: 生存竞争能力.http:∥www.moa.gov.cn/zwllm/tzgg/gg/200801/t20080110_952066htm.
朱家林, 贺娟, 牛建群, 张青文, 刘小侠. 2013. 风向因素对转基因抗虫棉花基因漂移效率的影响. 生态学报, 33(21): 6803-6812.
左娇, 郭运玲, 孔华, 黄启星, 徐林, 周霞, 郭安平. 2013. 转基因棉花环境安全性研究进展. 热带农业工程, 37(2): 5-10.
Bashir L. 2014. 同类型Bt棉花的抗虫性及棉铃虫对Bt蛋白的抗性动态监测. 北京:中国农业科学院.
McGaughey W H, MeGaughey and Whalon M E. 1992. Managing inscet resistance toBacillusthuringiensistoxins.Science, 258: 1451-1455.
Tabashnik B E, Liu Y B, Finson N, Masson L and Heekel D G. 1997. One gene in diamondback moth confers resistance to fourBacillusthuingiensistoxins.AgriculturalSciences, 94: 1640-1644.
猜你喜欢
学与玩(2022年10期)2022-11-23 08:32:00
今日农业(2022年3期)2022-06-05 07:12:08
新疆农垦科技(2016年2期)2016-08-21 13:50:22
中国农业文摘-农业工程(2016年5期)2016-04-12 05:38:09
现代农业(2016年5期)2016-02-28 18:42:35
现代农业(2016年4期)2016-02-28 18:42:09
创新科技(2015年1期)2015-12-24 06:23:21
现代农业(2015年5期)2015-02-28 18:40:42
新疆农垦科技(2014年12期)2014-02-28 19:21:46
新疆农垦科技(2014年6期)2014-02-28 19:20:16
